不同倍性滇杨叶片解剖结构差异分析
2017-12-19辛亚龙唐军荣何承忠辛培尧
邱 芬 ,辛亚龙 ,唐军荣 ,李 斌 ,何承忠 ,许 程 ,辛培尧
(1.西南林业大学 国家林业局西南地区生物多样性保育重点实验室,云南 昆明 650224;2. 西南林业大学 云南省高校林木遗传改良与繁育重点实验室,云南 昆明 650224)
不同倍性滇杨叶片解剖结构差异分析
邱 芬1,2,辛亚龙1,2,唐军荣1,2,李 斌1,2,何承忠1,2,许 程2,辛培尧1,2
(1.西南林业大学 国家林业局西南地区生物多样性保育重点实验室,云南 昆明 650224;2. 西南林业大学 云南省高校林木遗传改良与繁育重点实验室,云南 昆明 650224)
以前期诱导获得的5株多倍体滇杨一年生无性系植株为材料,二倍体滇杨为对照,对其叶片进行石蜡包埋、切片,观察其解剖结构差异,测定叶片厚度、栅栏组织厚度、海绵组织厚度,分析多倍化后滇杨叶片的变异特性。结果表明:不同倍性滇杨在叶片解剖结构各性状间均存在不同表现。不同个体间叶片厚度、CTR、SR、上表皮相对厚度、下表皮相对厚度间均存在显著差异,滇杨经多倍化处理后能引起栅栏组织、海绵组织的变化,表现为基因的剂量效应差异。同时,不同倍性滇杨叶片CTR、SR的变化并不与滇杨的倍性成正相关趋势,这为滇杨遗传改良提供了选择的物质基础。研究结果可为滇杨的抗性育种相关性状的早期选择提供理论依据。
滇杨;多倍体;解剖结构;差异
滇杨Populus yunnanensis属杨柳科Salicaceae杨属Populus,又名云南白杨、白杨树、东川杨柳、白泡桐等[1]。滇杨是我国特有的青杨派树种,主要分布于云南中部和北部、四川西南部和贵州西部等地,海拔范围在1 300~3200 m,是我国西南地区特有的乡土树种之一 。滇杨具有速生、耐寒、适应性强、易无性繁殖、抗叶斑病和叶锈病等特性,滇杨与杨属其他树种相比生长更加迅速,在我国低纬度高海拔地区具有良好的应用前景[2-3]。
但是,滇杨的自然分布比较分散,优良成片林分较少,基础性研究较为薄弱。另外,滇杨还有易受杨树黑斑病、溃疡病及蛀干害虫等危害的弊端,致使其优良特性不能得到充分的发挥和利用[4]。目前,滇杨群体以雄株为主,雌株几乎没有,可利用的遗传资源相对较少,这在一定程度上影响了滇杨不良性状的遗传改良。多倍体育种是有效创造植物新种质,改良植物不良性状的方法之一。由于基因剂量的增加,林木多倍体较其二倍体通常表现抗性方面的优势[5-6]。大量研究表明,多倍体植物在低温、高温、干旱、盐胁迫、病害等逆境下,具有更强的竞争力和生存能力[7-11]。
植物叶片是反映植物对环境适应的最敏感的器官,运用解剖学方法,研究植物叶片的解剖结构及其变化规律已成为逆境生理研究的一个重要方面[12-13]。对前期研究所得的5个多倍体滇杨无性系叶片进行石腊切片,并以二倍体滇杨为对照,显微观察并分析不同无性系叶片解剖结构的特征差异,可为滇杨高抗性种质的选育提供理论依据。
1 材料与方法
1.1 试验材料
采集西南林业大学树木园中叶龄基本一致的5株多倍体滇杨(h1、h2、h3、h4、h5)一年生枝条的叶片,和普通二倍体材料叶片(CK),以叶主脉为中线切为长或宽约为1.0 cm的方块,并放入含有FAA固定液的透明小瓶中固定,固定时间不少于48 h,备用。
1.2 试验方法
1.2.1 石蜡切片
试验采用杨捷频[14]、陈丽莉等[15],提出的试验方法,对处理好的叶片进行石蜡切片。
1.2.2 封片及观察
用中性树胶封片后,将切片放入37℃恒温箱中烘干,数日后观察叶片栅栏组织、海绵组织、上表皮、下表皮、叶主脉和叶片厚度,并进行拍照。用系统自带软件测量相关数据。
1.2.3 统计分析
所有数据利用SPSS17.0软件进行分析。同时,计算叶片组织结构的紧密度(CTR)和疏松度(SR),计算公式如下:
叶片组织结构紧密度(CTR)=(栅栏组织厚度/叶片厚度)×100%;
叶片组织结构疏松度(SR)=(海绵组织厚度/叶片厚度)×100%;
上表皮相对厚度=(上表皮厚度/叶片厚度)×100%;
下表皮相对厚度=(下表皮厚度/叶片厚度)×100%。
2 结果与分析
2.1 二倍体滇杨叶片横切解剖结构观察
二倍体滇杨叶片横切解剖结构(图1,见封二)。由图1可知,二倍体滇杨叶片上、下表皮均由1层细胞构成,且有明显的细胞差别,上表皮排列整齐,下表皮排列不整齐。叶肉细胞有明显的海绵组织和栅栏组织的分化,海绵组织多为椭圆形或不规则形状,栅栏组织为圆柱体,叶片的厚度为309.895±26.084 μm。二倍体滇杨叶片在主脉附近的叶肉细胞明显大于周围的叶肉细胞,且靠近主脉的表皮细胞近圆形,其余的表皮细胞多为狭长形。主脉的直径为2 606.663±423.963 μm(图2,见封二)。
2.2 多倍体滇杨h1叶片横切解剖结构观察
多倍体滇杨h1叶片的形态,叶片上、下表皮由1层细胞组成,靠近上表皮的部分地方存在一层复表皮,栅栏组织分2层,且多为棍棒状,排列整齐紧实,靠近下表皮的叶肉细胞大小不一,多为圆形或者椭圆形且排列不整齐。叶片厚度为371.801±32.387 μm(图3,见封二)。多倍体滇杨h1叶片主脉周围上表皮和下表皮的细胞大小相似,近圆形。下表皮比上表皮厚。主脉直径为1 988.976±62.828 μm(图4,见封二)。
2.3 多倍化体滇杨h2叶片横切解剖结构观察
多倍体滇杨h2叶片横切解剖结构如图5(见封二)所示。图5表明:多倍体滇杨h2叶片上、下表皮由1层细胞组成,靠近上表皮有的地方存在一层复表皮,且叶肉细胞多为椭圆形,靠近下表皮的叶肉细胞大小不一,多为不规则形且排列不整齐。叶片厚度为409.583±36.417 μm。多倍体滇杨h2叶片主脉周围上表皮和下表皮的细胞大小相似。主脉直径为1 161.459±439.040 μm(图6,见封二)。
2.4 多倍体滇杨h3叶片横切解剖结构观察
多倍体滇杨h3叶片的形态,叶片上、下表皮由1层细胞组成,靠近上表皮未发现有复表皮的存在,栅栏组织不分层,且栅栏组织细胞多为棍棒状,靠近下表皮的海绵组织细胞大小不一,多为不规则形且排列不整齐。叶片厚度为269.623±51.033 μm(图7,见封三)。图8(见封三)为多倍体滇杨h3叶片主脉横切解剖结构。图8显示,该材料叶片主脉周围上表皮比下表皮的细胞小,上表皮细胞紧实。主脉直径为1 073.874±282.998 μm。
2.5 多倍体滇杨h4叶片横切解剖结构观察
多倍体滇杨h4叶片横切解剖结构如图9所示。由图9(见封三)可以看出,该材料叶片上、下表皮由1层细胞组成,靠近上表皮的部分存在一层复表皮,栅栏组织分为2~3层,棍棒状,靠近下表皮的海绵组织大小不一,且海绵组织细胞多为椭圆形,大多呈不规则形状且排列不整齐。叶片厚度为333.336±17.027 μm。多倍体滇杨h4叶片的主脉周围上表皮和下表皮的细胞大小相似,椭圆或者圆形,下表皮比上表皮厚。主脉直径为2 570.191±383.251 μm(图10,见封三)。
2.6 多倍体滇杨h5叶片横切解剖结构观察
多倍体滇杨h5叶片上、下表皮由1层细胞组成,且不存在复表皮。靠近上表皮的栅栏组织分2层,为棍棒状,靠近下表皮的海绵组织细胞大小不一,海绵组织细胞大多为椭圆形,且形状不规则排列不整齐。叶片厚度为359.536±8.981 μm(图11,见封三)。多倍体滇杨h5叶片横切主脉解剖结构如图12(见封三)所示。图12表明,该材料主脉周围上表皮和下表皮的细胞大小相似,分布均匀,直径为2 732.666±259.164 μm。
2.7 不同倍性滇杨叶片解剖结构差异分析
不同倍性滇杨叶片的紧密度、疏松度、上表皮相对厚度及下表皮相对厚度差异分析结果列于表1。
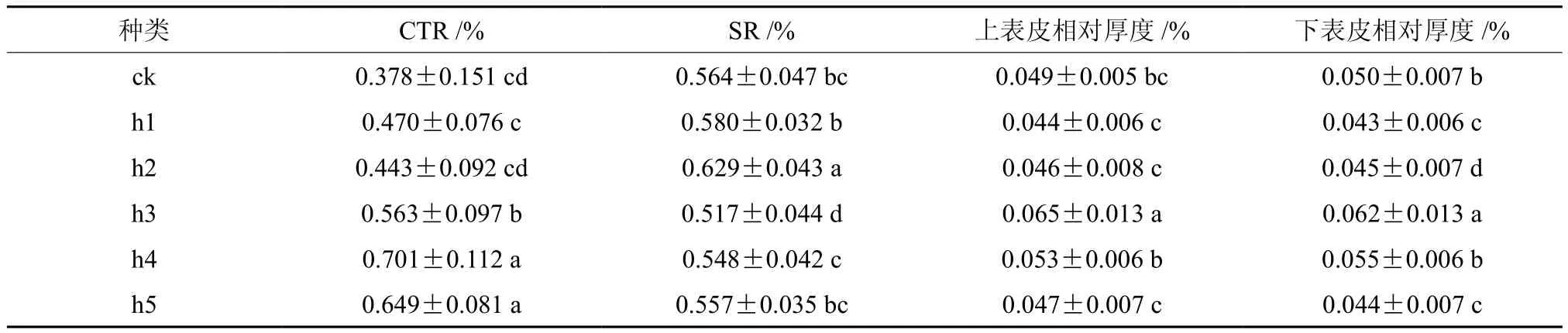
表1 不同倍性滇杨叶片解剖结构特征差异分析†Table 1 Analysis of leaf anatomical structure differences in different ploidy Populus yunnanensis
由表1可知,不同倍性滇杨在CTR、SR、上表皮相对厚度,下表皮相对厚度这4个性状间均存在显著差异。
其中,多倍体材料h3的上表皮和下表皮相对厚度为所有测试材料中最高(分别为6.5%和6.2%)且与其他处理均存在显著差异,而多倍体材料h2、h5与二倍体对照的上表皮相对厚度并无显著差异,多倍体材料h2、h4与二倍体对照的下表皮相对厚度无显著差异。多倍体材料h4和h5的CTR值相对较高,分别为70.1%和64.9%,而以多倍化材料h2和对照较低(分别为44.3%和37.8%),这说明,滇杨经多倍化处理后,并不一定引起叶片紧实度的降低;多倍体材料h2的SR值最高,而h3的最低,分别为62.9%和51.7%,多倍化材料h1、h5和对照之间在SR方面无显著差异,同样表明,多倍化处理滇杨也不一定引起叶片疏松度的增加。可见,滇杨经多倍化处理后,其CTR、SR、上表皮相对厚度、下表皮相对厚度这4个性状间的变化是多样的,表现出植物染色体加倍所引起变异的随机性。
3 讨 论
对滇杨进行加倍处理之后,其叶片解剖结构与二倍体对照相比较发生了显著变化。不同倍性滇杨在CTR、SR、上表皮相对厚度、下表皮相对厚度这4个性状间均存在显著差异。
多倍化材料h3的上表皮和下表皮相对厚度均在所有材料中最高(分别为6.5%和6.2%)且与其他处理均存在显著差异。这种表皮相对厚度的变化证实了李凤兰等[16]、唐军荣等[17]、蔡冬元等[18]等的观点,他们均发现多倍化处理能引起植物在叶片面积、厚度等方面的巨大化。然而,这种变化并不是绝对的,由于基因组剂量效应的复杂性,不同的表型性状可能在不同材料中存在截然不同的变化,这与供试材料基因型的不同和生长环境各异有一定关联。而本研究也表明,并不是所有的多倍体滇杨与二倍体在叶片厚度方面存在显著差异。罗向东等[19]在黄瓜属间不同倍性多倍体形态的研究中也表明了这个观点。杨桂英等[20]在探讨不同倍性山茶花的耐寒潜力时发现,在栅栏组织细胞长度、海绵组织和叶片厚度等性状上,并非所有的多倍体都优于二倍体。李赟[21]在果树多倍体倍性鉴定的研究中发现,在苹果、梨、草莓的不同倍性植株叶片解剖结构中,虽然栅栏组织细胞长度上有增加,在叶片厚度和海绵组织厚度这两个性状方面则并没有显著差异。因此认为多倍化后的植物材料在叶片及栅栏组织的变厚并不是绝对的。同样,国内学者王君[22]在对青杨派树种进行多倍体诱导研究时,也发现了这种现象。这与本研究的结果相一致。这些变异的多样性,为植物的人工进化提供了丰富的物质基础。
植物表型差异是识别植物遗传差异的一个快速通道,尤其在短期内评价无性系植物变异种质资源时,更为快捷。植物叶片形态结构与植物的新陈代谢紧密相关,因此可利用相关指标替代遗传改良时的目标性状进行早期选择。CTR和SR在多种林木上被用来指示耐寒性的强弱,一般CTR与耐寒性呈正相关,SR则呈负相关[23]。王泽华等[24]利用CTR、SR以及栅海比等指标,对不同生境下新疆野苹果5个自然居群的抗寒性进行了评价。史刚荣[25]研究了7种常绿阔叶植物叶片解剖结构后也认为,海绵组织与栅栏组织一般都与植物的抗旱和抗寒性有关,在不同的选择压下可形成多种不同的生态类型。李欢等[26]认为,在旱生环境下,一般植物叶片小,角质膜厚,表皮和蜡被比较发达,有明显的栅栏组织。试验中,多倍化处理后的叶片除h2外在CTR值上均高于二倍体,有明显增幅。王怡[27]的研究表明栅栏组织密度越大,植物的抗虫性就越强。因此,抗虫性与紧密度成正相关,由于刺吸类蚜虫的口针要穿过叶表皮细胞的一定部位摄取食物,以获取生长发育和存活必需的营养物质,而多倍体滇杨致密的上、下表皮和栅栏组织可能对刺吸类蚜虫的口针刺入构成机械障碍,从而使其具备较高的抗虫性。李海英等[28]的研究表明,栅栏组织排列整齐、紧密,而且层数相对较多,可以抵抗病菌的侵入和扩展,成为抗病的又一个结构屏障。纪楠楠[29]发现,在重金属胁迫下,8种观赏树木叶片栅栏薄壁细胞组织厚度逐渐变厚,海绵薄壁细胞组织逐渐分化、栅栏组织海绵组织的比值越来越大。所以,从理论上讲,多倍体滇杨的抗病虫的能力应比二倍体的强。该指标是否可以作为多倍体滇杨抗寒性和抗虫性增强的早期选择标记,还需进一步的研究来佐证。同时,研究还表明,经处理后的多倍体滇杨在CTR 和SR方面的增减并不与其倍性的大小呈正相关,这种变异同样为诱变材料后代的选择提供了较为广泛的基础材料。
[1]中国科学院中国植物志编辑委员会.中国植物志第二十卷第二分册[M].北京:科学出版社,1984.
[2]何承忠,车鹏燕,周修涛,等.滇杨基因资源及其研究概况[J].西南林学院学报,2010,30(1):83-88.
[3]纵 丹,员 涛,周安佩,等.滇杨优树遗传多样性的AFLP分析[J].西北林学院学报,2014,29(4):103-108.
[4]辛培尧,刘 岩,段安安,等.滇杨不同外植体分化培养[J].云南农业大学学报,2011,26(6):828-832.
[5]康向阳,王 君.杨树多倍体诱导技术研究[M].北京:科学出版社,2010.
[6]辛培尧,陈 杰,唐军荣,等.滇杨多倍体的诱导研究[J].中南林业科技大学学报,2014,34(3):24-28.
[7]魏 望,施富超,王东玮,等.多倍体植物抗逆性研究进展[J].西北植物学报,2016,36(4):846-856.
[8]黄金艳,付金娥,覃斯华,等.水分胁迫对二、四倍体薄皮甜瓜苗期生理生化特性的影响[J].安徽农业科学,2011,39(11):6305-6307,6355.
[9]王园园,尹翠翠,王广东.高温胁迫对二倍体和四倍体杂交兰相关生理生化特征的影响[J].西北植物学报,2013,33(4):735-740.
[10]王疆江.四倍体刺槐的组培快繁及抗盐性研究[D].哈尔滨:东北林业大学,2009.
[11]张 弛.红阳猕猴桃四倍体诱导及其抗溃疡病特性初探[D].重庆:西南大学,2011.
[12]康萨如拉,牛建明,张 庆,等.短花针茅叶片解剖结构及与气候因子的关系[J].草业学报,2013, 22(1):77-86.
[13]潘存娥,田丽萍,李贞贞,等.5种杨树无性系叶片解剖结构的抗旱性研究[J].中国农学通报,2011, 27(2):21-25.
[14]杨捷频.常规石蜡切片方法的改良[J].生物学杂志,2006,23(1): 45-46.
[15]陈丽莉,梁文斌,吴思政,等.短梗大参解剖学特性及生态学意义[J].经济林研究,2016,34(4):148-151.
[16]李凤兰,张志毅,张民侠.白杨染色体加倍技术研究及三倍体育种(Ⅲ)──加倍体某些形态特征的观察[J].北京林业大学学报,1994,16(2):15-18.
[17]唐军荣,李 斌,朱丽娜,等.滇杨多倍体苗期叶片形态及光合生理比较分析[J].林业科学研究,2016,29(1):103-109.
[18]蔡冬元,林继华,邓建平.戊炔草胺(propyzamide)作用于刺葡萄的倍性诱变研究[J].中南林业科技大学学报,2016,36(7):103-108.
[19]罗向东,戴亮芳,钱春桃,等.黄瓜属不同倍性异源多倍体的形态及生理特性分析[J]. 西北植物学报,2008,28(3):542-546.
[20]杨桂英,王兵益,何 瀚,等.从叶片解剖结构探讨云南山茶不同倍性的耐旱潜力[J].西南农业学报,2015, 28(6):2714-2719.
[21]李 赟.果树多倍体倍性鉴定方法的研究[D].泰安:山东农业大学,1997.
[22]王 君.青杨派树种多倍体诱导技术研究[D].北京:北京林业大学,2009.
[23]何小勇.翅荚木种源遗传多样性及其抗低温胁迫能力研究[D].长沙:中南林业科技大学,2007.
[24]王泽华,秦 伟,闫娟娟,等.不同生境新疆野苹果叶片解剖结构及其与抗寒性的关系[J].经济林研究,2016,34(3):163-168.
[25]史刚荣.七种阔叶常绿植物叶片的生态解剖学研究[J].广西植物,2004,24(4):334-338.
[26]李 欢,樊军锋,高建社,等.黑杨叶片旱生结构的比较[J].西北林学院学报,2013,28(3):113-118.
[27]王 怡.三种抗旱植物叶片解剖结构的对比观察[J].四川林业科技,2003,24(1):64-67.
[28]李海英,刘亚光,杨庆凯.大豆叶片结构与灰斑病抗性的研究Ⅱ.大豆叶片组织结构与灰斑病抗性的关系[J].中国油料作物学报,2002,24(2):58-60.
[29]纪楠楠.重金属胁迫对几种植物叶片解剖构造的影响[D].哈尔滨:东北林业大学,2012.
Analysis of leaf anatomical structure differences in Populus yunnanensis with different ploidy
QIU Fen1,2, XIN Yalong1,2, TANG Junrong1,2, LI Bin1,2, HE Chengzhong1,2, XU Cheng2, XIN Peiyao1,2
(1.Key Laboratory of Biodiversity Conservation in Southwest Region of State Forestry Administration, Southwest Forestry University,Kunming 650224, Yunnan, China; 2. Key Laboratory for Forest Genetic and Tree Improvement & Propagation in Universities of Yunnan Province, Southwest Forestry University, Kunming 650224, Yunnan, China)
Leaves from annual clone individual plant of fi ve polyploid Populus yunnanensis and a diploid plant, as control, were used to embed with paraf fi n and slice, in order to observe the leaves anatomical structure. The leaf thickness, palisade tissue thickness and spongy tissue thickness were measured to analyze the difference of P.yunnanensis with different ploidy. The results showed that different ploidy P.yunnanensis had the different representation on traits of the leaf anatomical structure. There were signi fi cant differences in leaf thickness, CTR, SR, relative thickness of epicuticle, and relative thickness of lower epidermis of P.yunnanensis in different individuals,which indicated that polyploidy could make the palisade tissue and spongy tissue changed, because of gene dose effect. At the same time. The changes on CTR and SR in leaves of P.yunnanensis were not positive correlation with the their ploidy, which offered the selective material basis of P.yunnanensis breeding. Theoretical basis on the early selection of P.yunnanensis resistance breeding could be provided from these results.
Populus yunnanensis; polyploid; anatomicale structure; differences
S718.47
A
1673-923X(2017)06-0056-04
10.14067/j.cnki.1673-923x.2017.06.009
2016-12-20
云南省教育厅科学研究基金重大专项项目(ZD2015014);西南林业大学科技创新基金(C16006);西南林业大学科研基金项目(XL21611);西南林业大学林学院林学一级学科后备人才计划项目;云南省林学一级学科博士点建设项目
邱 芬,硕士研究生
辛培尧,副教授,博士;E-mail:xpytgyx@163.com
邱 芬,辛亚龙,唐军荣,等. 不同倍性滇杨叶片解剖结构差异分析[J].中南林业科技大学学报,2017, 37(6): 56-59, 69.
[本文编校:吴 彬]
