SHU9119对小鼠发情周期中Ghrelin表达的影响
2017-11-09王彩云范奎奎张召议杜晨光
王彩云 , 潘 登 , 范奎奎 , 张召议 , 李 婷 , 李 强 , 杜晨光,2
(1.内蒙古农业大学兽医学院 , 内蒙古 呼和浩特 010018 ; 2.内蒙古农业大学职业技术学院 , 内蒙古 包头 014109)
SHU9119对小鼠发情周期中Ghrelin表达的影响
王彩云1, 潘 登1, 范奎奎1, 张召议1, 李 婷1, 李 强1, 杜晨光1,2
(1.内蒙古农业大学兽医学院 , 内蒙古 呼和浩特 010018 ; 2.内蒙古农业大学职业技术学院 , 内蒙古 包头 014109)
为了研究SHU9119与ghrelin之间的潜在的生理学关系,本试验选择4周龄的C57BL/6小鼠下丘脑和卵巢组织为研究材料,对小鼠发情周期中SHU9119介导的ghrelin mRNA在下丘脑和卵巢部位表达水平进行研究。结果表明,正常生理条件下小鼠间情期下丘脑保持较高水平的ghrelin mRNA表达,而在卵巢内其表达水平相对较低;腹腔注射SHU9119 (50 μg/kg体重)可极显著抑制下丘脑中ghrelin mRNA的表达(P<0.01),同时显著促进小鼠间情期卵巢Ghrelin mRNA的表达(P<0.05)。然而,SHU9119并未引起小鼠发情前期、发情期和发情后期ghrelin mRNA表达量的显著差异。上述结果表明,SHU9119可能通过MC4R系统调节间情期小鼠ghrelin的变化,而不参与其他性周期ghrelin mRNA表达量的相关调控。
小鼠 ; 下丘脑 ; 卵巢 ; Ghrelin ; SHU9119
作为胃肠道产生的28氨基酸多肽类激素[1],Ghrelin为生长激素促分泌素受体(GHSR)的内源性配体,其在促进生长激素(GH)分泌的同时,也可直接调控能量代谢和能量平衡,如促进脂类沉积,体重增加[2]。SHU9119作为一种人工合成的黑素皮质素4型受体(Melanocortin-4 receptor, MC4R),在研究MC4R和能量代谢方面被广泛使用[3]。
研究结果表明,MC4R基因缺失小鼠表现出食欲旺盛、体重增加和肥胖等症状[4],而且肥胖后生殖能力的严重下降也引起了人们广泛的关注[5],例如,除Ghrelin能量平衡效应外,实验证据表明其可直接影响卵巢生理活动[6]。Ghrelin在生长、能量代谢和生殖之间存在着复杂的生物学功能,且其功能性受体GHS-R1a广泛分布于下丘脑-垂体-性腺轴[7],提示ghrelin可能是通过下丘脑-垂体-性腺轴调节哺乳动物生殖调控的关键因子。然后研究表明,ghrelin不能刺激MC4R基因敲除小鼠的采食量增加,提示ghrelin的作用部分是通过下丘脑黑皮质素受体系统作用[8]。同时ghrelin可刺激正常小鼠采食行为并促进下丘脑黑皮质素受体的表达。
SHU9119作为MC4R特异的受体撷抗剂,可干扰MC4R的采食调控作用[9]。基于ghrelin受体-GSHR与MC4R均为7次跨膜藕联蛋白[10],均与能量代谢息息相关,生殖与能量代谢之间具有紧密关系。因此,本试验通过对不同性周期的雌性小鼠腹腔注射SHU9119,以下丘脑和卵巢为试验材料,通过RT-qPCR技术探究SHU9119对下丘脑和卵巢中ghrelin mRNA水平表达的影响,为研究SHU9119参与雌性小鼠能量代谢和生殖提供数据。
1 材料与方法
1.1 材料获取 雌性C57BL/6 (4周龄)小鼠(n=30),购自北京维通利华公司。试验前一周每天定时进行腹腔注射生理盐水,避免试验时应激。使用阴道涂片法将其分为发情前期、发情期、发情后期和间情期4组,选择体重和健康状况相同的雌鼠共24只,每组各6只。 其中,3只用于生理盐水注射,3只用于SHU9119(50 μg/kg体重)的腹腔注射。注射后1 h,脊椎脱臼法处死以获取下丘脑和卵巢材料,保存于-80 ℃备用。
1.2 小鼠性周期鉴定 选取体重健康状态相近的小鼠(n=24),使用50 μL室温的生理盐水对小鼠的阴道上皮细胞进行冲洗,之后将小鼠的阴道上皮细胞滴加在防脱载玻片上,待液体自然风干后,滴加95%的乙醇进行固定10 min,之后使用蒸馏水进行冲洗。自然风干后向其中滴加1%龙胆紫溶液,作用10 min,冲洗干净后使用光学显微镜进行观察(奥林巴斯XSP-2C)。
1.3 RNA提取和反转录 采用天根生化科技(北京)有限公司RNA提取试剂盒,按说明书提取RNA。反转录直接将6.5 μL的RNA加入到反转录体系中合成cDNA。反应体系为10 μL,其中5×PrimeScript RT Master Mix 2 μL(TaKaRa DRR036)。于PCR仪进行反转录,反应条件为:37 ℃ 15 min,4 ℃。cDNA保存于-20 ℃,并避免反复冻融。
1.4 实时定量PCR (RT-qPCR) 基因序列及引物信息如下:Ghrelin (163 bp, GenBank accession no. NM_021488.4)上游5′-CAAGAAGCCACCAGCTAA ACT-3′,下游5′-GCCATGCTGCTGATACTGA-3′ ;GAPDH (294 bp, GenBank accession no. XM_004438729.1) 上游5′-AAGGGTGGAGCCAAAAGG-3′ ;下游5′-GGATGCAGGGATGATGTTCT -3′,上海生工生物工程技术服务有限公司合成。qPCR反应体系为20 μL,其中荧光酶10 μL、上下游引物各0.4 μL(10 μmol/L)、cDNA 2 μL、dH2O 7.2 μL。扩增条件为:94.0 ℃预变性1 min;94.0 ℃变性20 s,64.0 ℃ghrelin/58 ℃GAPDH退火20 s,72.0 ℃延伸20 s,40个循环。相对Ghrelin mRNA表达量采用2-ΔCT公式进行计算,ΔCT = CTGhrelin-CTGAPDH。
1.5 数据处理 试验数据利用统计学软件GraphPad Prism 5.0的统计程序进行T test统计分析。数据采用平均值±标准误(Mean ± S.E.M)进行表示。
2 结果
2.1 小鼠阴道涂片上皮细胞变化特点 试验观察基本总结见表1和图1。

表1 小鼠阴道涂片上皮细胞变化特点
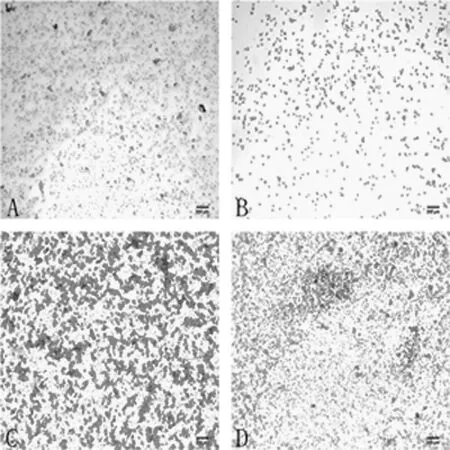
图1 不同发情时期阴道上皮细胞变化 (100×)A:发情前期; B:发情期; C:发情后期; D:间情期
2.2 PCR产物特异性 扩增效率曲线表明扩增效果良好,经熔解曲线分析可知ghrelin和GAPDH基因荧光定量PCR扩增产物在85 ℃和86.5 ℃的熔解温度处出现单一峰。且凝胶电泳结果显示,ghrelin和GAPDH PCR产物分别得到了单一的163 bp和294 bp条带,故产物大小及预期TM值与设计一致(图2)。
2.3 小鼠发情周期内下丘脑和卵巢ghrelin mRNA表达量 从图3可知,正常生理情况下,下丘脑ghrelin mRNA呈逐渐升高趋势,以间情期最高,发情后期次之。腹腔注射SHU9119后下丘脑中ghrelin mRNA水平表达量较对照组在发情后期略有下降,但在间情期呈显著的下降趋势(P<0.01),而发情前期和发情期较生理盐水组均未见显著性差异。
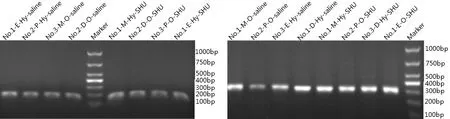
图2 Ghrelin的荧光定量PCR电泳结果(左)和GAPDH的荧光定量PCR电泳结果(右)注:E-Estrus; P-proestrus; M-metaestrus; D-diestrous; Hy-Hypothalamus; O-Ovary
卵巢正常生理条件下,Ghrelin mRNA呈先低后高,至间情期最低的趋势,发情前期、发情期和发情后期未见显著性差异。因试验重复差异相对较大,未体现出统计差异,但与小鼠,大鼠,绵羊,猪等哺乳动物卵巢Ghrelin表达趋势一致。然而,SHU9119处理组较对照组在间情期可显著促进卵巢中Ghrelin mRNA水平表达(P<0.01)。
ghrelin mRNA在间情期正常小鼠卵巢中的表达水平与下丘脑相反,其保持较低的表达水平。SHU9119处理后,ghrelin mRNA在下丘脑和卵巢均出现了极显著的逆转(P<0.01)。其中,SHU9119处理后的下丘脑中Ghrelin mRNA表达水平与正常生理条件下间情期卵巢的表达量接近,均处于较低的水平;SHU9119处理后的卵巢中Ghrelin mRNA表达水平与正常生理条件下间情期小鼠下丘脑的表达量接近,均处于较高水平。
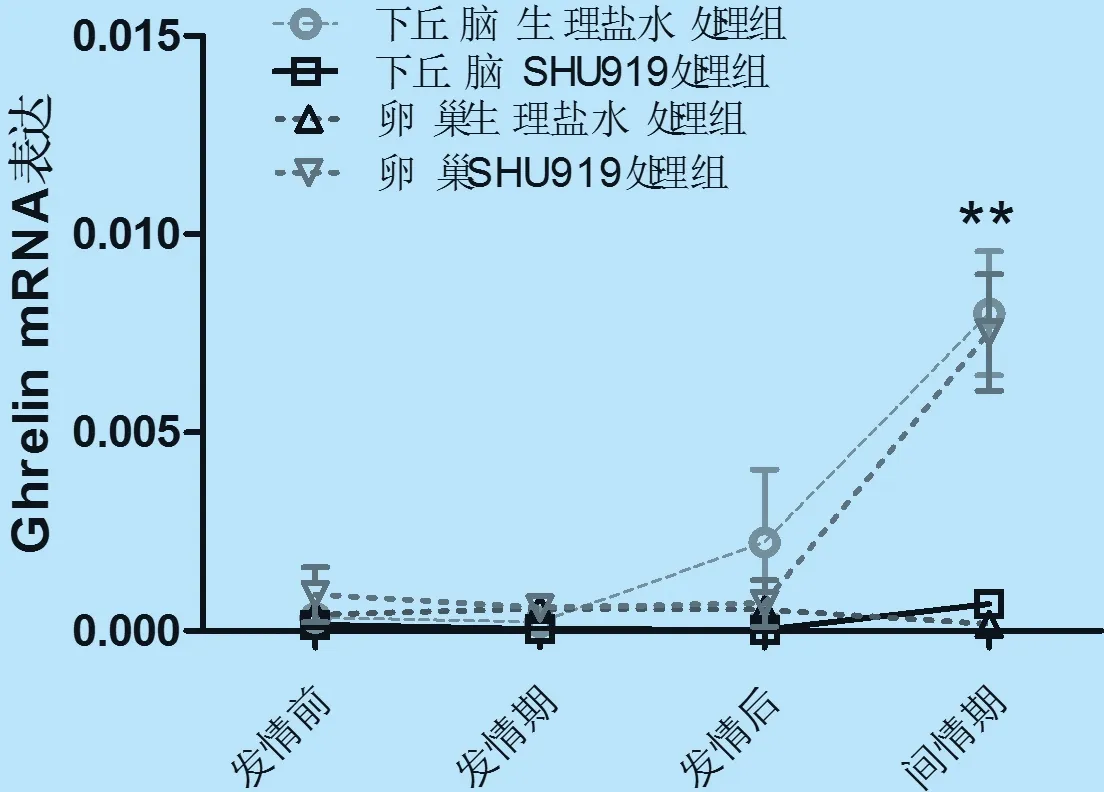
图3 SHU9119对小鼠发情周期内下丘脑和卵巢Ghrelin mRNA表达的影响
3 讨论
中枢神经系统特别是下丘脑,包含主要ghrelin效应位点,其中弓状核区域是重要的中枢整合核团[11]。在该区域,ghrelin通过传出神经纤维投射到神经肽Y/刺鼠相关肽(NPY/AgRP)神经元,促进与食欲相关的肽类物质释放[12]。本试验结果表明,SHU9119作为能量代谢关键调控基因MC4R的撷抗剂,可对小鼠间情期下丘脑和卵巢ghrelin mRNA进行逆向调控,而在发情前情和发情期未见负反馈调节。有报道表明,ghrelin促进体重增加的作用依雌二醇水平而动。ghrelin仅在大鼠间情期雌二醇水平较低时才可促进采食,在雄性大鼠中,外源雌二醇的注射可抑制Ghrelin所产生促进采食效应[13]。但,SHU9119在发情前情和发情期未见作用的结果是否与雌二醇水平有关尚需验证。
试验结果可知,正常生理条件下ghrelin在卵巢中表达未见显著差异(图3),但可hrelin mRNA表达水平在发情前期较低,发情期和发情后期升高,间情期降低的正态分布趋势,这种趋势与人、猪和绵羊等哺乳动物相类似[7, 14-15]。
小鼠下丘脑中,ghrelin mRNA表达却呈现逐渐升高(从发情前期到间情期)的过程。SHU9119作为一种能量调控抑制剂,在下丘脑-卵巢的性腺轴间存在着协调一致表达模式,支持雌性动物的生殖系统与能量平衡存在着紧密关系[16]。
是否可以这样理解,下丘脑中ghrelin mRNA在间情期维持较高水平即可为启动下一个发情期周期进行物质准备。正常生理条件下,ghrelin mRNA卵巢中表达量低较,下丘脑中较高,正是性腺轴的负反馈调节在控制的结果。另外,SHU9119作为能量代谢关键基因MC4R的撷抗剂致使间情期ghrelin mRNA表达水平在下丘脑和卵巢间出现相反的表达结果,推测表达能量代谢的相关基因在卵巢周期性变化过程中具有积极作用。
本试验结果表明,能量代谢相关基因MC4R受体抑制剂SHU9119可做为研究能量代谢与生殖间作用机理的潜在调控药物,同时SHU9119在雌性小鼠生殖方面可以起到类似正、负反馈调节的作用。生殖系统中激素对于能量代谢平衡影响积极,且生殖是一个复杂且一直围绕生物体能量代谢的生命活动过程,故从下丘脑-垂体-性腺轴的调控网络去理解ghrelin的功能,是一种研究ghrelin在动物体生殖和能量调控方面具有的潜在作用调控方向。
[1] Alvarez-Castro P, Pena L, Cordido F. Ghrelin in obesity, physiological and pharmacological considerations[J]. Mini Rev Med Chem, 2013,13(4):541-552.
[2] Kojima M, Hosoda H, Date Y,etal. Ghrelin is a growth-hormone-releasing acylated peptide from stomach[J]. Nature, 1999,402(6762):656-660.
[3] Sutton G M, Josephine B M, Gu X,etal. A derivative of the melanocortin receptor antagonist SHU9119 (PG932) increases food intake when administered peripherally[J]. Peptides, 2008,29(1):104-111.
[4] Shah B P, Vong L, Olson D P,etal. MC4R-expressing glutamatergic neurons in the paraventricular hypothalamus regulate feeding and are synaptically connected to the parabrachial nucleus[J]. Proceedings of the National Academy of Sciences, 2014,111(36):13 193-13 198.
[5] Huszar D, Lynch C A, Fairchild-Huntress V,etal. Targeted disruption of the melanocortin-4 receptor results in obesity in mice[J]. Cell, 1997,88(1):131-141.
[6] Rak-Mardyla A. Ghrelin role in hypothalamus-pituitary-ovarian axis[J]. J Physiol Pharmacol, 2013,64(6):695-704.
[7] Tena-Sempere M. Interaction between energy homeostasis and reproduction: central effects of leptin and ghrelin on the reproductive axis[J]. Horm Metab Res, 2013,45(13):919-927.
[8] Shaw A M, Irani B G, Moore M C,etal. Ghrelin-induced food intake and growth hormone secretion are altered in melanocortin 3 and 4 receptor knockout mice[J]. Peptides, 2005,26(10):1 720-1 727.
[9] Sutton G M, Josephine B M, Gu X,etal. A derivative of the melanocortin receptor antagonist SHU9119 (PG932) increases food intake when administered peripherally[J]. Peptides, 2008,29(1):104-111.
[10] Kojima M. Ghrelin: Structure and Function[J]. Physiological Reviews, 2005,85(2):495-522.
[11] Hassouna R, Labarthe A, Zizzari P,etal. Actions of Agonists and Antagonists of the ghrelin/GHS-R Pathway on GH Secretion, Appetite, and cFos Activity[J]. Frontiers in Endocrinology, 2013,4.
[12] Cowley M A, Smith R G, Diano S,etal. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis[J]. Neuron, 2003,37(4):649-661.
[13] Butera P C, Wojcik D M, Clough S J. Effects of estradiol on food intake and meal patterns for diets that differ in flavor and fat content[J]. Physiol Behav, 2010,99(1):142-145.
[14] Miller D W, Harrison J L, Brown Y A,etal. Immunohistochemical evidence for an endocrine/paracrine role for ghrelin in the reproductive tissues of sheep[J]. Reprod Biol Endocrinol, 2005,3:60.
[15] Garcia M C, Lopez M, Alvarez C V,etal. Role of ghrelin in reproduction[J]. Reproduction, 2007,133(3):531-540.
[16] Olofsson L E, Pierce A A, Xu A W. Functional requirement of AgRP and NPY neurons in ovarian cycle-dependent regulation of food intake[J]. Proc Natl Acad Sci U S A, 2009,106(37):15 932-15 937.
EffectofSHU9119onExpressionoftheGhrelininMiceduringtheEstrousCycle
WANG Cai-yun1, PAN Deng1, FAN Kui-kui1, ZHANG Zhao-yi1, LI Ting1, LI Qiang1, DU Chen-guang1,2
(1.College of Veterinary, Inner Mongolia Agricultural University,Hohhot 010018 , China; 2.Vocational and Technical College, Inner Mongolia Agricultural University, Baotou 014109 , China)
To confirm the biological role of ghrelin and SHU9119, 4 week old mice were used in our study to check the effects of SHU9119 on ghrelin mRNA expression using the hypothalamus and ovary at different estrous cycle. The results showed that under the normal physiological conditions, mice diestrus hypothalamus maintained higher level of ghrelin mRNA, while in the ovary remain the level was relatively low. SHU9119(50 μg / kg) significantly inhibited the hypothalamic ghrelin mRNA expression (P<0.01), but stimulated ovarian ghrelin mRNA expression in mouse diestrus(P<0.05). Meanwhile, there are no significant differences in mouse proestrus, estrus and metestrus. These results suggest that SHU9119 is as a regulator in mouse diestrus, and the effects of ghrelin are partly mediated by the hypothalamic melanocortinergic system.
Mouse ; Hypothalamus ; Ovary ; Ghrelin ; SHU9119
DU Chen-guang
S856.1+3
A
0529-6005(2017)09-0103-04
2017-04-17
国家自然科学基金(31660701,31160455);内蒙古高校自然科学重点项目(NJZZ049)
王彩云(1969-),女,副教授,博士,研究方向为家畜解剖学E-mail:wangcaiyun688@163.com
杜晨光,E-mail:dcglxj@163.com
