3种模型对毛竹快速生长期冠层叶片叶绿素荧光—快速光响应曲线(RLCS)拟合的比较
2017-12-25温国胜
朱 弘, 温国胜
(1.浙江农林大学亚热带森林培育国家重点实验室,浙江 临安 311300;2.南京林业大学南方现代林业协同创新中心/生物与环境学院,江苏 南京 210037)
3种模型对毛竹快速生长期冠层叶片叶绿素荧光—快速光响应曲线(RLCS)拟合的比较
朱 弘1,2, 温国胜1
(1.浙江农林大学亚热带森林培育国家重点实验室,浙江 临安 311300;2.南京林业大学南方现代林业协同创新中心/生物与环境学院,江苏 南京 210037)
为定量研究亚热带经济竹种毛竹(Phyllostachysedulis)在大小年快速生长期的叶绿素荧光特性及其光响应规律.以2龄竹、4龄竹和6龄竹的3个竹龄毛竹为材料,采用便携式调制式叶绿素荧光仪测定各竹龄毛竹出笋前期(4月)冠层叶片叶绿素荧光快速响应曲线(RLCS),应用直角双曲线修正模型、双指数方程、非直角双曲线模型拟合,计算对应参数,筛选出最佳的光合响应曲线拟合模型,并进一步测定拟合整个快速生长期(4-7月)3个竹龄的快速响应曲线参数变化.结果表明:直角双曲线修正模型在最大电子(Jmax)、饱和光强(PARsat)、初始斜率(α)及拟合系数(R2)均优于传统双指数方程和非直角双曲线模型,为拟合毛竹叶绿素荧光快速响应曲线(RLCS)最优模型;毛竹叶片光合机制大小年差异显著,大年与小年的光合高峰分别出现在6月和5月,快速生长期内的光合响应能力总体表现为Ⅱ度竹>Ⅰ度竹>Ⅲ度竹.因此,建议在竹林经营中,应集中在大小年光合高峰期间追施肥料,并且注意维持各竹龄的适当立竹结构,以促进毛竹林整体可持续发展.
毛竹; 快速生长期; 林冠层; 叶绿素荧光—快速响应曲线; 直角双曲线修正模型
叶绿素荧光—快速光响应曲线(rapid light curves, RLCS)是在传统荧光淬灭分析技术实践基础上发展起来的快速、无损、便捷的手段,由于它能体现植物光合作用活性及PSⅡ电子传递速率的特点,同时反映光合器官对瞬时及长期生活光照的响应,例如光适应、光抑制及光保护等生理现象,近年来被广泛应用在植物生理研究中[1-4].
毛竹(Phyllostachysedulis)是我国南方分布面积最大,开发利用程度最高,集经济、生态、社会效益于一体的笋材两用竹种[5].毛竹林生长中的大小年现象十分明显,一般以大量出笋的自然年为大年(出笋从3月中旬开始,快速成竹期主要集中在4—5月,成竹展叶期到6月中下旬基本完成);反之为小年(主要进行地下生鞭、地上换叶).毛竹长势高大、光合分层效应显著,异龄毛竹间在冠层空间的光能异质性显著,进而会影响整个竹林的生长更新与林分状况[6-7].
确定植物光合作用的光响应曲线对于研究植物的光合性能具有重要意义.迄今为止,国内外学者建立了许多光响应模型,由于不同模型均存在一定的优缺点,在实际应用中拟合的效果参差不齐[8].张利阳等[9]比较了4 种光响应模型对毛竹光响应曲线的拟合,以直角双曲线模型和非直角双曲线模型拟合精度最高,但各模型仍存在一定缺陷.叶子飘等[10-11]在近年提出了新的Ye氏光响应机理模型,并逐渐推广,为更加准确地分析植物的光合能力提供了有效工具.然而对光响应模拟的研究主要集中在光合、CO2—响应曲线方面[12-15],叶绿素荧光—快速响应曲线的拟合以往多集中于水生植物[16]的应用,适用于陆生单子叶毛竹的响应模型未见报道.因此,本研究基于叶绿素荧光技术,通过测量毛竹RLCS曲线并选用2种经验模型和Ye氏新模型进行拟合比较,旨在筛选最适毛竹的荧光响应模型,并依此模型及其参数定量分析毛竹大小年快速生长期不同月份、不同竹龄对于环境(尤其是光能)的动态适应特征,为毛竹林的科学经营、高效利用提供理论依据.
1 材料与方法
1.1 试验地概况
试验在浙江农林大学竹类研究所毛竹生理生态定位监测站进行,样地位于浙江省临安市青山镇研里村(地理坐标30°14′ N,119°42′ E).地形地貌为低山丘陵,研究样地海拔100~250 m.该区域属亚热带季风气候,年平均气温15.9 ℃,年降水量1 442.0 mm,毛竹林土壤发育于凝灰岩和粉砂岩的红壤土类,土层厚度约60 cm.pH 5.20~5.90,有机质 3.97 g·kg-1,全氮、速效磷、速效钾分别为0.14,4.25和78.16 mg·kg-1.试验区所属为毛竹林地一般经营类型,密度约3 200株·hm-2.林下尚生长少量灌木及草本植物[17].
1.2 试验材料
研究区毛竹林样地采取隔年留养新竹及采伐老竹的方式,依据成竹标记及毛竹换叶规律方法[18]表明现存竹龄结构为2龄竹、4龄竹、6龄竹,并均已钩梢.选取有代表性的西南坡中部地段,搭建定位观测塔.毛竹的冠层位于观测塔的8.5~10.0 m处,荧光测定均在塔上进行.
1.3 叶绿素荧光快速响应曲线(RLCS)的测定
基于样地毛竹生长规律和研究需要,将生长大年(2014年)4—7月均定义为快速生长期,其中进一步划分为快速生长前期(4月23日)、生长中期(5月23日)、生长后期(6月7日)、对照期(7月29日),以及毛竹生长小年(2015年)对应同时期的4月14日、5月20日、6月24日、7月15日,分别展开野外测定.测定当天为晴朗或多云天气,测定时间为9:00—11:00,使用便携式调制式叶绿素荧光仪(PAM-2100,Walz,德国),在光适应阶段即冠层叶自然光照强度下,利用仪器自带光量子探头和热电偶记录光合有效辐射(PAR)及叶表面温度,随机选取3个竹龄林冠的中上部向阳面枝条2龄生成熟健康竹叶,在每张叶片中部避开中脉区域测定毛竹3个竹龄叶片的快速光响应曲线(rapid light curves, RLCS),光化学梯度设定从int 1至int 10,每个梯度光照持续时间为10 s,如此重复,直到达到饱和光强.经检测光和饱和脉冲光测得Ft、Fm′,由此得出PSⅡ的实际量子产量Yield=(Fm′-F)/Fm′,根据ETR=Yield×PAR×0.84×0.5公式[19],计算得到各梯度所对应毛竹叶片的电子传递速率(electron transport rates, ETR).
1.4 数据处理
利用Mircosoft Excel 2007进行数据整理,采用Ye氏模型建立者叶子飘博士提供的光合计算4.1.1软件,对叶绿素荧光快速响应曲线(RLCS)及其参数初始斜率(α)、最大电子(Jmax)、饱和光强(PARsat)、决定系数(R2)的拟合,并利用Origin 8.0软件绘图.拟合软件选取以下3种模型与实测数据进行拟合比较(表1).

表1 3种快速光响应曲线(RLCS)的经验模型Table 1 Three models for rapid light curve (RLCS)
2 结果与分析
2.1 3种快速响应曲线模型拟合的比较
图1是利用3种快速光响应曲线模型分别拟合了毛竹在大年快速生长前期(2014年4月23日)的3个竹龄,即2龄竹、4龄竹、6龄竹,图上直观地可以看到电子传递速率(ETR)随光合有效辐射(PAR)增加而增大,到达光饱和点(PARsat)时所获得最大电子(Jmax);之后,随着光合有效辐射继续增大,2龄竹(图1A)、4龄竹的电子传递速率趋于饱和(图1B),而6龄竹(图1C)由于饱和点出现较早,因而下降显著.

A:2龄竹;B:4龄竹;C:6龄竹.图1 3种快速光响应曲线模型拟合的比较Fig.1 Comparison on degree of fitting of 3 rapid light curves model
表2进一步给出了3种模型拟合曲线所得的参数并与实测值的比较.从表中可以看出用直角双曲线模型拟合对毛竹2龄、4龄、6龄的决定系数(R2)较高,分别达到0.957、0.961、0.968,初始斜率(α)、最大电子(Jmax)、饱和光强(PARsat)各参数拟合结果与实测值也最为接近;双指数方程决定系数次之,但其中,毛竹4龄饱和光强的拟合值2 006.71 μmol·m-2·s-1,比实测值1 141.00 μmol·m-2·s-1高75.81%;非直角双曲线模型除了对6龄竹拟合偏差较大外(R2=0.658),其余参数也较为接近实测值,4龄竹的决定系数(R2=0.979)甚至高于直角双曲线模型(R2=0.961),却不能直接拟合出饱和光强拟合值.
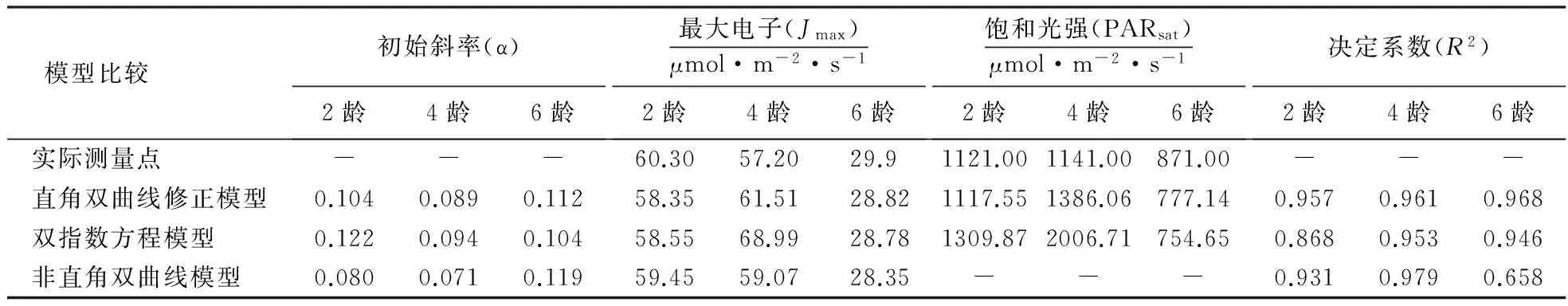
表2 3种模型拟合毛竹冠层叶片快速光响应曲线的拟合值结果与实测值比较Table 2 Comparison on fitted and experimental values of chlorophyll fluorescence characteristics of moso bamboo based on 3 rapid light curves (RLCS)
2.2 直角双曲线修正模型对毛竹快速生长期的拟合
图2列出了采用直角双曲线修正模型拟合后的毛竹不同时期、大小年份的快速响光响应曲线(RLCS)参数动态变化及其比较,在2014年即毛竹生长大年,3个竹龄的初始斜率(α)总体随月份呈现逐渐增大趋势(图2A),其中毛竹4龄、6龄在5月分别获得斜率在各自快速生长期内最大值和次大值;3个竹龄的最大电子(Jmax)在快速生长期内总体呈现先升后降的趋势(图2B),6—7月,降幅显著,均由最高点或者次高点降到最低点;对于饱和光强(PARsat)而言,与初始斜率(α)相反,3个竹龄总体随月份呈现下降趋势,但均在6月获得各自次大或者最大值(图2C).

A:大年初始斜率(α);B:大年最大电子(Jmax);C:大年饱和光强(PARsat);D:小年初始斜率(α);E:小年最大电子(Jmax);F:小年饱和光强(PARsat).图2 毛竹大小年不同时期的快速响光响应曲线(RLCS)参数的比较Fig.2 Comparison of parameters fitted with rapid light curves of moso bamboo in major and minor year
与大年生长相对,毛竹在2015年同期的RLCS各参数的变化如下,3个竹龄的初始斜率(α)峰值均出现在5月,最大值表现为毛竹3龄>1龄>5龄(图2D);3个竹龄的最大电子(Jmax)总体表现为先升后降趋势,峰值也均出现在5月(图2E);毛竹3个竹龄的饱和光强在1龄、3龄表现为先升后降的趋势,但峰值出现的时间不一致,分别出现在5月和6月,而5龄随月份呈逐渐下降趋势(图2F).
3 小结和讨论
光响应曲线的确定,对深入了解植物光反应过程的效率非常重要.尽管国内外学者针对不同植物物种、气候生态环境、植物生理生态等特性建立了多种模型,但还没有一个光响应模型能同时拟合植物在光异质性较大的光响应数据,因而在理论与应用上存在着分歧[22].这其中,以非直角双曲线模型和双指数方程模型应用最为广泛,然而非直角双曲线是一条没有极值的渐近线,不能拟合在PSⅡ动力学下调时的特殊情况,因而无法得到植物在光胁迫环境下的饱和光强(PARsat)参数;而双指数方程在解释光饱和区域时也存在一定的缺陷,往往会出现拟合值远高出实际值的结果,上述模型的不足也在水稻品种[23]、甘薯品种[8]、黄山栾树[24]等不同植物材料的研究中得到了验证,从而阻碍了人们对植物叶片光合作用过程的深入研究.近年来Ye氏直角双曲线修正模型逐渐崭露头角,该模型亦涉及PSⅡ叶绿素分子光能的吸收、激发、退激发及电子传递等过程,并可拟合光抑制部分.在本研究中,笔者也得到了类似的结果,虽然与非直角双曲线模型比较,双指数模型是存在极值的方程,可以拟合出3个竹龄的饱和光强,然而其拟合值相比实测值相差较大,例如在拟合毛竹4龄时的饱和光强比实测值高竟达75.81%,这显然是不合理的.综合比较而言,直角双曲线修正模型与实测值最为接近,无论是最大电子(Jmax)还是决定系数(R2),均优于双指数方程,适合用来拟合毛竹不同竹龄的快速光曲线(表2).
PSⅡ快速光响应曲线RLCS利用ETR随PAR变化规律,可反映植物叶片对光强瞬间变化的适应机制,是研究植物光合生理特性及其适应性的重要手段,在相同光强下ETR越高,为光合作用碳同化所形成的同化力(ATP和NADPH)就越高[25].在毛竹生长大年(2014),初始斜率(α)在毛竹4年生和6年生均在5月份达到高峰,6月受到到抑制,而毛竹2年生表现最佳,在整个快速生长期内稳步增加,总体表现为毛竹2年生>6年生>4年生,说明在相同环境条件下,毛竹2年生对光照的响应能力最强,具备较大的潜在光能;而在最大电子(Jmax)和饱和光强(PARsat)两个参数上,毛竹2年生尤其是4年生均在6月份达到峰值,7月份却显著降低,表明高温迫使毛竹叶片用于非光化学反应的耗散能量比例增大,PSⅡ反应中心电子传递受阻,降低了原初光能转化效率,进而出现了光抑制现象.而毛竹生长小年(2015),RLCS各参数在整个快速生长期内增幅较缓,在初始斜率(α)和最大电子(Jmax)参数上,在5月份出现一个峰值,随后平缓降低.
综合分析比较RLCS各荧光参数,毛竹各竹龄光合作用大小年差异显著,其中在生长大年的光合高峰出现在6月,生长小年光合作用的高峰则推提前至5月,同时,大年在7月出现了明显光抑制现象,小年光抑制不明显.推测这种不同年份间对光照的响应机制差异,对毛竹林大小年的形成机制起着重要的作用,主要是与隔年换叶有关,一般大年毛竹新展叶期在6月结束,而小年毛竹经历了早春换叶期的调整,在5月份基本完成新老叶的集中替换,随后迅速提高了叶片PSⅡ反应中心电子传递能力,即使受到初夏高温胁迫,仍能维持较好的光能转化效率.光合响应总体表现为Ⅱ度竹>Ⅰ度竹>Ⅲ度竹,本研究表明,低龄毛竹在成竹期对自然光照更加敏感,具备较大的光合潜力,中龄毛竹光合能力最强,老龄老竹光合能力最差,因此在竹林经营中应集中在大小年光合高峰期间追施肥料,并且注意维持各竹龄的适当立竹的结构,以促进毛竹林可持续发展.叶绿素荧光—快速响应曲线及直角双曲线修正模型可以作为今后毛竹荧光监测和生长评价的统一标准和有效手段.
致谢:江西井冈山大学数理学院叶子飘教授百忙之中在叶绿素荧光修正模型拟合、软件制图方面提供了大力支持和宝贵意见,在此表示衷心感谢.
[1] RALPH P J, GADEMANN R. Rapid light curves: A powerful tool to assess photosynthetic activity[J]. Aquatic Botany, 2005,82(3):222-237.
[2] 胡文海,张斯斯,肖宜安,等.两种杜鹃花属植物对长期遮阴后全光照环境的生理响应及其光保护机制[J].植物生态学报,2015,39(11):1 093-1 100.
[3] 王春萍,雷开荣,李正国,等.低温胁迫对水稻幼苗不同叶龄叶片叶绿素荧光特性的影响[J].植物资源与环境学报,2012,21(3):38-43.
[4] 马锦丽,江洪,舒海燕,等.天目山自然保护区典型阔叶林的光合特性[J].福建农林大学学报(自然科学版),2016,45(4):381-390.
[5] 应叶青,郭璟,魏建芬,等.水分胁迫下毛竹幼苗光合及叶绿素荧光特性的响应[J].北京林业大学学报,2009,31(6):128-133.
[6] 陈嘉琦,温国胜,王艳红,等.春季毛竹光化学效率空间异质性比较[J].福建林学院学报,2014,34(1):52-56.
[7] 朱强根,金爱武,唐世刚,等.毛竹枝叶生物量的冠层分布对钩梢和施肥的响应[J].中南林业科技大学学报,2015,35(1):24-29.
[8] 张磊,刘维正,辛国胜,等.3种专用型甘薯光合光响应曲线及其模型拟合研究[J].中国农学通报,2015,31(15):71-77.
[9] 张利阳,温国胜,王圣杰,等.毛竹光响应模型适用性分析[J].浙江农林大学学报,2011,28(2):187-193.
[10] YE Z P, SUGGETT D J, ROBAKOWSKI P, et al. A mechanistic model for the photosynthesis-light response based on the photosynthetic electron transport of photosystem Ⅱ in C3 and C4 species[J]. New Phytologist, 2013,199(1):110-120.
[11] 叶子飘.光响应模型在超级杂交稻组合——Ⅱ优明86中的应用[J].生态学杂志,2007,26(8):1 323-1 326.
[12] 闫小红,周兵,王宁,等.基于光响应机理模型的3种草本植物光合特性差异解析[J].南京林业大学学报(自然科学版),2016,40(3):63-69.
[13] 黄滔,刘玮,唐红,等.4个观赏竹种的光合特性及其影响因子分析[J].植物资源与环境学报,2016,25(1):24-33.
[14] 叶子飘,高峻.光响应和CO2响应新模型在丹参中的应用[J].西北农林科技大学学报(自然科学版),2009,37(1):129-134.
[15] 李力,张祥星,郑睿,等.夏玉米光合特性及光响应曲线拟合[J].植物生态学报,2016,40(12):1 310-1 318.
[16] 王锦旗,宋玉芝,薛艳.紫外辐射对菹草(Potamogetoncrispus)成株快速光响应曲线的影响[J].湖泊科学,2015,27(3):451-458.
[17] 袁佳丽,温国胜,张明如,等.毛竹快速生长期的水势变化特征[J].浙江农林大学学报,2015,32(5):722-728.
[18] 朱弘,温国胜.不同竹龄毛竹冠层叶片PSⅡ叶绿素荧光特性的比较[J].中南林业科技大学学报,2017,37(1):12-19.
[19] 张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[20] PLATT T, GALLEGOS C L, HARRISON W G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton[J]. Publications-Astronomical Society of Japan, 1980,57(57):341-345.
[21] ZHAO D L, REDDY K R, KAKANI V G, et al. Leaf and canopy photosynthetic characteristics of cotton (Gossypiumhirsutum) under elevated CO2concentration and UV-B radiation[J]. Journal of Plant Physiology, 2014,161(5):581-590.
[22] 刘强,李凤日,谢龙飞.人工长白落叶松冠层光合作用—光响应曲线最优模型[J].应用生态学报,2016,27(8):2 420-2 428.
[23] 闫小红,尹建华,段世华,等.四种水稻品种的光合光响应曲线及其模型拟合[J].生态学杂志,2013,32(3):604-610.
[24] 叶子飘,康华靖,陶月良,等.不同模型对黄山栾树快速光曲线拟合效果的比较[J].生态学杂志,2011,30(8):1 662-1 667.
[25] 钱永强,周晓星,韩蕾,等.3种柳树叶片PSⅡ叶绿素荧光参数对Cd2+胁迫的光响应[J].北京林业大学学报,2011,33(6):8-14.
ThecomparisonofthreemodelsofPhyllostachysedulisincanopyduringthefastgrowthperiodfittingwithPSⅡchlorophyllfluorescencerapidlightcurves(RLCS)method
ZHU Hong1,2, WEN Guosheng1
(1.State Key Laboratory of Subtropical Silviculture, Zhejiang Agriculture and Forestry University, Lin′an, Zhejiang 311300, China; 2.Co-Innovation Center for Sustainable Forestry in Southern China/College of Biology and the Environment, Nanjing Forestry University, Nanjing, Jiangsu 210037, China)
To study the chlorophyll fluorescence characteristics of moso banmboo (Phyllostachysedulis) quantitatively and its light response mechanism, moso bamboo at different growth stages, including 2, 4, 6 years old, was measured by portable fluorescence measurement system at early shooting stage in April. Then rapid light curves (RLCS) based on previous figure were matched by modified rectangular hyperbolic model, nonrectangular hyperbolic model and ambiexponent equation to screen the optimum model. The results showed that modified rectangular hyperbolic model was the optimum model in terms of maximum rate of photosynthetic elactron transport (Jmax), saturation intensity (PARsat) and coefficient of determination (R2). Photosynthetic mechanism of bamboo varied significantly between on-year and off-year, with the maximum photosynthesis rate being in June for on-year and in May for off-year. General photosynthetic response ability in rapid growth period in a descending order were degreeⅡ, degreeⅠ, and degreeⅢ. We suggest fertilization during the peaks of photosynthesis in on-year and off-year respectively, and maintaining a sustainable structure of bamboo at different ages.
Phyllostachysedulis; rapid growth period; canopy layer; chlorophyll fluorescence-rapid light curve; modified rectangular hyperbolic model
2017-02-07
2017-03-15
国家自然科学基金(31270497、30972397).
朱弘(1990-),男,博士研究生.研究方向:植物生态学.Email:1107401987@qq.com.通讯作者温国胜(1959-),男,教授.研究方向:植物生理生态学.Email:695708715@qq.com.
Q945
A
1671-5470(2017)06-0659-06
10.13323/j.cnki.j.fafu(nat.sci.).2017.06.010
(责任编辑:吴显达)
