三江黄牛Y染色体ZFY基因遗传多样性研究
2017-12-29柴志欣宋娜娜钟金城何世明吴锦波蹇尚林
柴志欣,宋娜娜,钟金城*,何世明,吴锦波,蹇尚林
三江黄牛Y染色体ZFY基因遗传多样性研究
柴志欣1,宋娜娜1,钟金城1*,何世明2,吴锦波2,蹇尚林3
(1.西南民族大学青藏高原研究院,成都 610041;2.阿坝州畜牧科学研究所,四川 汶川 623000;3.阿坝州畜牧工作站,四川 汶川 623000)
三江黄牛原产于四川省阿坝藏族羌族自治州汶川县,具有躯干较长、役用性能良好、适应性强、肉质好等特点,是经长期人工选育的优良地方黄牛品种,为遗传资源宝贵基因库。试验对三江黄牛Y染色体特异ZFY基因作遗传多态性分析,发掘三江黄牛遗传起源。结果表明,ZFY基因第11外显子长度为1 090 bp,T、A、C、G平均含量分别为24.2%、34.2%、20.7%、20.9%,存在碱基偏好性;发现22个突变位点,18个转换,4个颠换,其核苷酸多样性(Pi)为0.007 61,检出16种单倍型,单倍型多样性(Hd)为0.993,说明三江黄牛群体ZFY基因遗传多样性较丰富;系统进化和遗传距离分析表明,三江黄牛优先和大额牛、瘤牛、野牛聚为一类,研究为濒危三江黄牛品种保种、选育及来源提供依据。
三江黄牛;ZFY基因;遗传多样性
三江黄牛产于四川省阿坝藏族羌族自治州汶川县,三江、水磨、白石和映秀等乡镇为主产区,理县、茂汶等地有少量分布,具有躯干较长、役用性能良好、肉质好、抗病力和适应性强等特点,经长期选育形成的优良地方黄牛品种。但随当地经济社会发展和农业生产方式改变,三江黄牛产区功能布局空间受限,三江黄牛养殖规模、种群数量锐减,目前仅存2 000余头,濒临灭绝。因此,加强三江黄牛遗传资源研究和保护尤为重要。
目前,三江黄牛分子遗传学研究较少,主要集中在线粒体基因组等母系遗传基础上,种群内Y染色体重组多样性相对较低,但分化程度较高[1-4]。因此,从Y染色体角度分析物种遗传多样性具有理论意义[5]。 ZFY基因是Y染色体锌指蛋白(Zinc Finger Protein)编码基因,位于 Y染色体短臂,包含 11个外显子和 1个随机重复区域“锌-指”结构,“锌-指”结构由 2个组氨酸和 2个半胱氨酸构成[6-7]。ZFY基因共编码 801个氨基酸,其碱基序列属于 X-退化[8],在未发现SRY基因为性别决定因子前,ZFY基因作为重要候选基因用于物种性别鉴定[9]。ZFX和 ZFY基因具有较高一致性,但研究表明二者进化过程不同,ZFY基因变异率和遗传变异程度更高,进化中更活跃,ZFX基因相对 ZFY基因较保守[10-12]。针对 Y染色体基因系统进化关系研究较多。毛德才等通过扩增牦牛ZFX、ZFY基因第 11外显子部分序列,与其他 9个物种比较分析发现 ZFX和 ZFY序列具有较高一致性[13]。朱睦楠等通过系统进化分析岩羊和矮岩羊 226份粪便cytb基因和 Y染色体ZFY基因发现,喜马拉雅塔尔羊和岩羊、山羊遗传距离最近,绵羊与山羊、塔尔羊、岩羊亲缘关系相对较远[14]。熊勇等通过测序获得藏系绵羊ZFX、ZFY基因序列,探讨藏系绵羊遗传进化情况,发现藏系绵羊与摩弗伦羊亲缘关系最近[15]。金梅等利用ZFX、ZFY基因鉴定绒山羊性别和进化情况[16]。Isaäcj等分析性染色体相关ZFX和 ZFY基因序列特征和多态性差异[17]。Han等探讨马ZFX、ZFY基因片段长度差异[18]。本研究通过测序 Y染色体特异性ZFY基因,结合其他牛亚科该基因序列数据,探讨三江黄牛群体遗传多样性水平,为三江黄牛遗传资源的保护与利用提供理论依据。
1 材料与方法
1.1 材料
样本采集于四川省阿坝藏族羌族自治州汶川县三江乡和水磨镇,选取毛色黄色、体型较大、特征明显三江黄牛个体,采集耳组织样 60份(60头个体),75%乙醇保存,带回实验室,-80℃备用。
1.2 总DNA提取
三江黄牛总 DNA采用动物组织基因组DNA提取试剂盒(购自天根生物技术有限公司),DNA纯度采用2%琼脂糖凝胶电泳和紫外分光光度计(A260/A280比值)检测。
1.3 引物序列及合成
利用 Primer primer5.0软件对三江黄牛 ZFY基因第 11外显子设计两对引物(见表1)。引物由英潍捷基(上海)生物技术有限公司合成。
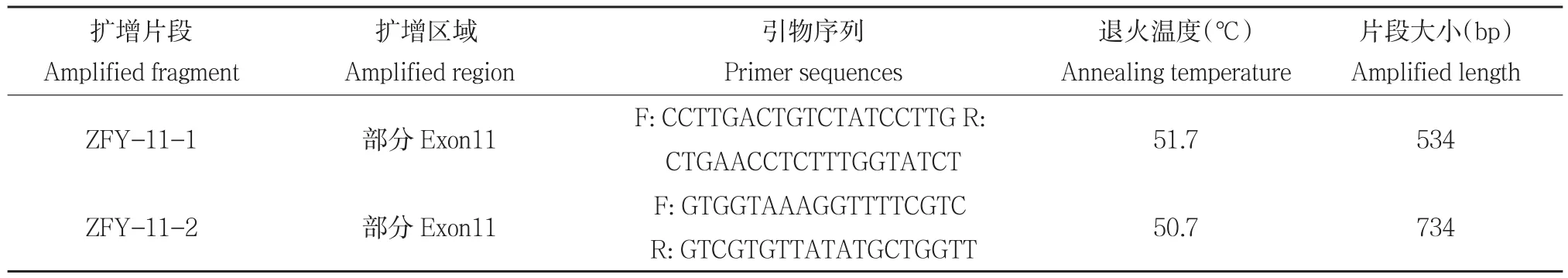
表1 三江黄牛ZFY基因引物序列Table 1 Primer of ZFY gene in Sanjiang cattle
1.4 PCR扩增
PCR 反应体系(25 μL):ddH2O 9.5 μL,上游引物、下游引物、DNA模板各 1μL,2×long Taq DNA 预混酶(Thermo Scientific)12.5 μL。
PCR反应条件:94℃预变性 4 min;94℃变性 1 min;51.7℃复性 1 min;72℃延伸 45 min(引物1)和50 min(引物2),30 个循环;最后72℃延伸 7 min, 4℃保存。PCR 产物用 2%琼脂糖凝胶电泳,95 V恒压电泳 30 min,EB染色,凝胶成像系统拍照检测。
1.5 克隆及测序
将PCR产物胶回收、分离、纯化得目的片段,与 PMD19-T载体连接成重组质粒,转化入感受态细胞(E.coli DH5a)中,涂于含有 IPTG、X-Gal、Ampr抗生素平板上筛选蓝白斑,获白色单一菌落,接种于含 Amp LB液体培养基上获得阳性菌落,送英潍捷基(上海)生物技术有限公司测序。
1.6 牛亚科代表物种ZFY序列
测序得三江黄牛 ZFY基因序列,Genebank获得大额牛(Bos frontalis)(DQ336550)、非洲水牛(Syncerus caffer)(DQ336554)、家牦牛(Bos grunni⁃ens)(DQ 336551)、 美 洲 野 牛(Bison bison)(DQ336552)、普通牛(Bos taurus)(DQ336546)、欧洲野牛(Bison bonasus)(DQ336553)、水牛(Bubalus bubalis)(DQ 336555) 、 野 牛 (Bos gaurus)(DQ336549)、瘤牛(Bosindicus)(DQ336547)等牛亚科代表物种ZFY基因序列。
参考汪琦等测序方法获得三江黄牛cytb序列[19], 从 Genebank 获 得 大 额 牛(Bos frontalis)(EF061237) 、 非 洲 水 牛 (Syncerus caffer)(AF036275) 、 家 牦 牛 (Bos grunniens)(AF091631) 、 美 洲 野 牛 (Bison bison)(AF036273)、普通牛(Bostaurus)(D34635)、欧洲野牛(Bison bonasus)(Y15005)、水牛(Bubalusbubalis)(D88631)、野牛(Bos gaurus(AY079128)、 瘤牛(Bosindicus)(AF419237)等牛亚科代表物种 cytb基因序列。
1.7 数据处理
采用 DNAMAN、Seqman和 DNASTAR等软件校正、比对、拼接所测序列。clustalX1.83比对Genebank下载同源序列[20];选取牛亚科代表物种,NJ(Neighbor Joining Method)法构建分子系统进化树,MEGA5.0计算遗传距离[21];DNAsp4.0软件统计多态位点(Polymorphic Site)、核苷酸多样性(Nucleotide diversity)、核苷酸单倍型数目(Number of Haplotypes)、单倍型多样度(Haplo⁃type diversity)及 Tajima′s D 中性检验值[22]。
2 结果与分析
2.1 三江黄牛ZFY基因PCR扩增
参考普通牛 ZFY基因全序列(Accession No.:NC_016145),利用 ZFY-11-1引物扩增三江黄牛不同个体第 11外显子部分序列,片段长 602 bp(见图1),测序所得序列与参考基因比对完全匹配,说明扩增结果正确。ZFX基因和 ZFY基因一致性较高,属同源基因,主要差异为ZFX基因上存在 AvaⅡ 酶切位点(识别位点),而 ZFY基因不存在[23-24],因此根据酶切位点鉴定雄性个体。将扩增出 60个个体序列利用在线酶切位点分析软 件 (http://watcut.uwaterloo.ca/template.php?act=snp_new)分析发现共有 17个个体序列未被 AvaⅡ酶切,鉴定 17个个体为雄性,43个个体为雌性。利用 ZFY-11-2引物扩增雄性个体,目的片段长度为 734 bp(见图2)。将两对引物扩增序列拼接获得 1 090 bp片段,用于后续分析。
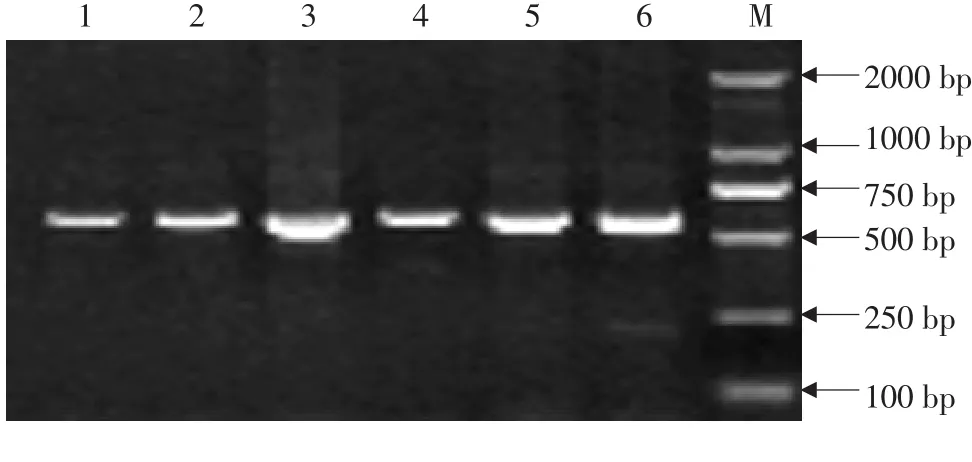
图1 三江黄牛ZFY-11-1引物PCR产物电泳结果Fig.1 Electrophoresisof PCR products of ZFY-11-1 primers of Sanjiang cattle
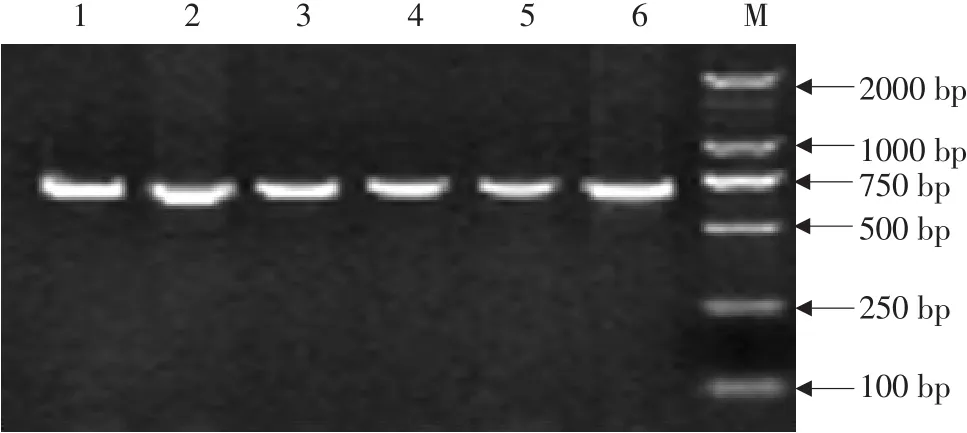
图2 三江黄牛ZFY-11-2引物PCR产物电泳结果Fig.2 Electrophoresisof PCR products of ZFY-11-2 primers of Sanjiang cattle
2.2 三江黄牛ZFX/ZFY基因一致性比对
采用 DNAMAN软件比对ZFY-11-1引物扩增出 43头雌性个体 ZFX基因和 17头雄性个体 ZFY基因序列,一致性为 97.3%。将三江黄牛与 NCBI上获得绵羊、家猫、家犬、人、黑猩猩、猴子、家鼠 ZFY/ZFX基因核苷酸序列比对,三江黄牛 ZFY基因核苷酸序列与绵羊ZFY序列一致性最高为 95.55%,与家猫、家犬、人、黑猩猩、猴子、家鼠一致性为 87.72~92.35%。ZFX基因核苷酸序列与绵羊 ZFX序列一致性为98.72%,和家猫、家犬、人、黑猩猩、猴子、家鼠的一致性为 92.35~95.81%(见表2)。ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/) 在 线 分 析将核苷酸序列翻译为氨基酸序列,结合 DNA⁃MAN软件将三江黄牛候选基因氨基酸序列与其他 7个物种比对,可知三江黄牛与其他物种ZFY基因氨基酸序列一致性为 92.13%~97.19%,ZFX基因氨基酸序列一致性为98.85%~100%(见表3)。

表2 ZFY/ZFX基因核苷酸序列一致性比较Table 2 Consistency comparison of theof ZFY/ZFX gene nucleotide sequences (%)
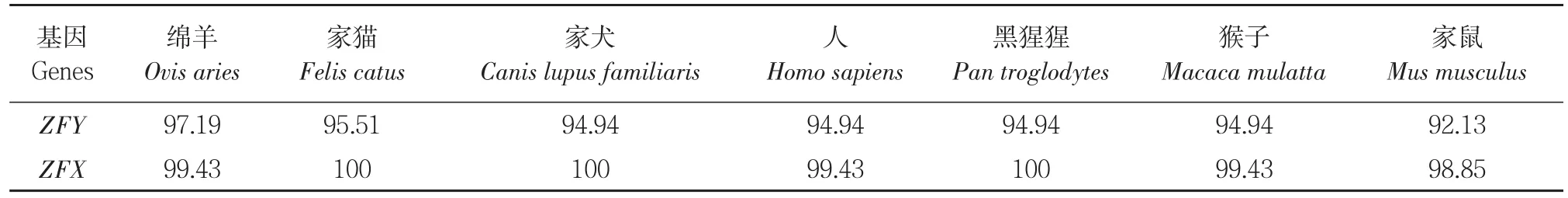
表3 ZFY/ZFX基因编码氨基酸序列一致性比较Table3 Consistency comparison of amino acid sequenceencoded by ZFY/ZFX gene (%)
2.3 三江黄牛ZFY基因遗传多样性
两对引物扩增序列拼接得基因11外显子部分片段长度为 1 090 bp,个体间序列无长度差异,T、A、C、G平均比例分别为 24.2%、34.2%、20.7%、20.9%,A+T含量为 58.3%,G+C含量为 41.7%,说明 ZFY基因 A+T含量相对丰富,存在一定碱基偏倚性。
利用DNAsp软件计算三江黄牛 ZFY基因核苷酸多样性和单倍型多样性,获 16种单倍型(见表4)和 22个同义突变位点,其中 18个转换,4个颠换,单倍型多样性(Hd)为 0.993,核苷酸多样性(Pi)为 0.007 61。单倍型分类中仅第 2种单倍型包含2个个体(18号,55号),其余 15种单倍型仅包含1个个体,但单倍型种类较多,说明三江黄牛群体遗传多样性较丰富。试验获得ZFY基因序列保守区域1个,长度 374~1 076 bp,A、T、C、G含量分别为 34.1%、24.5%、21.1%、20.3%,在该保守区域 A+T含量为 58.6%,与总A+T碱基含量相近。Tajima值为 1.04 088,中性检验结果不显著(P>0.1)。

表4 三江黄牛ZFY基因单倍型分布Table 4 ZFY gene haplotypesof Sanjiang cattle
2.4 三江黄牛与其他牛种的遗传距离及系统进化分析
利用三江黄牛 ZFY基因序列,选取 9个牛亚科代表性物种,NJ法构建系统进化树(见图3),可聚为 2大类,三江黄牛和大额牛、瘤牛、野牛聚为一类;普通牛、欧洲野牛、美洲野牛、家牦牛聚为一类;之后三江黄牛、大额牛、瘤牛、野牛、普通牛、欧洲野牛、美洲野牛、家牦牛聚为一大类;非洲水牛和水牛聚为一类。利用Cytb基因序列聚类结果也为2类,三江黄牛首先与大额牛聚为一类;瘤牛和野牛聚为一类;三江黄牛、大额牛、瘤牛、野牛又与普通牛、欧洲野牛聚为一类;家牦牛与美洲野牛聚为一类;之后三江黄牛、大额牛、瘤牛、野牛、普通牛、欧洲野牛与家牦牛、美洲野牛聚为一大类;非洲水牛和水牛聚为一类。三江黄牛 ZFY基因和Cytb基因聚类结果相同,均优先与大额牛、瘤牛、野牛聚为一类,说明其亲缘关系较接近。MEGA5.0软件分析 10个牛种间的遗传距离,ZFY基因双参数遗传距离(见表5),三江黄牛与大额牛、瘤牛、野牛的遗传距离为 0,说明三者之间遗传距离较近。普通牛、欧洲野牛、美洲野牛、家牦牛为0.004,非洲水牛最远为0.012,水牛为0.010,与地理位置分布一致。cytb基因 Kimura双参数遗传距离(见表6),可知三江黄牛与大额牛遗传距离最近为 0.002,与瘤牛、野牛遗传距离相对较近,分别为 0.016和0.018。10个品种间普通牛和非洲水牛距离最远为0.153。ZFY基因计算遗传距离结果低于cytb,说明线粒体基因的分化程度高于 Y染色体基因,但两种结果均说明三江黄牛与大额牛、瘤牛、野牛的遗传距离更接近,且与聚类分析结果一致。
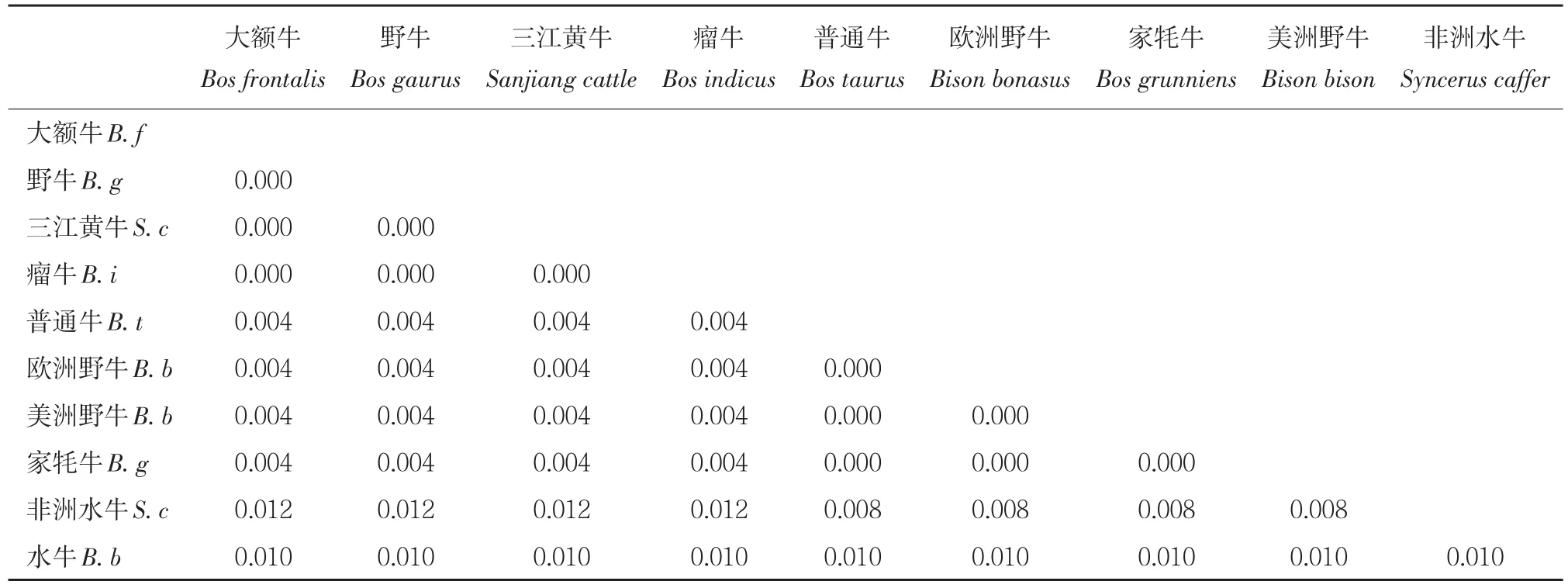
表5 基于ZFY基因牛亚科不同物种间遗传距离Table 5 genetic distance among species of Bovinae ZFY gene
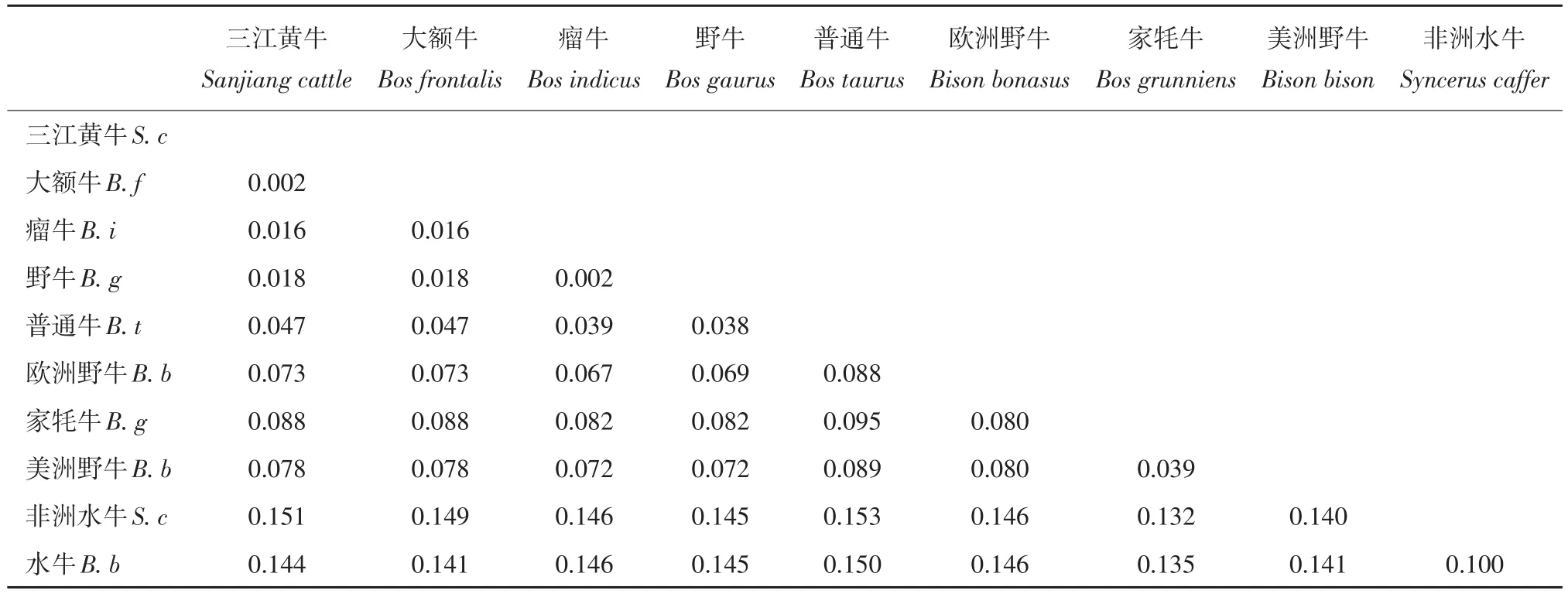
表6 基于cytb基因牛亚科不同物种间遗传距离Table 6 genetic distance among species of Bovinae cytb gene
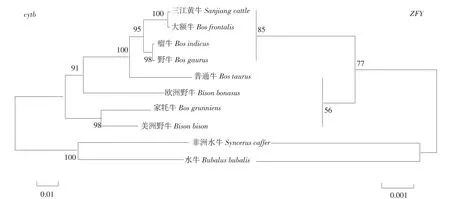
图3 NJ法构建牛亚科系统发育树Fig.3 Neighborjoining treereconstructed in Bovinae
3 讨论与结论
近年研究发现,在生物进化过程中,ZFX较ZFY稳定、保守,ZFY基因变异率大于ZFX。ZFX/ZFY作为性别鉴定主要工具,依据样品类型如粪便、羽毛等对稀有物种作性别鉴定。线粒体基因组作为母系遗传研究主要依据,为物种起源进化及遗传多样性研究提供理论支持;而动物Y染色体遵循父系遗传,突变率低,不易受重组和回复突变等因素影响,是研究父系起源进化及遗传多样性理想工具。近年来,随Y染色体SNPs遗传标记(Y-SNPs)广泛用于家牛起源进化研究[25]。
本研究利用Y染色体ZFY基因,参考已测序三江黄牛cytb序列作联合分析,利用ZFY-11-1引物扩增出ZFY基因第11外显子602 bp序列,通过在线酶切位点软件分析发现无AvaII酶切位点17个个体,鉴定为雄性个体(与采样记录性别结果一致),ZFX/ZFY基因序列一致性比对结果为97.3%,与魏丽敏等研究马鹿ZFX/ZFY基因序列一致性结果相同,说明不同物种ZFX/ZFY基因间存在变异[26]。通过对三江黄牛与另外7个物种ZFX/ZFY基因核苷酸序列比对分析可知,ZFX/ZFY基因核苷酸序列与绵羊序列一致性高,与其他6个物种序列一致性为87.72%~92.35%,氨基酸序列一致性为92.35%~95.81%,同时不同物种ZFX基因比对一致性结果均高于ZFY,说明ZFX基因在进化过程中相对稳定。
ZFY-11-2引物扩增17个雄性个体第11外显子部分序列,长734 bp,两对引物扩增序列拼接获得1 090 bp片段,针对拼接片段分析得T、A、C、G平均比例分别为24.2%、34.2%、20.7%、20.9%,表现一定偏倚性。17个雄性个体中共鉴定出22个突变位点,其中18个转换,4个颠换,转换数量明显多于颠换,说明ZFY基因碱基变异具有同位性、保守性特点,其核苷酸多样性(Pi)为0.007 61,与郝荣超等报道荷斯坦牛核苷酸多样性结果(0.006 25)接近[27]。针对17个个体检出16种单倍型,单倍型多样性(Hd)为0.993,说明三江黄牛群体单倍型多样性丰富,高于刘若余等报道贵州黄牛4个品种单倍型多样性(0.695~0.909)[28],与赖松家等研究四川5个黄牛品种单倍型多样性结果相比,高于巴山黄牛(0.9048)、峨边花牛(0.6000)、四川荷斯坦(0.5000),低于汉源黄牛(1.0000)[29],说明三江黄牛群体遗传多样性较丰富,Tajima值为1.040 88,中性检验结果不显著。
系统进化和遗传距离分析,结果聚为2大类,三江黄牛优先和大额牛、瘤牛、野牛聚为一类,再与其他牛种聚为一类,与汪琦等对三江黄牛D-loop序列聚类结果一致[30]。cytb序列系统进化分析得到2大类,三江黄牛与大额牛、瘤牛、野牛先聚为一类,再与其他牛种聚为一类。分析结果均说明三江黄牛与大额牛、瘤牛、野牛亲缘关系较近、遗传相似性高,而与其他牛属遗传相似性相对较低。优先与瘤牛聚为一类说明三江黄牛有瘤牛血统,与张成忠通过Y染色体G带带型、Ag-NORs、血液蛋白多态性等分析的结果一致,认为四川黄牛属瘤牛种[31]。张桂香等分析推测云南和四川等地区可能是中国瘤牛发源地之一[32],说明三江黄牛可能为瘤牛后代。大额牛主产于云南贡山县独龙江一带,从地理位置上和四川接近,推测为地缘或贸易等因素促进两种牛类基因交流,使之聚为一类。
[1] Hellborg L,Ellegren H.Low levels of nucleotide diversity in mammalian Y chromosomes[J].Molecular Biology and Evo⁃lution,2004,21(1):158-163.
[2] Lawson L J,Hewitt G M.Comparison of substitution rates in ZFX and ZFY introns of sheep and goat related species supports the hypothesis of male-biased mutation rates[J].Journal of Molecular Evolution,2002,54(1):54-61.
[3] Hughes J F,Skaletsky H,Pyntikova T,et al.Conservation of Y-linked genes during human evolution revealed by compar⁃ative sequencing in chimpanzee[J].Nature,2005,437(7055):100-103.
[4] Brändli L,Handley L J,Vogel P,et al.Evolutionary histo⁃ry of the greater white-toothed shrew(Crocidura russula)in⁃ferred from analysis of mtDNA,Y,and X chromosome mark⁃ers[J].Molecular Phylogenetics and Evolution,2005,37(3):832-844.
[5] Zluvova J,Janousek B,Negrutiu I,et al.Comparison of the X and Y chromosome organization in Silene latifolia[J].Genetics,2005,170(3):1431-1434
[6] Palmer M S,Berta P,Sinclair A H,et al.Comparison of human ZFY and ZFX transcripts[J].Proceedings of the National Academy of Sciences,1990,87(5):1681-1685.
[7] Brown R S,Sander C,Argos P.The primary structure of transcription factor TFIIIA has 12 consecutive repeats[J].FEBS Letters,1985,186(2):271-274.
[8] Skaletsky H,Kuroda-Kawaguchi T,Minx P J,et al.The male-specific region of the human Y chromosome is a mosa⁃ic of discrete sequence classes[J].Nature,2003,423(6942):825-837.
[9] Mardon G,Page D C.The sex-determining region of the mouse Y chromosome encodes a protein with a highly acid⁃ic domain and 13 zinc fingers[J].Cell,1989,56(5):765-770.
[10] Iwase M,Satta Y,Hirai Y,et al.The amelogenin loci span an ancient pseudoautosomal boundary in diverse mammalian spe⁃cies[J].Proceedings of the National Academy of Sciences,2003,100(9):5258-5263.
[11] Gerrard D T,Filatov D A.Positive and negative selection on mammalian Y chromosomes[J].Molecular Biology and Evolution,2005,22(6):1423-1432.
[12] Jegalian K,Page D C.A proposed path by which genes common to mammalian X and Y chromosomes evolve to be⁃come X inactivated[J].Nature,1998,394(6695):776-780.
[13] 毛德才,徐亚欧,熊勇,等.牦牛与其他物种 ZFX/ZFY基因片段间的进化关系[J].中国生物化学与分子生物学报,2007,23(6):450-456.
[14] 朱睦楠,周材权,何娅,等.基于线粒体Cytb和核基因ZFY探讨羊族物种之间的系统发生关系[J].兽类学报,2014,34(4):366.
[15] 熊勇,徐亚欧.藏系绵羊ZFX/ZFY基因片段进化分析[J].生物技术,2010,20(6):1-5
[16] 金梅,傅忠扬,栾媛媛,等.辽宁新品系绒山羊 ZFX,ZFY基因片段的进化研究与性别鉴定[J].畜牧兽医学报,2006,37(6):530-536.
[17] Isaäcj N,Dickcj V B,Lisettem V C,et al.Phylogeny of Y chromosomes from bovine species[J].Cladistics-the Interna⁃tional Journal of the Willi Hennig Society,2010,24(5):723-726.
[18] Han S H,Yang B C,Ko M S,et al.Length difference be⁃tween equine ZFX and ZFY genes and its application for molecular sex determination[J].Journal of assisted reproduc⁃tion and genetics,2010,27(12):725-728.
[19] 汪琦,钟金城,柴志欣,等.三江黄牛mtDNA Cytb基因序列多态性及其系统进化分析[J].中国畜牧杂志,2016,52(15):20-27.
[20] Thompson J D,Gibson T J,Plewniak F,et al.The CLUST⁃AL_X windows interface:Flexible strategies for multiple se⁃quence alignment aided by quality analysis tools[J].Nucleic acids research,1997,25(24):4876-4882.
[21] Tamura K,Dudley J,Nei M,Kumar S.MEGA4:Molecular evolutionary genetics analysis(MEGA)software version 4.0[J].Molecular biology and evolution,2007,24(8):1596-1599.
[22] Rozas J,Sánchez-DelBarrio J C,Messeguer X,et al.DnaSP,DNA polymorphism analyses by the coalescent and other methods[J].Bioinformatics,2003,19(18):2496-2497.
[23] Aasen E,Medrano J F.Amplification of the ZFY and ZFX genes for sex identification in humans,cattle,sheep and goats[J].Nature Biotechnology,1990,8(12):1279-1281.
[24] Reynolds R,Varlaro J.Gender determination of forensic sam⁃ples using PCR amplification of ZFX/ZFY gene sequences[J].Journal of Forensic Science,1996,41(2):279-286.
[25] 马志杰,黄永震,党瑞华,等.日本和牛Y染色体USP9Y基因多态性与父系起源研究[J].中国牛业科学,2017,43(1):1-3,7.
[26] 魏丽敏,孙丽荣,张永生,等.马鹿 Zfx/Zfy基因的克隆及序列分析[J].中国畜牧兽医,2016,43(9):2272-2278.
[27] 郝荣超,王国华,常玉霞,等.中国荷斯坦牛和鲁西黄牛 mtD⁃
Genetic diversity of ZFY gene on Y chromosome in Sanjiang cattle/
CHAIZhixin,SONG Nana,ZHONG Jincheng,HE Shiming,WU Jinbo,JIAN Shanglin
(1.Institute of Qinghai-Tibetan Plateau,Southwest Minzu University,Chengdu 610041,China;2.Animal Husbandry Science Institute of Aba Autonomous Prefecture,Sichuan Wenchuan 623000,China;3 Animal Husbandry and Veterinary Station of Aba Autonomous Prefecture,Sichuan Wenchuan 623000,China)
Native to Wenchuan County,Aba Tibetan and Qiang Autonomous Prefecture,Sichuan Province,Sanjiang Cattle,featured by long body,good working capacity and adaptability as well as highquality meat,was a local improved cattle breed artificially selected and bred in long time,and it was also an extremely precious gene pool in genetic resources.This test made an analysis on its genetic polymorphism of Sanjiang Cattle in term of Y-chromosome specific ZFY gene to explore its genetic ancestry.According to the results,the exon 11 of ZFY gene was 1 090 bp long,and the contents of T,A,C and G were 24.2%,34.2%,20.7%and 20.9%in average respectively,showing a base bias;there were 22 mutation sites,including 18 in conversion and four in transversion,with the nucleotide diversity(Pi)of 0.007 61,and 16 haplotypes detected with the diversity(Hd)of 0.993,all of which meant that the Sanjiang Cattle group had a richer genetic diversity in ZFY gene.From the view of phyletic evolution and genetic distance analysis,Sanjiang Cattle firstly should be included into the same classification with gayal,zebu and buffalo.Such result could provide the theoretical basis for breed conservation and further breeding of the endangered Sanjiang Cattle breed and might be referenced as a basis for the possible source of Sanjiang Cattle.
Sanjiang cattle;ZFY gene;genetic diversity
Q93-332;S852.62
A
1005-9369(2017)12-0028-08
时间2017-12-18 13:44:45 [URL]http://kns.cnki.net/kcms/detail/23.1391.S.20171218.1344.014.html
柴志欣,宋娜娜,钟金城,等.三江黄牛Y染色体ZFY基因遗传多样性研究[J].东北农业大学学报,2017,48(12):28-35.
Chai Zhixin,Song Nana,Zhong Jincheng,et al.Genetic diversity of ZFY gene on Y chromosome in Sanjiang cattle[J].Journal of Northeast Agricultural University,2017,48(12):28-35.(in Chinese with English abstract)
2017-09-19
中央高校基本科研项目(2016NZYQN30);四川省科技计划项目(2015JY0248);国家肉牛牦牛产业技术体系(CARS-37)
柴志欣(1984-),女,硕士,讲师,研究方向为牦牛遗传育种。E-mail:chaizhixin2525@163.com
*通讯作者:钟金城(1963-),男,博士,教授,研究方向为动物遗传育种学。E-mail:zhongjincheng518@126.com
