鱼山渔场披山海域虾类群落结构及其与环境因子的关系
2017-12-27谢旭俞存根郑基夏陆军毕耜瑶张平邓小艳苗露
谢旭,俞存根,郑基,夏陆军,毕耜瑶,张平,邓小艳,苗露
鱼山渔场披山海域虾类群落结构及其与环境因子的关系
谢旭,俞存根,郑基,夏陆军,毕耜瑶,张平,邓小艳,苗露
(浙江海洋大学水产学院,浙江舟山316022)
为分析鱼山渔场披山海域虾类种类组成、优势种、生物多样性等群落结构特征,根据2015年11月、2016年2月、5月、8月在鱼山渔场披山海域进行底拖网生物资源调查所获得的15个站位资料,定量分析了其群落结构的相似性,以及群落结构与水文环境因子间的关系。结果表明:本次调查共鉴定出虾类15种,隶属于8科11属,其中优势种为脊尾白虾Exopalaemon carinicauda、哈氏仿对虾Parapenaeopsis hardwickii、中华管鞭虾Solenocera crassicornis、细巧仿对虾Parapenaeopsis tenella、中国毛虾Acetes chinensis、葛氏长臂虾Palaemon gravieri和日本鼓虾Alpheus juponicus共7种;不同季节的虾类种类组成差异较大,优势种类组成变化较大,其中以秋季的虾类种类数最多,冬季最少;优势种种类数春季最多,冬季最少;不同季节渔获种类的水深分布有较大差异,虾类种类以5~10 m水深带分布较多,小于5 m水深带较少;多样性指数冬季低于其他季节,其中小于5 m水深带较高,10~15 m水深带较低,虾类多样性指数随水深的增加呈递减趋势。研究表明,水深、底层水温、表层盐度、底层盐度和化学需氧量是影响调查海域虾类种类组成和群落结构特征的主要环境因子。
鱼山渔场披山海域;虾类;多样性;群落结构;环境因子
虾类味道鲜美,经济价值高,属优质海产品。近年来,在中国近海渔业资源不断衰退的情况下,虾类的捕捞已成为海洋捕捞的重要组成部分。同时,虾类作为许多鱼类的饵料,是组成海洋生态系统的重要环节,虾类资源量变动会对整个海洋生态系统的平衡造成明显影响。因此,研究虾类群落变动规律及其与环境因子间的关系,对维护海洋生态系统稳定性及海洋渔业可持续发展具有十分重要的意义。
目前,对东海虾类的研究,主要集中在区系[1-4]、 虾类数量分布[5]和虾类资源量[6-8]方面,而对东海虾类群落特征的研究方面,仅见俞存根等[9]、陈小庆等[10]报道了东海虾类群落结构特征;何贤保等[11]报道了舟山渔场及附近海域虾类群落结构特征;夏陆军等[12-13]、晁文春等[14]报道了南麂列岛海域虾类、甲壳类种类组成和群落结构特征;徐捷等[15]报道了吕泗渔场虾类群落结构特征;齐海明等[16-17]报道了浙江椒江口海域虾类、甲壳类群落结构特征及其与环境因子的关系;张洪亮等[18]报道了浙江南部沿岸产卵场虾类群落结构特征。国外学者对虾类群落结构的研究也较多,Lobry等[19]和Munga等[20]报道了Gironde河口和Malindi-Ungwana湾虾类群落的结构特征。
鱼山渔场披山海域位于瓯江入海口,为台湾暖流、浙江沿岸流和大陆径流交汇区。该海域水质肥沃,营养物质丰富、饵料生物充足、水文环境适宜,是多种海洋生物繁殖、索饵、生长的优良栖息地,有鱼、虾、蟹、贝等400余种。为了保护鱼山渔场披山海域海洋生物多样性及其生境,促进鱼类、贝类资源的恢复,当地政府于2011年成立了玉环披山省级海洋特别保护区[21]。为了解保护区内虾类资源开发及利用现状,研究人员开展了本次虾类的群落结构特征与环境因子关系的调查研究。本研究中,主要根据2015—2016年在鱼山渔场披山海域开展渔业资源拖网调查所获得的虾类数据,分析研究了鱼山渔场披山海域虾类的群落结构特征与水文环境间的关系,旨在掌握鱼山渔场披山海域虾类资源现状,以期为玉环披山省级海洋特别保护区虾类资源的科学保护、管理及其可持续利用提供参考依据。
1 材料与方法
1.1 调查船及调查站位
本研究中数据采自 2015年 11月 (秋季)、2016年2月 (冬季)、2016年5月 (春季)、2016年8月 (夏季)在鱼山渔场披山海域20 m水深以内浅海区开展渔业资源定点拖网调查结果。调查区域为 28°01′41″~ 28°08′11″N, 121°21′18″~121°32′38″E, 共设置15 个调查站位 (图1)。 调查船租用 “浙乐渔休00122号”底拖网渔船,主机功率为220 kW,网具为底拖网,规格为530目×60 mm。船上配备有卫导、探鱼仪,以及导航、定位等助渔助航设备。

图1 调查站位图Fig.1 Survey stations of fishery resources
1.2 方法
1.2.1 样品的采集 调查时每站拖曳约1 h,拖速约3.5 kn。渔业资源调查方法按照 《海洋渔业资源调查规范》 (SC/9403—2012)[22]进行。同时,利用温盐深测定仪CTD同步测定各调查站位的水温、盐度、叶绿素数值。每个调查站位拖网所获的渔获物全部取样装入样品袋中,并进行编号、记录后冰鲜保存,带回实验室分析、鉴定,使用电子天平称重,精确度为0.1 g。
1.2.2 优势种及物种多样性的计算 优势种的判断依据为相对重要性指数 (IRI),其计算公式为

其中:ni、wi分别为第i种虾类的个体数和生物量(kg/m2);N、W分别为虾类总个体数和总生物量(kg/m2);fi为第i种虾在m次取样中出现的次数;m为总取样次数。
优势种的季节更替变化运用种类更替率A分析[23],即与前一季节比较的更替情况,计算公式为

其中:C为相临两个季节间减少或增加的物种数;S为相临两个季节间相同的物种数。
物种多样性主要采用Shannon-Wiener多样性指数 (H′)、均匀度指数 (J)、丰富度指数 (d)3个指标表达,计算公式分别为

其中:ni为第i种虾的个体数;N为虾类总尾数;S为虾类总种数。
1.2.3 调查海域虾类群落结构分析 本研究中采用等级聚类 (非加权组的平均,即UPGAMA)的聚类分析方法 (CLUSTER)和非度量多维标度排序方 法 (Non-metric multidimensional scaling,MDS)分析鱼山渔场披山海域虾类群落结构特征。在Bray-Curtis相似性矩阵基础上作聚类分析图和MDS二维图[24]。聚类 (CLUSTER)和非度量多维标度排序分析均在PRIMER 5.0软件包 (Plymouth Marine Laboratory,UK)[25]上完成。 用 Bray-Curtis相似性指数 (B)计算各季节虾类群落结构组成的差异[26]:

其中:xij、xim分别为第i个种类在j站位和m站位单位时间的渔获量 (经4次方根转换)。
群落季节更替指数(AI)和迁移指数(MI)[27]计算公式分别为

其中:A为各季节实际物种数;C为本季节新迁入物种数;D为本季节将要迁出物种数;R为全年各季节均出现的物种数;AI反映物种更替导致群落稳定性降低的节律,其值越大,表示群落稳定性越差;MI表示研究系统外迁入与迁出种的两种成分在群落中的相对比例,当C>D时,MI为正值,表示迁入种数多于迁出种数,当C=D时,MI则为零,表示群落动态平衡,当C<D时,MI为负值,表示迁出种数多于迁入种数。
1.3 数据处理
虾类数量分布图采用Surfer 8软件绘制。生物量和水文环境之间的关系采用冗余分析 (Redundancy analysis, RDA) 方法分析[28-29], 用 CANOCO 3.0软件进行运算,排序结果用物种-环境因子关系的双序图表示。
DCA排序前4个轴中,最大值大于4时,选用单峰模型CCA,最大值介于3和4之间时,选用单峰模型CCA或者选用线性模型RDA均可,最大值小于3时,选用线性模型RDA;DCA排序前4个轴中,最大值小于3时,选用线性模型RDA。RDA是一种线性多元直接梯度分析方法[30],它把对应分析与多元回归结合起来,每一步计算结果均与环境因子进行回归,以确定虾类群落与环境的对应关系。在RDA排序图中,环境因子用带有箭头的线段表示,连线的长短表示虾类群落和种类分布与该环境因子关系的相关性大小,连线越长,说明相关性越大,反之越小。箭头连线与排序轴的夹角表示该环境因子与排序轴相关性大小,夹角越小,相关性越高,反之越低。箭头连线与某一站位(或种类)的垂直线距离表示其相关性大小,垂线距离越小,正相关性越高,反之越低,处于另一端的则表示与该类环境因子具有负相关性。
2 结果与分析
2.1 种类组成
由表1可见:根据本次调查采集样品共鉴定出虾类15种,隶属于8科11属;秋季出现种类最多,为12种;春、夏季次之,均为11种;冬季较少,为10种。各季节均以对虾科出现种类数最多,周年4个季节均出现的共有种为细巧仿对虾Parapenaeopsis tenella、哈氏仿对虾Parapenaeopsis hardwickii、周氏新对虾Metapenaeus joyneri、脊尾白虾Exopalaemon carinicauda、葛氏长臂虾Palaemon gravieri、鲜明鼓虾Alpheus distinguendus和日本鼓虾Alpheus juponicus7种,占总种类数的46.67%;仅在一个季节出现的虾类有2种,占总种类数的13.33%。本研究中的生物种类名录主要参照 《中国海洋生物名录》[31]。

表1 鱼山渔场披山海域虾类科、属、种的组成Tab.1 Compositions of shrimp at species, genus and family levels in Pishan waters of the Yushan fishing ground
从表2可见:将调查海域按水深分为小于5、5~10、 10~15、 15~20 m 4个水深带, 分析鱼山渔场披山海域的虾类种类组成。从表3可见:春、夏、秋3个季节虾类种类随水深增加呈先递增后递减的趋势;春季,虾类种类数在水深10~15 m区域最多,小于10 m区域次之,15~20 m区域最少;夏、秋季,虾类种类数在水深5~10 m区域最多,10~20 m区域次之,小于5 m区域最少;冬季,虾类种类随水深增加呈递增趋势,在水深10~20 m区域的虾类最多,5~15 m区域次之,小于5 m区域最少。水深小于5 m的区域,虾类春季最多 (9种),冬季节最少 (4种);水深5~10 m的区域,虾类夏、秋季较多 (10种),冬季最少 (8种);水深10~15 m的区域,虾类春季最多 (10种),夏、秋、冬3季较少 (8种);水深15~20 m的区域,虾类冬季最多 (10种),春、夏季较少 (7种)。3个水深带均有分布的共有种较少,春季共有种最多 (7种),夏、冬季次之 (4种),秋季共有种最少 (3种)。这表明,栖息在水深5~10 m区域的虾类种类最多,栖息水深小于5 m区域的虾类种类最少。

表2 鱼山渔场披山海域不同水深带站位点分布Tab.2 The survey stations at different water depths in Pishan waters of the Yushan fishing ground

表3 鱼山渔场披山海域不同水深带虾类种类数组成Tab.3 The shrimp species composition at different water depths in Pishan waters of the Yushan fishing ground
2.2 优势种
根据相对重要性指数 (IRI)分析虾类群落中的优势种成分,结果如表4所示。将IRI>1000的定为优势种,将IRI=100~1000的定为常见种[32]。结果表明,本次调查优势种为脊尾白虾、哈氏仿对虾、中华管鞭虾Solenocera crassicornis、细巧仿对虾、中国毛虾Acetes chinensis、葛氏长臂虾和日本鼓虾等7种,不同季节的优势种存在明显的季节更替,春-夏季、夏-秋季、秋-冬季和冬-春季间优势种更替率分别为14.29%、80.00%、66.67%和80.00%,春季优势种有脊尾白虾、细巧仿对虾、中国毛虾、葛氏长臂虾和日本鼓虾等5种,夏季优势种有哈氏仿对虾、中华管鞭虾和葛氏长臂虾等3种,秋季优势种有脊尾白虾、中华管鞭虾和细巧仿对虾等3种,冬季优势种仅有脊尾白虾1种。

表4 鱼山渔场披山海域主要虾类相对重要性指数 (IRI)的季节变化Tab.4 Seasonal changes in the IRI of shrimp dominant species in Pishan waters of the Yushan fishing ground
2.3 物种多样性指数
比较不同季节虾类的多样性指数值,由表5可见:虾类各季节的丰富度指数以春季最高,秋、冬季次之,夏季最低;均匀度指数以秋季最高,春、夏季次之,冬季最低;多样性指数以秋季最高,春、夏季次之,冬季最低。

表5 鱼山渔场披山海域虾类群落多样性指数的季节变化Tab.5 Seasonal change in the diversity index of shrimp communities in Pishan waters of the Yushan fishing ground
比较不同水深虾类的多样性指数值,由表6可见:虾类的丰富度指数,随水深增加,四季平均值呈先减后增的趋势,春、夏季呈递减,秋季呈先减后增的趋势,冬季呈递增;虾类的均匀度指数,随水深的增加,四季平均值呈先减后增的趋势,春、冬季呈先减后增的趋势,夏季呈递减,秋季呈先增后减的趋势;虾类的多样性指数,随水深的增加,四季平均值呈先减后增的趋势,春、冬季呈先减后增的趋势,夏季呈递减,秋季呈先增后减的趋势。
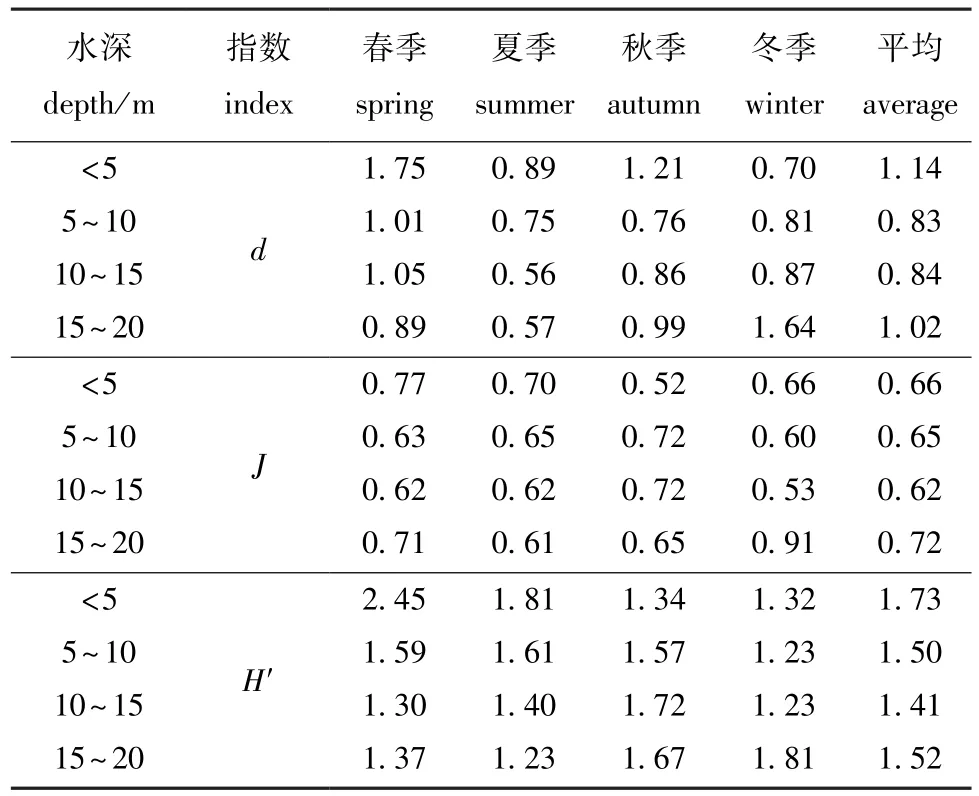
表6 鱼山渔场披山海域不同水深虾类群落多样性指数的季节变化Tab.6 Seasonal changes in diversity indices of shrimp communities in different water depths in Pishan waters of the Yushan fishing ground
2.4 聚类、排序结果分析
对鱼山渔场披山海域虾类生物量进行聚类分析和NMDS排序,结果如图2所示。从图2可知,四季NMDS分析结果的胁迫系数为0.09~0.16,表明该图形较好地反映了群落间的相似性程度[33]。各季节站位的聚类分析和NMDS分析结果基本一致,春、夏季该海域的虾类均可划分为3个群落,分别为群落Ⅰ、群落Ⅱ和群落Ⅲ;秋、冬季均可划分为2个群落,分别为群落Ⅰ和群落Ⅱ。

图2 鱼山渔场披山海域虾类采样站点的聚类分析和NMDS排序图Fig.2 Group average clustering and NMDS ordination of shrimp sampling stations in Pishan waters of the Yushan fishing ground
2.5 群落相似性分析
从表7可见:春季,群落Ⅰ的组内相似性为15.29%,主要特征种为细巧仿对虾、中国毛虾和周氏新对虾等;群落Ⅱ的组内相似性为34.88%,主要特征种为葛氏长臂虾、脊尾白虾、日本鼓虾、细巧仿对虾和周氏新对虾等;群落Ⅲ的组内相似性为28.33%,主要特征种为脊尾白虾和细巧仿对虾等。夏季,群落Ⅰ的组内相似性为58.36%,主要特征种为哈氏仿对虾、脊尾白虾和中华管鞭虾等;群落Ⅱ的组内相似性为58.23%,主要特征种为葛氏长臂虾、哈氏仿对虾、脊尾白虾和中华管鞭虾等;群落Ⅲ的组内相似性为74.68%,主要特征种为哈氏仿对虾和中华管鞭虾等。秋季,群落Ⅰ的组内相似性为57.18%,主要特征种为脊尾白虾、细巧仿对虾和中华管鞭虾等;群落Ⅱ的组内相似性为27.39%,主要特征种为葛氏长臂虾和脊尾白虾等。冬季,群落Ⅰ的组内相似性为57.60%,主要特征种为脊尾白虾等;群落Ⅱ的组内相似性为42.95%,主要特征种为鲜明鼓虾等。

表7 鱼山渔场披山海域虾类各群落内平均相似性贡献率之和占90%以上的特征种Tab.7 Characteristic species for different community groups with average similarity contribution rates accounting for over 90%in Pishan waters of the Yushan fishing ground %
由表8可见:春季,群落Ⅰ和群落Ⅱ的相异性为81.74%,主要分歧种为葛氏长臂虾、脊尾白虾、日本鼓虾、细巧仿对虾、中国毛虾和周氏新对虾等;群落Ⅰ和群落Ⅲ的相异性为90.98%,主要分歧种为脊尾白虾、细巧仿对虾和中国毛虾等;群落Ⅱ和群落Ⅲ的相异性为69.01%,主要分歧种为葛氏长臂虾、脊尾白虾、日本鼓虾、细巧仿对虾和周氏新对虾等。夏季,群落Ⅰ和群落Ⅱ的相异性为41.72%,主要分歧种为葛氏长臂虾、哈氏仿对虾、脊尾白虾和中华管鞭虾等;群落Ⅰ和群落Ⅲ的相异性为38.66%,主要分歧种为哈氏仿对虾、脊尾白虾和中华管鞭虾等;群落Ⅱ和群落Ⅲ的相异性为44.77%,主要分歧种为葛氏长臂虾、哈氏仿对虾、脊尾白虾和中华管鞭虾等。秋季,群落Ⅰ和群落Ⅱ的相异性为68.48%,主要分歧种为脊尾白虾、细巧仿对虾和中华管鞭虾等。冬季,群落Ⅰ和群落Ⅱ的相异性为94.45%,主要分歧种为葛氏长臂虾、脊尾白虾和鲜明鼓虾等。
2.6 群落结构的稳定性
各季节的渔获物种类数、迁入物种数、迁出物种数和各季节均出现的物种数由表9所示,根据这些参数可以算出各季节的更替指数和迁移指数,从而可描绘出季节的变化趋势。由图3可见:虾类群落稳定性最好的为秋季,其次是夏季,春、冬季虾类群落稳定性较差,群落更替指数>120。由图3还可见:春、秋季虾类物种迁入量超过迁出量,冬季虾类物种迁出量超过迁入量。

表8 鱼山渔场披山海域虾类各群落间平均相异性贡献率之和占90%以上的分歧种Tab.8 Discriminating species for different community groups with average dissimilarity contribution rates accounting for over 90%in Pishan waters of the Yushan fishing ground %
注:Av.Diss表示平均差异性,Contrib表示贡献率
Note:Av.Diss=average dissimilarity, Contrib=contribution

表9 鱼山渔场披山海域虾类群落季节更替指数和迁移指数的参数Tab.9 Seasonal AI and MI of shrimp community in Pishan waters of the Yushan fishing ground
2.7 虾类群落与水文环境因子的关系
调查海域虾类群落与表层温度 (ST)、表层盐度 (SS)、底层温度 (BT)、底层盐度 (BS)、水深(depth)、化学需氧量 (COD)和叶绿素(Chl-a)等7个环境因子的RDA排序结果如图4和表10所示。

图3 鱼山渔场披山海域虾类群落更替指数和迁移指数Fig.3 AI and MI of shrimp community in Pishan waters of the Yushan fishing ground

图4 鱼山渔场披山海域虾类与环境因子的RDA排序Fig.4 Seasonal distribution of RDA sort of shrimp and environmental factors in Pishan waters of the Yushan fishing ground

表10 鱼山渔场披山海域虾类群落结构和水文环境因子之间的相关性分析Tab.10 Correlationship between shrimp community structure and environmental factors in Pishan waters of the Yushan fishing ground
从表10和图4可见:春季,调查海域虾类群落受表层盐度和底层盐度极显著影响 (p<0.01),表层盐度与第一、第二排序轴的相关性分别为0.200和-0.980,底层盐度与第一、第二排序轴的相关性分别为0.129和-0.992,PS12号站位受其影响显著。夏季,调查海域虾类群落受底层温度和水深影响显著 (p<0.05),底层温度与第一、第二排序轴的相关性分别为-0.934和-0.356,葛氏长臂虾、细巧仿对虾和刀额新对虾Metapenaeus ensis受其影响较大,PS12和PS14号站位受其影响显著;水深与第一、第二排序轴的相关性分别为0.998和0.063,中国毛虾受其影响较大,PS1和PS9号站位受其影响显著。秋季,调查海域虾类群落受化学需氧量影响较大 (p<0.05),化学需氧量与第一、第二排序轴的相关性分别为0.227和0.974,PS10号站位受其影响显著。冬季,调查海域虾类群落受底层温度影响较大,底层温度与第一、第二排序轴的相关性分别为0.078和-0.997,安氏白虾Exopalaemon annandalei受其影响较大,PS4和PS9号站位受其影响显著。
3 讨论
3.1 虾类群落种类组成
本次调查中,共鉴定出虾类15种,隶属于8科11属,秋季 (12种)>春季 (11种)=夏季(11种)>冬季 (10种)。4个季节均有分布的种类有细巧仿对虾、哈氏仿对虾、周氏新对虾、脊尾白虾、葛氏长臂虾、鲜明鼓虾和日本鼓虾7种,约占虾类总种数的46.67%,而大部分虾类仅在某些季节出现,如东海红虾Plesionika izumiae仅在春季出现,戴氏赤虾Metapenaeopsis dalei仅在秋季出现,这表明鱼山渔场披山海域虾类种类以季节性种类为主。与夏陆军等[12-13]于2013—2014年对相邻海域南麂列岛海洋自然保护区调查结果相比,种类数减少了40%,4个季节均出现的种类增加了40%。从生态类型看,15种虾类大多为广温、广盐性种类,这可能与台湾暖流和浙江沿岸流交互影响有关[13]。从不同水深区域分析,春、夏、秋3个季节虾类种类数随水深的增加呈先增后减的趋势,以水深在5~10 m区域虾类种数最多,在水深小于5 m区域最少,冬季虾类种类数随水深增加呈递增趋势。说明春、夏、秋3个季节许多虾类洄游到此索饵、生殖,并且主要栖息于水深在5~10 m区域,冬季随温度降低,虾类向深水区迁移。
3.2 优势种
与夏陆军等[12-13]于2013—2014年对相临海域南麂列岛海洋自然保护区的调查结果相比,本次调查优势种中减少了鲜明鼓虾,增加了中国毛虾和葛氏长臂虾2种。不同季节的优势种存在明显的季节更替,夏-秋季间优势种更替率为80.00%,这可能是优势种脊尾白虾在 4—6月处于产卵高峰期[34],且夏季饵料生物丰富,水温环境适宜,个体迅速成长,到秋季成为较大个体,使其从夏季的常见种演变为秋季的第一优势种;秋-冬季间优势种更替率为66.67%,冬季随着水温降低,中华管鞭虾和细巧仿对虾等多数虾类向深水区洄游[35],优势种由秋季的脊尾白虾、中华管鞭虾和细巧仿对虾演变为冬季的脊尾白虾;冬-春季间优势种更替率为80.00%,春季随着水温的升高,大量虾类洄游到调查海域进行索饵和生殖,优势种由冬季的脊尾白虾演变为春季的脊尾白虾、细巧仿对虾、中国毛虾、葛氏长臂虾等。
3.3 物种生物多样性
从不同季节虾类的多样性指数来看,冬季调查海域虾类均匀度指数和多样性指数均低于其他季节。这可能是由于调查海域受台湾暖流和浙江沿岸流交互影响导致的,冬季调查海域主要受浙江沿岸流影响[36],导致冬季广温广盐性种类减少 (表4),冬季优势种仅有脊尾白虾1种,而其他季节优势种均有3种以上。另外,冬季鱼类个体较大,许多鱼类捕食虾类[37],也可能是冬季虾类生物多样性最低的原因之一。从不同水深区域分析调查海域的虾类多样性指数可见,春、夏季虾类丰富度指数、均匀度指数和多样性指数均随水深的增加呈递减趋势 (表6)。
3.4 群落结构特征
调查海域虾类群落结构变化明显 (图2)。根据CLUSTER分析,春季和夏季可划分为3个不同的群落,秋季和冬季可划分为2个不同的群落,NMDS二维标序图较好地反映了4个季节群落间的相似性程度。夏季和冬季群落相似性较高,春季群落相似性较低,对相似性影响较大的特征种主要有中国毛虾、葛氏长臂虾、脊尾白虾、日本鼓虾、细巧仿对虾和周氏新对虾等。春季和冬季群落间相异性较大,夏季群落间相异性较小,对相异性影响较大的分歧种主要有葛氏长臂虾、哈氏仿对虾、脊尾白虾和中华管鞭虾等。秋季群落稳定性最好,冬季随着台湾暖流的减弱,浙江沿岸流影响增强[36],水温的降低,大量虾类向外部较深海域进行洄游,迁入量小于迁出量,冬季更替率大于其他季节,群落稳定性最差。
3.5 群落结构特征与环境因子的关系
根据虾类群落与水文环境因子相关性结果分析:周年4个季度月调查所获15种虾类中,4个季节均出现的种类仅有7种,53.33%的虾类是洄游性虾类,而虾类的洄游能够直接影响虾类的分布情况,通过RDA分析,这可能是因为调查海域水深相对较浅,离大陆较近,且处在台湾暖流和浙江沿岸流的交汇区[36],受其交互影响,以及降水、大陆径流等影响,导致底层温度成为影响虾类分布最为重要的环境因子,夏冬2个季节底层温度变化范围较大,分别为1.7℃和0.4℃,虾类分布受底层温度影响显著,特别是冬季,随着温度的降低,虾类开始越冬洄游,各水深带虾类种类数随水深的增加而递增;其次是表层盐度和底层盐度对虾类春季的分布影响较明显,可能是由于调查海域水深较浅,离岸边较近,受台湾暖流和浙江沿岸流,以及降水、大陆径流等因素影响,导致调查海域春季盐度变化较大;而化学需氧量仅在秋季对虾类的分布影响较明显,可能是秋季藻类生物量较多,导致水体化学需氧量含量较低;水深仅在夏季对虾类的分布影响较明显,其他季节其影响较弱。
4 结论
本次调查中所获15种虾类中,主要以广温、广盐性种类为主,4个季节均出现的种类仅有7种,53.33%的虾类是洄游性虾类;从不同水深区域分析,以水深在5~10 m区域虾类种数最多,水深小于5 m的区域虾类种数最少。优势种为脊尾白虾、哈氏仿对虾、中华管鞭虾、细巧仿对虾、中国毛虾、葛氏长臂虾和日本鼓虾等7种。多样性指数以秋季最高,春、夏季次之,冬季最低,且随水深的增加均呈先减少后增加的趋势,在水深5~10 m区域多样性最低。对相似性影响较大的特征种主要有中国毛虾、葛氏长臂虾、脊尾白虾、日本鼓虾、细巧仿对虾和周氏新对虾等,对相异性影响较大的分歧种主要有葛氏长臂虾、脊尾白虾、日本鼓虾、细巧仿对虾、中国毛虾和周氏新对虾等。秋季群落稳定性最好,虾类物种迁入量大于迁出量,夏季群落稳定性较好,虾类物种迁入量和迁出量均处于平衡状态,冬季群落稳定性最差,虾类物种迁入量小于迁出量。通过RDA分析,虾类分布受底层温度影响最大,其次是表层盐度和底层盐度影响较明显。
致谢:本研究中赵盛龙老师、陈健老师帮助种类鉴定,谨致谢忱!
[1] 刘瑞玉.黄海及东海经济虾类区系的特点[J].海洋与湖沼,1959,2(1):35-42.
[2] 刘瑞玉.黄、东海虾类动物地理学研究[J].海洋与湖沼,1963,5(3):230-244.
[3] 王彝豪.舟山沿海经济虾类及其区系特点[J].海洋与湖沼,1987,18(1):48-54.
[4] 宋海棠.东海虾类的生态群落与区系特征[J].海洋科学集刊,2002,44(5):124-133.
[5] 宋海棠,姚光展,俞存根,等.东海虾类的种类组成和数量分布[J].海洋学报,2003,25(S1):171-179.
[6] 俞存根,宋海棠,丁跃平,等.浙江近海虾类资源的初步评估[J].浙江水产学院学报,1994,13(3):149-155.
[7] 卢占晖,薛利建,张龙,等.东海大陆架虾类资源量评估[J].水生生物学报,2013,37(5):855-862.
[8] 齐海明,徐兆礼.椒江口海域春秋季虾类资源分布与环境的关系[J].生态学杂志,2013,32(8):2062-2069.
[9] 俞存根,陈小庆,宋海棠,等.春季东海海域虾类群落结构及其多样性[J].生态学报,2009,29(7):3593-3604.
[10] 陈小庆,俞存根,宋海棠,等.东海中北部海域虾类群聚结构特征及空间分布[J].海洋学研究,2010,28(4):50-58.
[11] 何贤保,俞存根,覃涛,等.舟山渔场及附近海域虾类群落结构特征分析[J].水生生物学报,2013,37(3):553-559.
[12] 夏陆军,陈万东,郑基,等.南麂列岛海洋自然保护区的虾类种类组成和数量分布[J].中国水产科学,2016,23(3):648-660.
[13] 夏陆军,俞存根,蔡厚才,等.南麂列岛海洋自然保护区虾类群落结构及其多样性[J].海洋学报,2016,38(2):73-83.
[14] 晁文春,何贤保,苗振清,等.春夏季南麂列岛海域甲壳类种类组成及分布特征[J].浙江海洋学院学报:自然科学版,2013,32(3):214-221.
[15] 徐捷,陈佳杰,徐兆礼.吕泗渔场沿岸海域春夏季虾类群落特征[J].水产学报,2014,38(8):1097-1105.
[16] 齐海明,孙岳,徐兆礼,等.椒江口海域春秋季虾类群落结构及其影响因素[J].应用生态学报,2013,24(12):3546-3552.
[17] 齐海明.椒江口海域甲壳动物群落结构和资源分布及其与环境因子的关系[D].上海:上海海洋大学,2014:1-54.
[18] 张洪亮,王忠明,朱增军,等.浙江南部沿岸产卵场春季虾类群落结构特征分析[J].水生生物学报,2013,37(4):712-721.
[19] Lobry J,Lepage M,Rochard E.From seasonal patterns to a reference situation in an estuarine environment:example of the small fish and shrimp fauna of the Gironde estuary(SW France)[J].Estuarine,Coastal and Shelf Science,2006,70(1-2):239-250.
[20] Munga C N,Mwangi S,Ong’anda H,et al.Species composition,distribution patterns and population structure of penaeid shrimps in Malindi-Ungwana Bay,Kenya,based on experimental bottom trawl surveys[J].Fisheries Research,2013,147:93-102.
[21] 浙江省玉环县海洋与渔业局.浙江玉环披山海洋特别保护区选划论证报告[R].台州:浙江省玉环县海洋与渔业局,2011.
[22] 中华人民共和国国家质量监督检验检疫总局,中国国家标准化管理委员会.GB/T 12763.6-2007海洋调查规范第6部分:海洋生物调查[S].北京:中国标准出版社,2008:1-157.
[23] 刘瑞玉.胶州湾生态学和生物资源[M].北京:科学出版社,1992.
[24] Brazner J C,Beals E W.Patterns in fish assemblages from coastal wetland and beach habitats in Green Bay,Lake Michigan:a multivariate analysis of abiotic and biotic forcing factors[J].Canadian Journal of Fisheries and Aquatic Sciences,1997,54(8):1743-1761.
[25] Clarke K R.Non-parametric multivariate analyses of changes in community structure[J].Austral Ecology,1993,18(1):117-143.
[26] Field J G,Clarke K R,Warwick R M.A practical strategy for analysing multispecies distribution patterns[J].Marine Ecology-Progress Series,1982,8(1):37-52.
[27] 朱鑫华,吴鹤洲,徐凤山,等.黄渤海沿岸水域游泳动物群落多样性及其相关因素的研究[J].海洋学报,1994,16(3):102-112.
[28] Ter Braak C J F.Canonical correspondence analysis:a new eigenvector technique for multivariate direct gradient analysis[J].Ecology,1986,67(5):1167-1179.
[29] Ter Braak C J F,Prentice I C.A theory of gradient analysis[J].Advances in Ecological Research,1988,18:271-317.
[30] 刘淑德,线薇微,刘栋.春季长江口及其邻近海域鱼类浮游生物群落特征[J].应用生态学报,2008,19(10):2284-2292.
[31] 刘瑞玉.中国海洋生物名录[M].北京:科学出版社,2008,
[32] 俞存根,宋海棠,姚光展.东海蟹类群落结构特征的研究[J].海洋与湖沼,2005,36(3):213-220.
[33] Quetglas A,Carbonell A,Sánchez P.Demersal continental shelf and upper slope cephalopod assemblages from the Balearic Sea(north-western Mediterranean).Biological aspects of some deepsea species[J].Estuarine,Coastal and Shelf Science,2000,50(6):739-749.
[34] 宋海棠,俞存根,薛利建,等.东海经济虾蟹类[M].北京:海洋出版社,2006:83-85.
[35] 曹善茂,宋博,王利明,等.温度、盐度和pH对中华原钩虾呼吸和排泄的影响[J].大连海洋大学学报,2015,30(5):519-523.
[36] 许建平,杨士英.南麂列岛及其附近海域的水文和气候特征[C]//南麂列岛国家级海洋自然保护区论文选(一).北京:海洋出版社,1992:1-9.
[37] 高音,刘明勇,汤勇,等.辽东湾渔业资源及生态环境的调查分析[J].大连海洋大学学报,2013,28(2):211-216.
Community structure and relationship with environment in shrimp in Pishan waters at Yushan fishing ground
XIE Xu, YU Cun-gen, ZHENG Ji, XIA Lu-jun, BI Si-yao, ZHANG Ping, DENG Xiao-yan, MIAO Lu
(Fishery College, Zhejiang Ocean University, Zhoushan 316022, China)
The shrimp community features including species composition, dominance species, and diversity as well as similarity of community structure and relationship between community structure and hydrological environmental factors were evaluated by the data from 15 positions by using bottom trawling around Pishan area in Yushan fish ground in November of 2015 and February, May, and August of 2016.A total of 15 species were found, belonging to 11 genera in 8 families, with dominance species of Exopalaemon carinicauda, Parapenaeopsis hardwickii, Solenocera crassicornis, Parapenaeopsis tenella, Acetes chinensis, Palaemon gravieri and Alpheus japonicus.The shrimp species composition was varied with seasons, resulting in the big difference in the composition of dominant species,with the maximal number of shrimp species in autumn and the minimal number in winter.There was the maximal number of dominant species in spring and the minimum in winter.The shrimp species and number in the catches were changed with water depth, with the most species at depths of 20-30 m and the minimum at depths of 30-40 m.There was lower Shannon-Wierer diversity index of shrimp community in winter than in other seasons, and decline with water depth, less at 5-10 m and more at 5 m.The findings indicate that water depths, temperature and salinity are the main environmental factors influencing species composition and feature of shrimp community structure.
Pishan waters in Yushan fishing ground; shrimp; diversity; community structure; environmental factor
S932.5
A
10.16535/j.cnki.dlhyxb.2017.06.013
2095-1388(2017)06-0713-11
2017-02-24
国家自然科学基金资助项目 (31270527);玉环县海洋与渔业局委托项目 “玉环披山省级海洋特别保护区渔业资源调查与潮间带生物资源调查2015.10—2017.05”
谢旭 (1991—),男,硕士研究生。E-mail:1426619449@qq.com
俞存根 (1960—),男,教授。E-mail:cgyu@zjou.edu.cn
