外源水杨酸、脯氨酸和γ-氨基丁酸对盐胁迫下水稻产量的影响
2017-10-17沙汉景胡文成王新鹏田雪飞于美芳赵宏伟
沙汉景 胡文成 贾 琰 王新鹏 田雪飞 于美芳 赵宏伟
东北农业大学水稻研究所, 黑龙江哈尔滨 150030
外源水杨酸、脯氨酸和γ-氨基丁酸对盐胁迫下水稻产量的影响
沙汉景 胡文成 贾 琰 王新鹏 田雪飞 于美芳 赵宏伟*
东北农业大学水稻研究所, 黑龙江哈尔滨 150030
水杨酸(SA)、脯氨酸(Pro)和γ-氨基丁酸(GABA)在植物生长发育以及抵御逆境胁迫中起重要作用。为了研究三者复配对盐胁迫下水稻产量是否具有协同增效作用, 本文采用三元二次通用旋转组合设计, 建立了 SA、Pro和GABA复配剂与盐胁迫下水稻产量之间的数学模型, 通过频数分析获得SA、Pro与GABA复配优化组合方案。结果表明, 二次模型拟合较好, 对盐胁迫下水稻产量的影响程度为SA>GABA>Pro, SA与GABA, Pro与GABA之间交互作用对盐胁迫下水稻产量有显著影响。模拟寻优获得提高盐胁迫下水稻产量10%以上的最佳复配组合为SA 0.44~0.51 mmol L–1、Pro 27.63~31.20 mmol L–1、GABA 3.55~4.28 mmol L–1。外源 SA、Pro 与 GABA 单剂和复配剂对盐敏感品种牡丹江30的调控作用大于耐盐品种龙稻5号。SA或Pro与GABA复配对盐胁迫下水稻产量的影响具有协同作用,而SA与Pro复配效果与品种耐盐性差异有关。SA、Pro和GABA三者复配效果优于两两复配, 任意单剂与其他2种外源物质复配对水稻产量的影响均存在协同作用。
二次通用旋转组合设计; 协同作用; 盐胁迫; 水稻; 产量
Abstract:Salicylic acid (SA), proline (Pro), and γ-aminobutyric acid (GABA) play important roles in plant growth and development and tolerating stress. In order to investigate whether the combination of SA, Pro, and GABA had synergistic effect on the yield of rice under salt stress, the mathematical models were established by means of quadratic general rotary unitized design. The optimal combination of SA, Pro, and GABA was obtained by frequency analysis. The results demonstrated that the quadratic models fit better. The effects on the yield of rice under salt stress were SA > GABA > Pro. The interactions between SA and GABA, and between Pro and GABA showed significant effects on rice yield under salt stress. The recommended optimal combination is 0.44-0.51 mmol L–1for SA, 27.63-31.20 mmol L–1for Pro and 3.55-4.28 mmol L–1for GABA, which can improve the rice yield more than 10% under salt stress. The effect of a single or a combination of SA, Pro, and GABA on the yield of salt-sensitive variety MDJ30 was larger than that of salt-tolerant variety LD5. The combination of SA and GABA, or Pro and GABA showed synergistic effect on the yield of rice under salt stress, while the combination effect of SA and Pro, was related to the genotype difference in salt tolerance. The joint application of SA, Pro, and GABA showed better effect on the yield of rice under salt stress than that of any two of them. The combination between any single agent and the other two agents showed synergistic effect.
Keywords:Quadratic general rotary unitized design; Synergistic effects; Salt stress; Rice; Yield
土壤盐渍化是世界范围内限制作物生长和产量 的主要逆境之一[1]。全球约有80亿公顷的陆地受盐渍化影响, 据估计其中 20%的灌溉土地(4.5亿公顷)受到不同程度的盐渍化影响[2]。中国受盐渍化影响的土地总面积为 3600万公顷, 占可利用土地的4.88%, 其中约920万公顷耕地受盐渍化影响, 占全国耕地面积的6.62%[3]。水稻(Oryza sativa L.)是世界上一种重要的粮食作物, 在谷类作物中对盐胁迫响应最为敏感[4]。我国盐/碱稻作区面积约占水稻栽培总面积的 20%[5]。盐胁迫已经成为限制我国水稻生产稳定发展的一个重要因素。水杨酸(salicylic acid,SA)是一种植物激素和信号分子, 在植物生长发育和抵御逆境胁迫中具有重要的功能, 影响植物水分代谢、矿质营养吸收和光合作用, 参与调节植物种子萌发、产热、开花、离子跨膜转运等生理过程[6-11]。有研究证实外源 SA促进盆栽控制盐胁迫条件下耐盐春小麦生长和产量[12], 以及大田盐渍条件下大麦产量[13]。逆境胁迫下, 植物体内大量积累脯氨酸(proline, Pro)和 γ-氨基丁酸(gamma-aminobutyric acid,GABA)等渗透调节物质。Pro和GABA在植物生长发育以及抵御逆境胁迫过程中起重要作用, 且二者在清除活性氧、维持渗透平衡、维持体内碳氮平衡以及作为信号分子等方面具有类似的功能。有研究证实外源 Pro提高了盆栽控制盐胁迫下水稻[14-15]和油菜[16]产量, 外源 GABA[17-18]提高作物幼苗耐盐性。目前, SA、Pro和GABA在提高作物耐盐性方面是否具有协同增效作用尚未见报道。本文在前人研究的基础上, 通过三元二次通用旋转组合设计试验,以盐胁迫下水稻产量为响应值, 模拟寻求三者复配优化浓度, 根据优化浓度研究 3种外源物质单剂和复配剂对盐胁迫下水稻产量的影响, 探讨三者复配是否具有协同增效作用, 以期为建立水稻耐盐栽培技术体系提供技术途径。
1 材料与方法
1.1 试验材料
采用本课题组前期筛选出的盐敏感品种牡丹江30 (MDJ30)和耐盐品种为龙稻5号(LD5)为材料。

表1 因子编码及实际值Table 1 Factor code and actual level
1.2 试验设计
2014年4 月至2016年10月在东北农业大学盆栽场进行盆栽试验。盆栽试验土壤 pH 5.8, 含有机质 31.25 g kg–1、全氮 1.73 g kg–1、碱解氮 138.1 mg kg–1、全磷 0.72 g kg–1、速效磷 46.8 mg kg–1、缓效钾 895.2 mg kg–1、速效钾 158.5 mg kg–1。盆规格: 高21.5 cm, 上底直径24.2 cm, 下底直径20.0 cm。土壤风干后过2 mm筛, 每盆10 kg土壤, 每盆施底肥尿素0.75 g, 磷酸二铵0.60 g, 硫酸钾0.56 g。采用大棚旱育秧, 4月20日播种, 秧龄35 d时移栽, 叶龄4.0~4.5叶。每盆4穴, 每穴3株。幼苗返青后进行盐胁迫处理, 为了防止水稻植株产生盐休克, 每隔1 d灌1 L 50 mmol L–1的NaCl溶液, 10 d后达到最终处理浓度, 此时土壤饱和浸提液电导率 ECe为(4.6±0.1) dS m–1(土壤中 NaCl量约为 0.15%)。此后为了模拟田间盐渍条件不再添加盐溶液, 为保证盐处理间浓度的一致性, 每盆插入相同标尺以控制水量, 移栽后每隔1 d补充水分至相应刻度。盐胁迫2周后喷施外源物质复配剂。2014—2015年, 采用SA(X1)、Pro (X2)和GABA (X3) 3种外源物质, 设每种物质5个水平(表1), 以中心组合设计复配, 共20个复配组合, 以喷施等量蒸馏水的常规处理(CK)和盐处理(Salt)为对照, 每个处理20盆。外源物质复配剂均含 0.1%的 Tween-20。选择晴朗无风的下午(15:00—17:00)连续喷施2 d, 正反面均喷, 以叶面完全湿润为宜(每盆约60 mL), 后期管理同大田生产。2016年选用零水平组合浓度, 分别进行单剂和复配剂验证试验, 各处理见表2, 其他处理方法同上。
1.3 测定项目及方法
完熟期取每处理9盆植株脱粒, 风干后测定产量。
1.4 统计分析
用Microsoft Excel 2007整理和计算数据, 采用Design-expert v8.05中的Quadratic回归分析模型分析响应面并作图, 参考徐中儒[19]介绍的方法优化分析回归方程, 采用因子贡献率法计算来判断各因子对产量作用的大小, 采用降维法分析单因子效应和二因子间交互作用, 采用频数分析法模拟寻优。

表2 试验处理及编码Table 2 Treatment and factor code
2 结果与分析
2.1 产量回归模型的建立及优化
由表 3可知, 盐胁迫下水稻产量显著下降。2014—2015年度, 盐胁迫下 MDJ30减产 34.0%和33.0%, LD5减产 18.3%和 17.9%; 盐胁迫下喷施外源复配剂的MDJ30产量范围分别为73.844~95.500 g pot–1和 70.756~90.859 g pot–1, 较 盐 处 理 增 加9.0%~48.2%; LD5的产量范围分别为80.067~97.244 g pot–1和 79.589~95.411 g pot–1, 较盐处理增加0.5%~24.8%。结果表明, 盐胁迫下喷施外源物质复配剂能够有效提高水稻产量, 对盐敏感品种的调控作用大于耐盐品种。
以 SA (X1)、Pro (X2)和 GABA (X3)为因变量, 产量(Y)为目标变量建立模型, 得到 MDJ30产量的回归模型 YMDJ30(2014)= 90.765 – 2.197 X1+ 0.468 X2–0.418 X3– 0.555 X1X2– 5.075 X1X3+ 2.617 X2X3–2.467 X12– 1.417 X22– 1.678 X32; YMDJ30(2015)= 88.147– 1.758 X1– 0.113 X2+ 0.322 X3+ 0.510 X1X2– 4.782 X1X3+ 3.102 X2X3– 4.463 X12– 2.697 X22– 2.261 X32。LD5 的回归模型为 YLD5(2014)= 94.797 – 1.019 X1–0.575 X2+ 0.269 X3+ 1.279 X1X2– 4.804 X1X3+ 1.813 X2X3– 3.244 X12– 1.413 X22– 1.845 X32; YLD5(2015)=91.503 – 1.025 X1– 0.460 X2+0.131 X3+ 0.564 X1X2–3.519 X1X3+ 1.697 X2X3– 3.082 X12– 1.666 X22–1.580 X32。
需要进一步检验拟合得到的方程, 以提高方程的稳定性。回归方程的检验结果列于表 4和表 5。首先, 对MDJ30和LD5的产量回归方程进行失拟性检验, F1-MDJ30(2014)= 0.277 (P>0.05), F1-MDJ30(2015)=0.564 (P>0.05), F1-LD5(2014)= 0.294 (P>0.05),F1-LD5(2015)= 0.317 (P>0.05), 说明可以忽略模型的失拟部分。其次, 对MDJ30和LD5产量拟合方程的总模型进行 F测验, F2-MDJ30(2014)= 12.653 (P<0.01),F2-MDJ30(2015)= 10.131 (P<0.01), F2-LD5(2014)= 10.116(P<0.01), F2-LD5(2015)= 7.709 (P<0.01), 说明模型在0.01水平上极显著, 表明试验设计可靠, 预测值与实际值吻合性好。通过复决定系数R2和校正复决定系数R2adj可以判断回归模型的拟合精度, MDJ30回归模型的复决定系数分别为0.919和0.901, LD5回归模型的复决定系数分别为0.901和0.874, 说明三因素对盐胁迫下 MDJ30产量影响达 90.1%~91.9%,其余8.1%~9.9%是由其他未控制因素和误差造成的;三因素对盐胁迫下 LD5产量影响达 87.4%~90.1%,其余 9.9%~12.6%是由其他未控制因素和误差造成的。2年试验结果表明, 利用响应面能够有效建立稳定的试验模型。
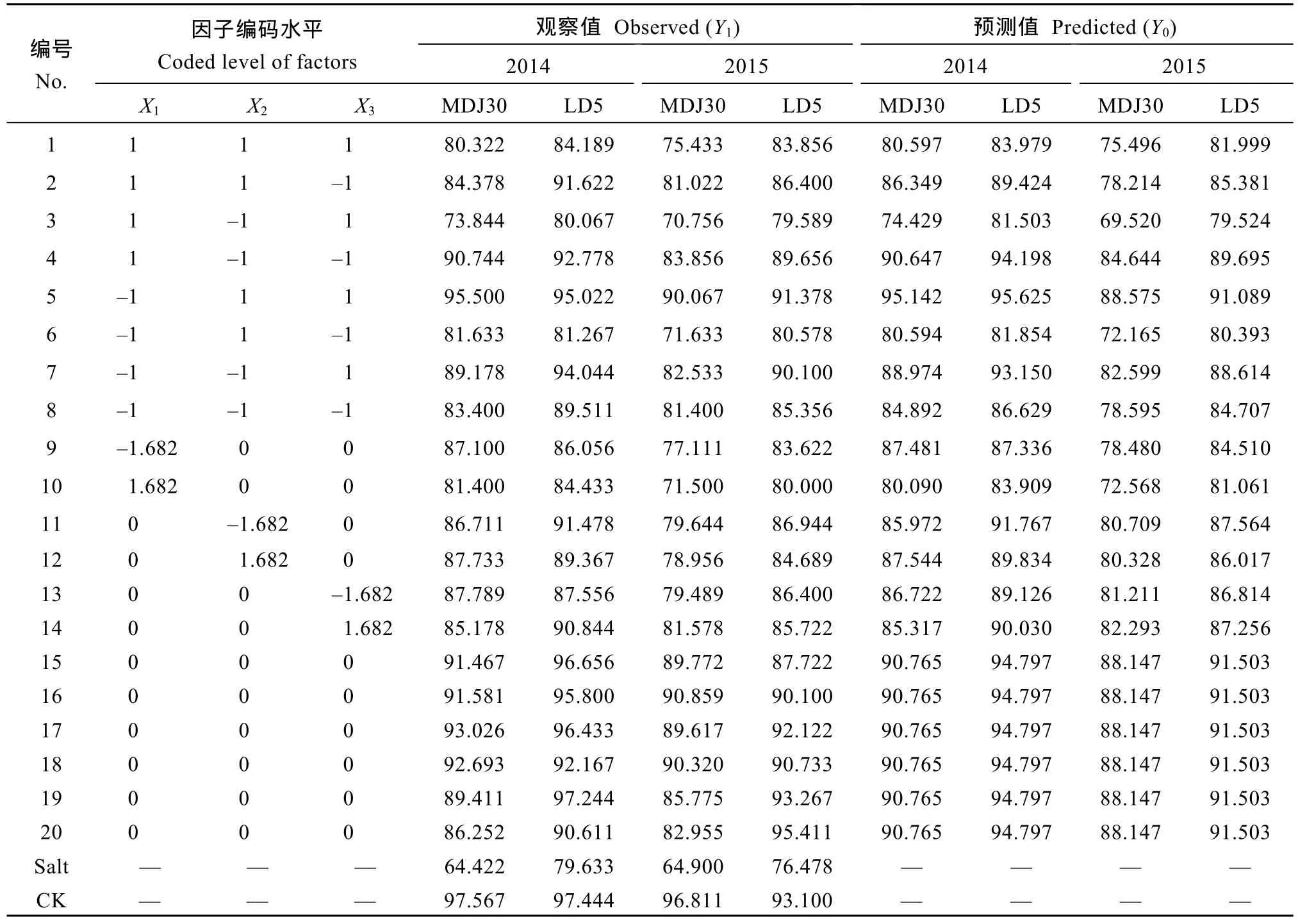
表3 外源水杨酸、脯氨酸和γ-氨基丁酸复配组合对盐胁迫下水稻产量的影响Table 3 Combination effect of SA, Pro, and GABA on the yield of rice under salt stress (g pot–1)
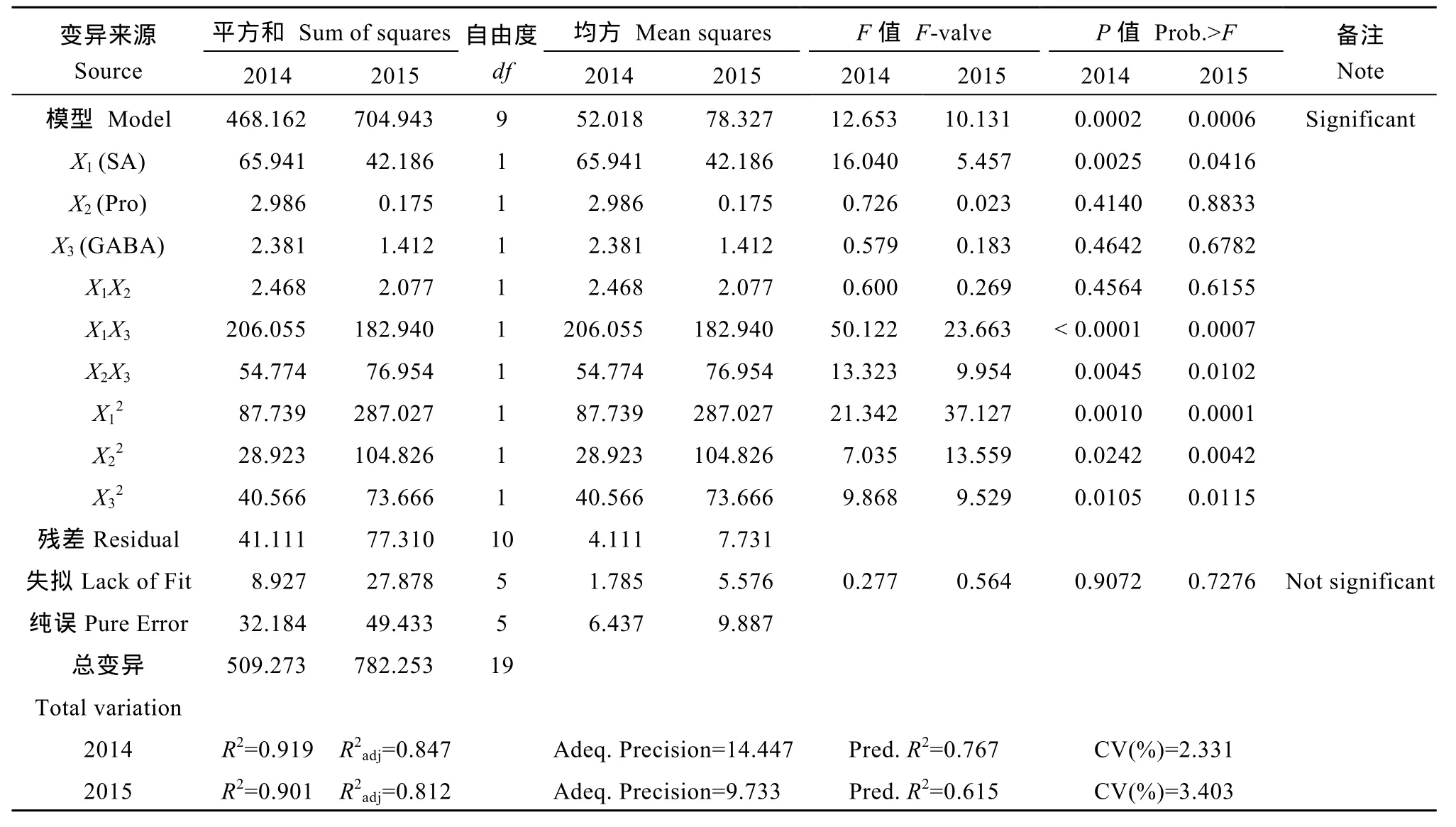
表4 牡丹江30产量响应面二次模型的方差分析表Table 4 ANOVA for response surface quadratic model to the yield of MDJ30

表5 龙稻5号产量响应面二次模型的方差分析表Table 5 ANOVA of response surface quadratic model of LD5 yield
对回归系数进行显著性检验, 在σ = 0.10水平下采用逐步回归剔除不显著项, 得优化方程YMDJ30(2014)= 90.765 – 2.197 X1+ 0.468 X2–0.418 X3–5.075 X1X3+ 2.617 X2X3– 2.467 X12– 1.417 X22–1.678 X32; YMDJ30(2015)= 88.147 – 1.758 X1– 0.113 X2+0.322 X3– 4.782 X1X3+ 3.102 X2X3– 4.463 X12– 2.697 X22– 2.261 X32。LD5 的回归模型为 YLD5(2014)= 94.797– 1.019 X1– 0.575 X2+ 0.269 X3– 4.804 X1X3+ 1.813 X2X3– 3.244 X12– 1.413 X22– 1.845 X32; YLD5(2015)=91.503 – 1.025 X1– 0.460 X2+0.131 X3– 3.519 X1X3+1.697 X2X3– 3.082 X12– 1.666 X22– 1.580 X32。
2.2 因子重要性分析
采用因子贡献率法[19]判断各因子对产量作用的大小, 经过计算得到SA、Pro和GABA对盐胁迫下MDJ30产量的贡献率, 2014年分别为2.381、1.320和1.851, 2015年分别为2.269、1.376和1.824; 三因子对盐胁迫下 LD5产量的贡献率, 2014年分别为2.427、1.553和 1.795, 2015年分别为 2.115、1.281和1.744。上述结果表明对盐胁迫下水稻产量的影响为 SA>GABA>Pro。
2.3 单因子效应分析
采用降维法分析单因子效应, 即将目标因子之外的二因子编码值固定在零水平上, 可得到目标因子与盐胁迫下水稻产量的一元回归模型, 并根据模型作图(图1)。由图1可知, 三因子对盐胁迫下水稻产量的影响均呈开口向下的抛物线状。盐胁迫下水稻产量随喷施SA浓度的升高先缓慢增加, 当SA浓度超过一定值后, 水稻产量迅速下降, 二者呈不对称抛物线变化; Pro和GABA对盐胁迫下水稻产量的影响相似, 产量均随喷施浓度的增加呈先缓慢升高,接近零水平时达到最高, 而后逐渐降低, 表现为对称的抛物线变化。
2.4 双因子互作效应分析
SA与 GABA互作对盐胁迫下水稻产量存在极显著影响(P<0.01), Pro与GABA互作对产量的影响显著(除2015年LD5产量外, P<0.05), 而SA与Pro互作对水稻产量影响不显著(P>0.05)。由图 2可知,2014—2015年间, SA与GABA的交互作用对盐胁迫下不同水稻品种产量的影响基本一致。当 Pro固定在零水平时, 随SA、GABA浓度的增加, 产量呈先增加后下降的趋势。SA在 GABA低水平下比高水平下增产效应大, 说明在 GABA浓度较低时, 增施SA有利于增产。当SA在高浓度水平时, 增施GABA造成产量逐渐下降; 当SA在低浓度水平时, 产量随GABA的增加而增加。说明在SA浓度低时, 应提高GABA的用量从而获得较高的产量。由图3可知, Pro与GABA互作效应对盐胁迫下2个水稻品种产量的影响在2014—2015年间大致相同。当SA固定在零水平时, 随Pro、GABA浓度的增加, 产量呈先增加后下降的趋势。Pro在GABA高水平下比低水平下增产效应大, 说明GABA在高浓度水平时, 增施Pro有利于增产。Pro在高浓度水平下, 增施GABA造成产量先快速增加后缓慢降低; Pro在低浓度水平下,产量随GABA的增加呈先缓慢增加后迅速下降, 说明Pro在高浓度水平时, 应控制GABA的过多施用。
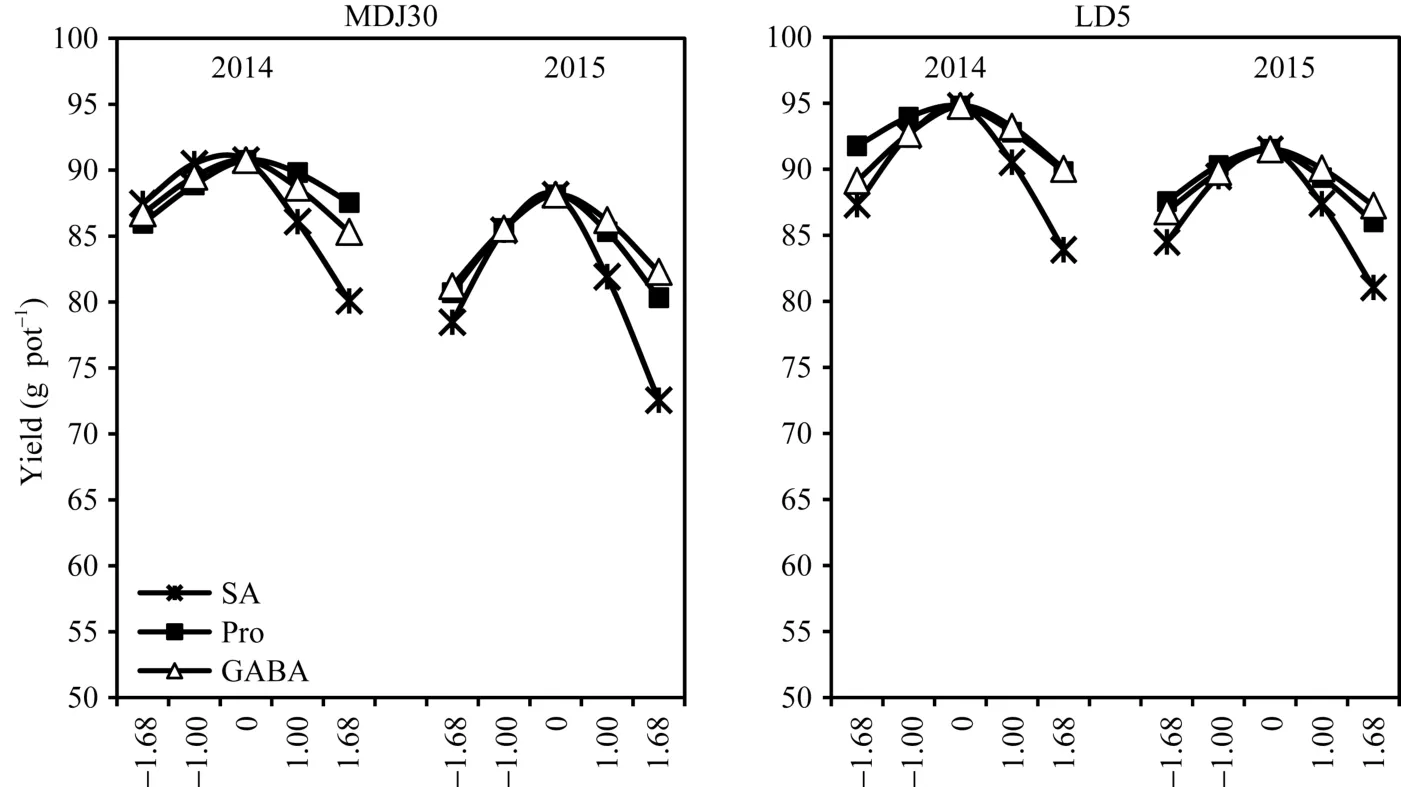
图1 任意二因子固定在零水平时的单因子效应分析Fig. 1 Single factor analysis with other two factors fixed at zero levels
2.5 复配组合优化方案及验证
根据试验所建立的数学模型, 利用计算机模拟和筛选优化组合方案。以提高盐胁迫下水稻产量10%以上为目标, 以 95%置信区间为取值范围, 选取两年间各指标取值的交叉值, 以进一步优化复配组合方案, 从而建立外源物质复配组合优化技术方案。试验的技术参数及各因素的优化组合频数分析见表6和表7。
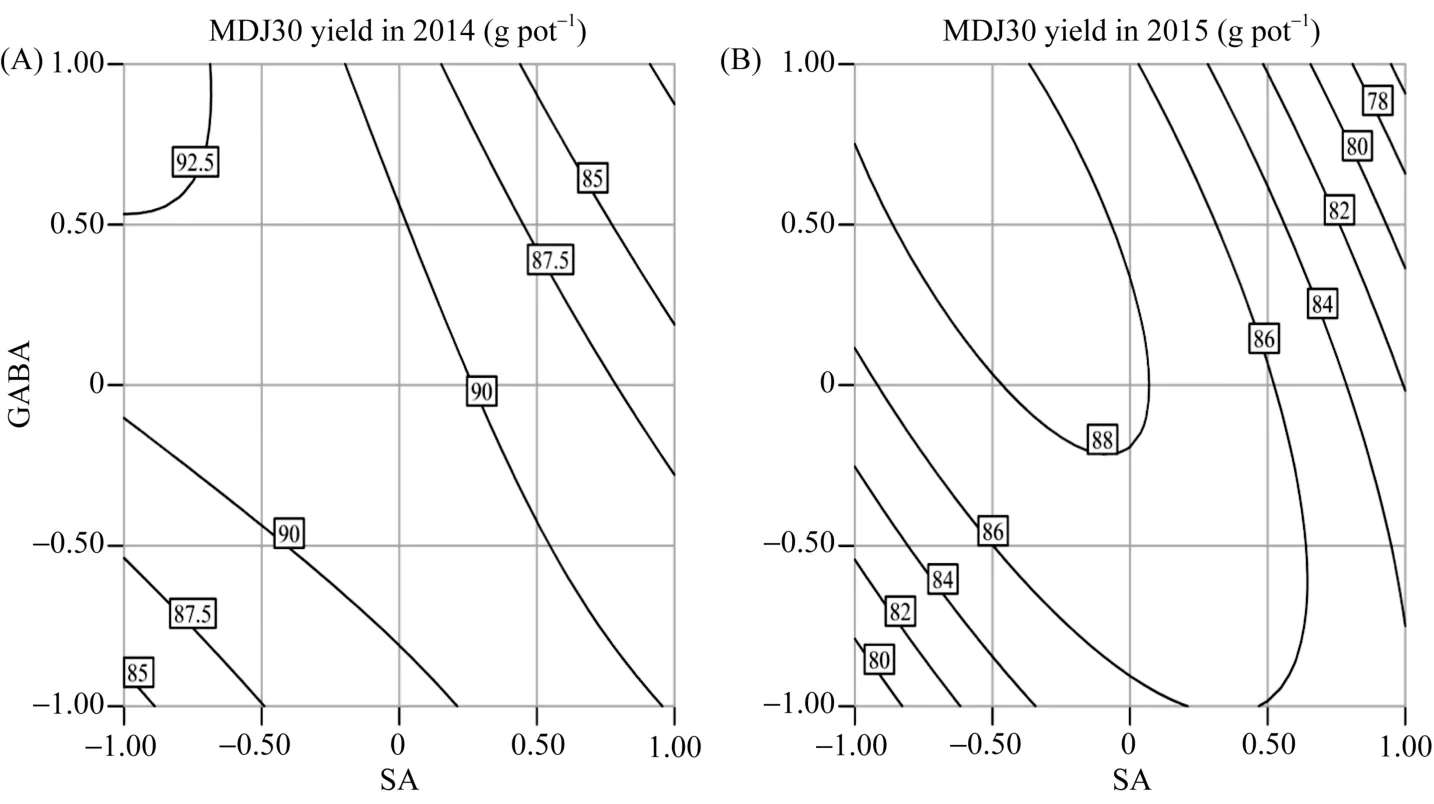
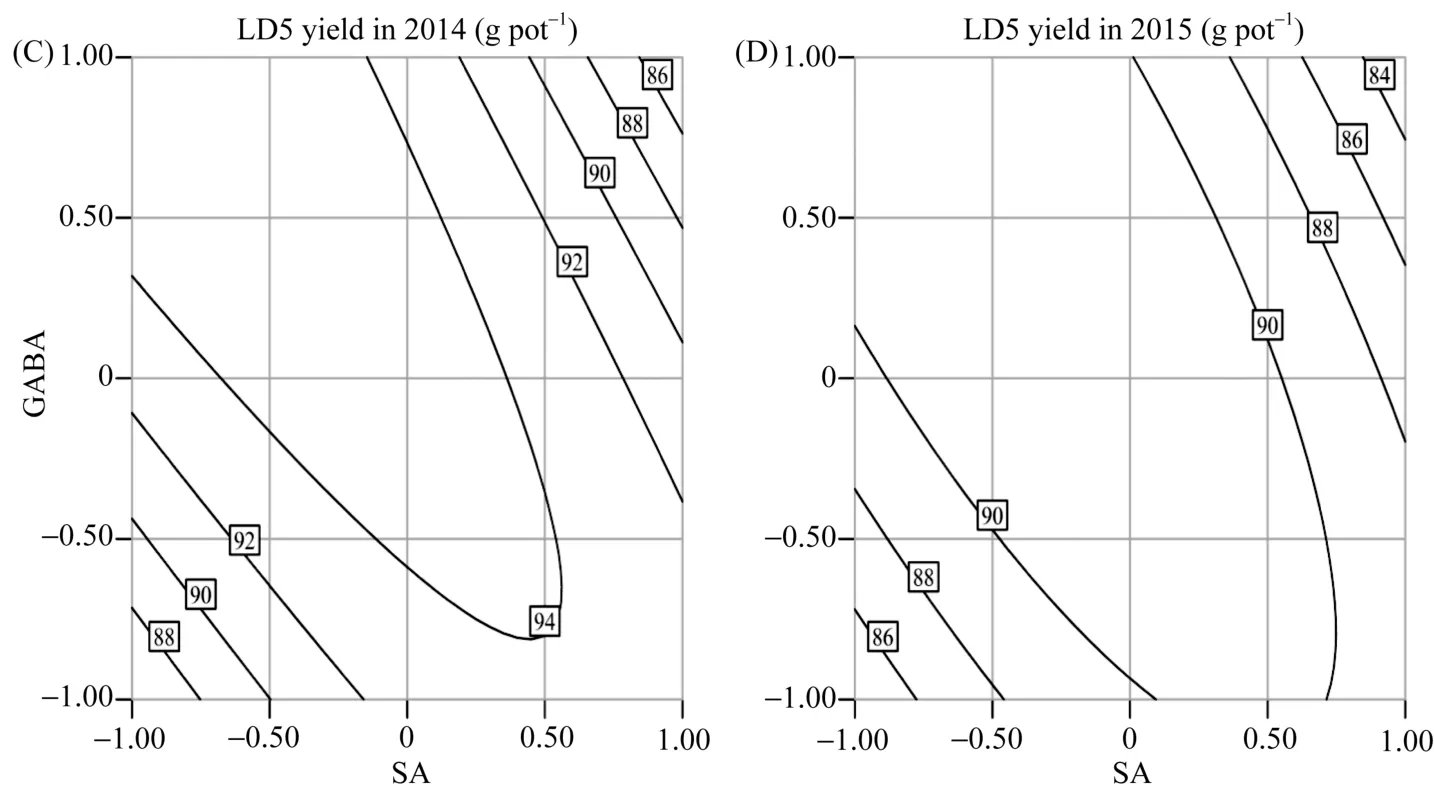
图2 水杨酸与γ-氨基丁酸交互作用对盐胁迫下水稻产量的影响Fig. 2 Interaction effect of SA and GABA on the yield of salt-stressed rice
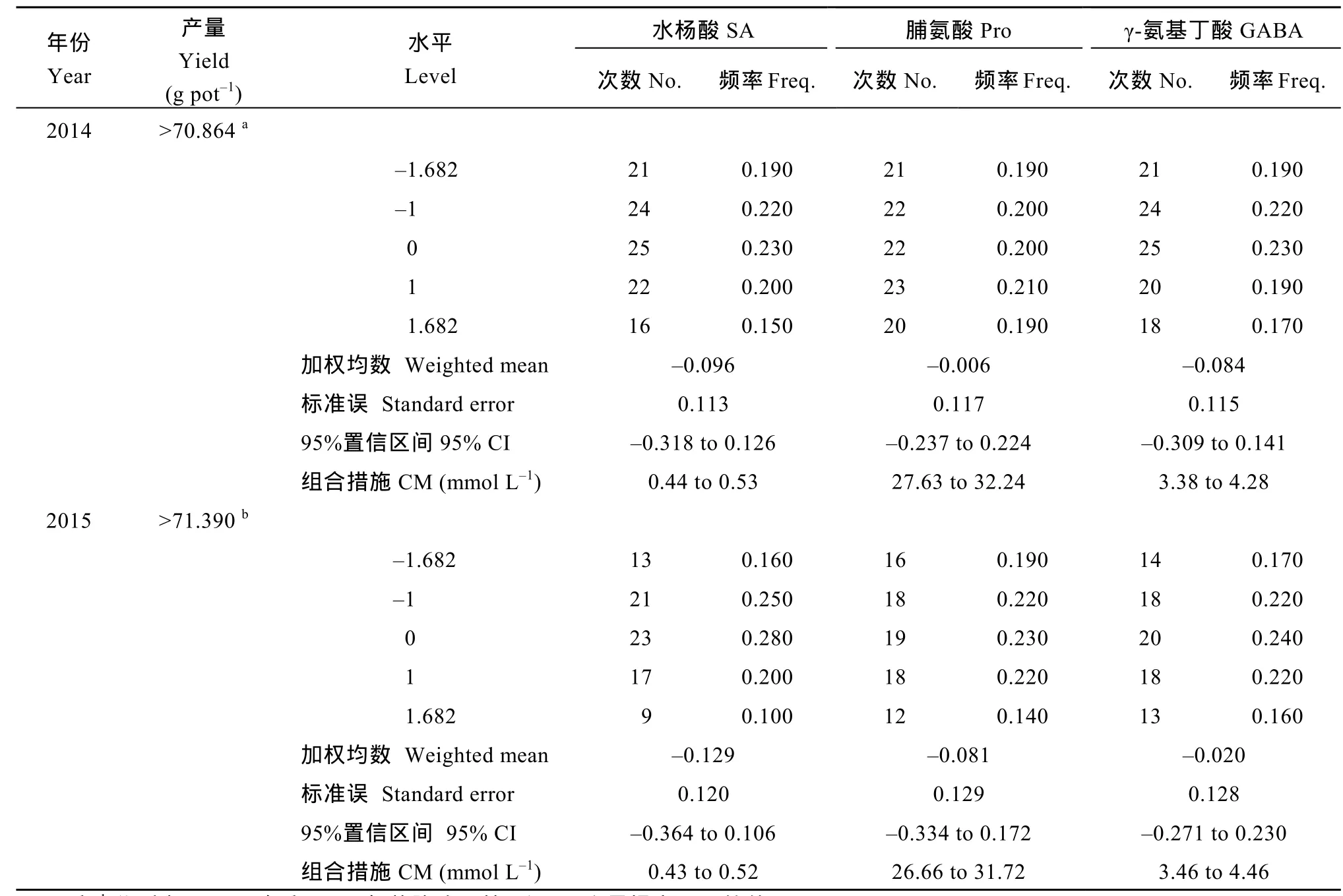
表6 盐胁迫下牡丹江30产量提高10%以上的水杨酸、脯氨酸和γ-氨基丁酸复配组合Table 6 Combination of SA, Pro, and GABA improving yield of MDJ30 more than 10% under salt stress
2014—2015 年, 盐胁迫下MDJ30产量提高10%以上的外源物质复配组合方案分别有108个和83个,综合2年分析结果, 推荐MDJ30的外源物质复配方案为 SA 0.44~0.52 mmol L–1, Pro 27.63~31.72 mmol L–1, GABA 3.46~4.28 mmol L–1。2014—2015 年, 盐胁迫下LD5产量提高10%以上的外源物质复配组合方案分别有66个和60个, 综合2年分析结果, 推荐LD5的外源物质复配方案为SA 0.41~0.51 mmol L–1,Pro 26.32~31.20 mmol L–1, GABA 3.55~4.69 mmol L–1。由此可以看出, 2个不同耐盐性水稻品种适用的外源物质复配组合浓度较为接近, 因此, 可以选取交叉值作为 2个品种共同适用的优化组合方案, 各指标取值为 SA 0.44~0.51 mmol L–1, Pro 27.63~31.20 mmol L–1, GABA 3.55~4.28 mmol L–1。由于推荐的组合方案包含本试验设计中的零点, 因此, 本试验选取各指标零点, 即SA 0.50 mmol L–1、Pro 30.0 mmol L–1、GABA 4.0 mmol L–1进行验证。

图3 脯氨酸与γ-氨基丁酸交互作用对盐胁迫下水稻产量的影响Fig. 3 Interaction effect of Pro and GABA on the yield of salt-stressed rice
2016年3 种外源物质单剂和复配剂对产量的影响列于表8。盐胁迫下MDJ30和LD5分别减产31.8%和 19.4%。盐胁迫下喷施 3种外源复配剂的单剂和复配剂均能显著提高水稻产量。与盐处理相比, 外源SA、Pro和GABA单剂处理下MDJ30产量分别增加18.4%、4.9%和10.8%, LD5产量分别增加8.6%、4.8%和 6.8%。与盐处理相比, 外源 SP、SG和 PG处理下MDJ30产量分别增加20.3%、29.9%和13.8%,LD5产量分别增加13.9%、16.5%和11.6%; 与单剂相比, SG或PG复配对盐胁迫下水稻产量的影响具有协同作用, 而SP复配效果与品种耐盐性有关, 即对MDJ30产量无协同作用, 对LD5具有协同作用。外源SA、Pro和GABA三者复配处理下MDJ30产量与 2014—2015年预测产量之间的相对误差为1.9%~4.7%, 较盐胁迫下产量提高34.9%, LD5产量与 2014—2015年预测产量之间的相对误差为3.3%~6.7%, 较盐胁迫下产量提高21.3%。从上述结果可以看出盐胁迫下外源SA、GABA和Pro单剂及复配剂对水稻产量的影响为 SPG>SG>SP>PG>SA>GABA>Pro, 3种外源物质对盐敏感品种MDJ30产量的调控作用大于耐盐品种 LD5, SA、Pro和 GABA任意单剂处理与其他 2种外源物质复配对水稻产量的影响均存在协同作用。综上所述, 3种外源物质复配较单剂或两两复配效果好, 且可以实现提高盐胁迫下水稻产量10%的预期目标。

表7 盐胁迫下龙稻5号产量提高10%以上的水杨酸、脯氨酸和γ-氨基丁酸复配组合Table 7 Combination of SA, Pro, and GABA improving yield of LD5 more than 10% under salt stress
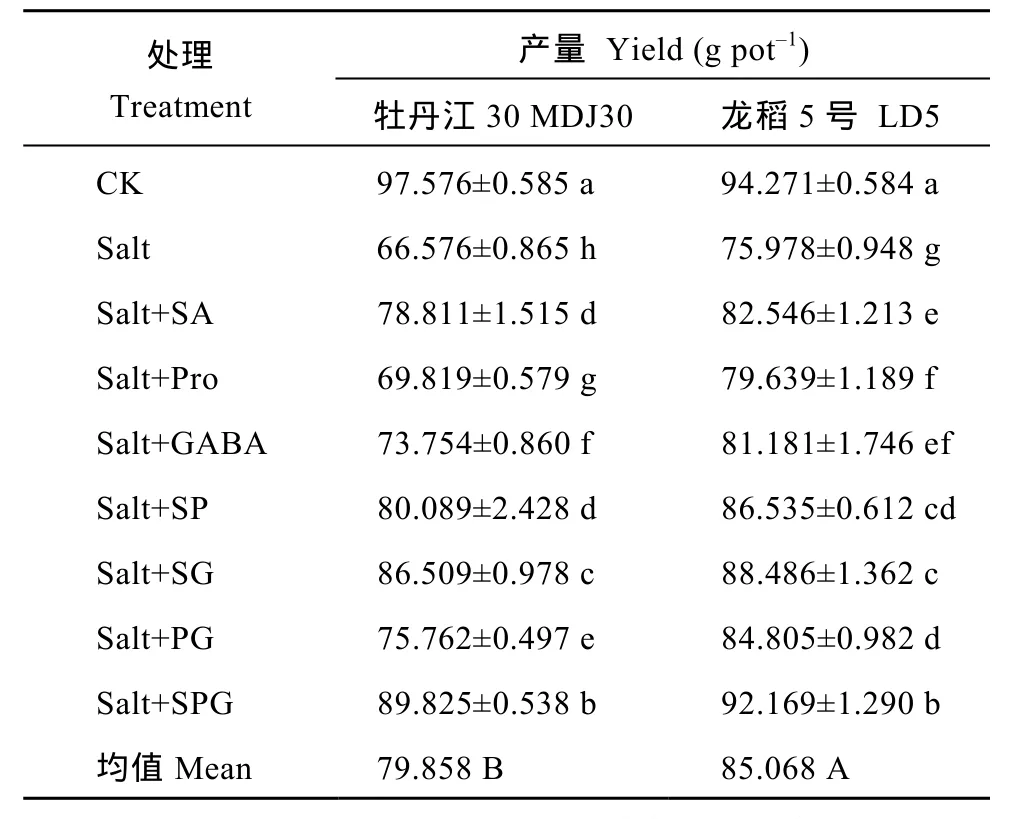
表8 外源水杨酸、脯氨酸和γ-氨基丁酸单剂及复配剂对盐胁迫下水稻产量的影响Table 8 Effect of exogenous SA, Pro, and GABA on the yield of rice under salt stress
3 讨论
水稻是对盐最敏感的谷类作物。基于室内研究发现水稻盐度阈值是 3.0 dS m–1, 超过阈值每增加1.0 dS m–1产量减少12.0%[20]。而另一项田间试验指出水稻的盐度阈值是 1.9 dS m–1, 超过阈值每增加1.0 dS m–1产量减少9.1%[21], 表明田间条件下水稻对盐渍环境更为敏感。本试验盐处理下的土壤电导率为 4.6 dS m–1, 按上述室内或田间鉴定阈值计算,水稻产量下降幅度理论上应为19.2%或24.6%, 而本试验中盐胁迫下耐盐品种 LD5在 2014—2016年实测产量分别下降 18.3%、17.9%和 19.4%, 盐敏感品种MDJ30产量分别下降34.0%、33.0%和31.8%, 说明耐盐品种 LD5盐度阈值接近 Mass等室内鉴定值[20],而盐敏感品种MDJ30盐度阈值低于Grattan等的田间鉴定值[21]。
近年来, 关于单独施用水杨酸[12-13,22-24]、脯氨酸[14-16,25-27]和 γ-氨基丁酸[17-18,28-29]能够提高植物耐盐性的报道比较多, 其施用方式主要有 3种, 即种子预处理[22,25,28]、生根基质[12,17-18,26-27]和叶面
喷施[13-16,23-24,29]。多数研究结果表明3种外源物质具有浓度效应, 并且其有效性与植物种类、发育阶段、胁迫强度和时间有关。大多数研究在作物的萌发期和幼苗期施用外源物质, 改善盐胁迫下作物种子萌发和幼苗生长。由于作物生长发育是一个连续过程, 前期施用外源物质的有效性能否延续至作物生育后期而影响作物产量, 目前对此报道尚少。
Arfan 等[12]研究发现, 根施 0.25 mmol L–1SA 可改善光合作用, 促进耐盐春小麦品种生长, 增加百粒重、穗粒数和每穗颖花数并提高产量, 但对中度盐敏感品种无明显作用。Pirasteh-Anosheh等[13]连续3年在大田条件下证实, 拔节期喷施 0.5~2.0 mmol L–1SA可以显著提高盐胁迫下大麦产量。我们前期研究证实分蘖期和孕穗期叶面喷施30 mmol L–1Pro可以有效提高水稻产量[14]。Bhusan等[15]研究发现,分蘖盛期叶面喷施25 mmol L–1和50 mmol L–1Pro可以有效提高25 mmol L–1NaCl胁迫下盐敏感水稻品种产量, 但对耐盐品种产量无显著作用, 且不能缓解 50~100 mmol L–1NaCl胁迫。El Habbasha等[16]报道称叶面喷施0.1 mmol L–1Pro可以提高油菜单株荚数、产量、秸秆产量和生物产量。外源 GABA能够提高盐胁迫下玉米种子萌发率及玉米幼苗的生长[17], 缓解 NaCl胁迫对黄瓜幼苗生长的抑制作用,显著提高叶片SOD、POD和CAT活性[18], 保护甜瓜叶绿体结构以及PSII功能[29], 但目前关于GABA对盐胁迫下作物产量影响的研究尚未见报道。本研究证实SA、Pro和GABA单剂和复配剂能够有效提高盐胁迫下水稻产量, 且 SA的作用最大, 其次为GABA和Pro。有研究称利用外源SA[12,30]或Pro[26,31]处理对耐盐品种的调控作用大于盐敏感品种, 但也有报道称外源Pro[15]或GABA[32]对盐/盐碱敏感品种的调控作用大于耐盐/盐碱品种。本研究中发现外源SA、Pro和GABA对盐敏感品种的调控作用大于耐盐品种, 这可能与品种特异性有关。此外, 本研究还发现SG或PG复配具有协同作用, 而SP复配效果与品种耐盐性有关, SA、Pro和GABA单剂处理与任意两种外源物质复配对水稻产量的影响均存在协同作用, 但其互作的内在机制尚不清楚。前人研究表明, 盐胁迫条件下水杨酸对植物体内的氮素同化过程具有促进作用[30,33-34]。而盐胁迫下脯氨酸和 γ-氨基丁酸的大量积累依赖于其从头合成过程, 且二者从头合成底物均为氮素同化的产物——谷氨酸。已有研究证实外源SA能够促进包括Pro在内的多种氨基酸积累[35], 通过 GAD磷酸化或去磷酸化影响基础代谢[36]。外源GABA能够促进内源GABA和Pro等氨基酸的积累[36]。因此我们推测SA、Pro与GABA之间的互作过程可能与氮素同化的代谢平衡有关,这有待进一步研究。
此外, 本试验于移栽返青后的分蘖始期开始盐胁迫处理, 所建立的模型以及由此决策获得的最优组合均是在盆栽控制条件下实现的, 因此对于大田生产应用还有待进一步验证。
4 结论
推荐了本试验条件下提高盐胁迫下水稻产量10%以上的外源物质复配方案, SA 为 0.44~0.51 mmol L–1, Pro 为 27.63~31.20 mmol L–1, GABA 为3.55~4.28 mmol L–1。外源 SA、Pro和 GABA 单剂和复配剂能够提高盐胁迫下水稻产量, 对盐敏感品种MDJ30的调控作用大于耐盐品种 LD5。SA或 Pro与GABA复配对盐胁迫下水稻产量的影响具有协同作用, 而 SA与 Pro复配效果与品种耐盐性差异有关。SA、Pro和GABA三者复配效果优于两两复配,任意单剂处理与其他 2种外源物质复配对水稻产量的影响均存在协同作用。
[1]Koca H, Bor M, Özdemir F, Türkan İ. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ Exp Bot, 2007, 60: 344–351
[2]Negrão S, Courtois B, Ahmadi N, Abreu I, Saibo N, Oliveira M M. Recent updates on salinity stress in rice: from physiological to molecular responses. Crit Rev Plant Sci, 2011, 30: 329–377
[3]杨劲松. 中国盐渍土研究的发展历程与展望. 土壤学报, 2008,45: 837–845 Yang J S. Development and prospect of the research on-salt-affected soils in china. Acta Pedol Sin, 2008, 45: 837–845 (in Chinese with English abstract)
[4]Walia H, Wilson C, Zeng L, Ismail A M, Condamine P, Close T J.Genome-wide transcriptional analysis of salinity stressed japonica and indica rice genotypes during panicle initiation stage.Plant Mol Biol, 2007, 63: 609–623
[5]祁栋灵, 韩龙植, 张三元. 水稻耐盐/碱性鉴定评价方法. 植物遗传资源学报, 2005, 6: 226–230 Qi D L, Han L Z, Zhang S Y. Methods of characterization and evaluation of salt or alkaline tolerance in rice. J Plant Genet Resour, 2005, 6: 226–230 (in Chinese with English abstract)
[6]Jayakannan M, Bose J, Babourina O, Rengel Z, Shabala S. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul, 2015, 76: 25–40
[7]Janda M, Ruelland E. Magical mystery tour: salicylic acid signalling. Environ Exp Bot, 2015, 114: 117–128
[8]Kang G, Li G, Guo T. Molecular mechanism of salicylic acid-induced abiotic stress tolerance in higher plants. Acta Physiol Plant, 2014, 36: 2287–2297
[9]Horváth E, Szalai G, Janda T. Induction of abiotic stress tolerance by salicylic acid signaling. J Plant Growth Regul, 2007, 26:290–300
[10]Ashraf M, Akram N A, Arteca R N, Foolad M R. The physiological, biochemical and molecular roles of brassinosteroids and salicylic acid in plant processes and salt tolerance. Crit Rev Plant Sci, 2010, 29: 162–190
[11]Hayat Q, Hayat S, Irfan M, Ahmad A. Effect of exogenous salicylic acid under changing environment: a review. Environ Exp Bot, 2010, 68: 14–25
[12]Arfan M, Athar H R, Ashraf M. Does exogenous application of salicylic acid through the rooting medium modulate growth and photosynthetic capacity in two differently adapted spring wheat cultivars under salt stress? J Plant Physiol, 2007, 164: 685–694
[13]Pirasteh-Anosheh H, Emam Y, Rousta M J, Ashraf M. Salicylic acid induced salinity tolerance through manipulation of ion distribution rather than ion accumulation. J Plant Growth Regul,2017, 36: 227–239
[14]沙汉景. 外源脯氨酸对盐胁迫下水稻耐盐性的影响. 东北农业大学硕士学位论文, 黑龙江哈尔滨, 2013 Sha H J. Effect of Exogenous Proline on the Salt-tolerance of Rice (Oryza sativa L.). MS Thesis of Northeast Agricultural University, Harbin, China, 2013 (in Chinese with English abstract)
[15]Bhusan D, Das D K, Hossain M, Murata Y, Hoque M A. Improvement of salt tolerance in rice (Oryza sativa L.) by increasing antioxidant defense systems using exogenous application of proline. Aust J Crop Sci, 2016, 10: 50–56
[16]El Habbasha S F, Mekki B B. Amelioration of the growth, yield and chemical constituents of canola plants grown under salinity stress condition by exogenous application of proline. Int J Adv Res, 2013, 2: 501–508
[17]周翔. 盐胁迫诱导玉米幼苗 GABA 积累的生理作用. 中国农业大学硕士学位论文, 北京, 2004 Zhou X. The Physiological Role of salt Stress-induced GABA Accumulation in Zea Mays. MS Thesis of China Agricultural University, Beijing, China, 2004 (in Chinese with English abstract)
[18]高洪波, 章铁军, 吕桂云, 吴晓蕾, 周志男. NaCl胁迫下外源γ-氨基丁酸对黄瓜幼苗生长和活性氧代谢的影响. 西北植物学报, 2007, 27: 2046–2051 Gao H B, Zhang T J, Lyu G Y, Wu X L, Zhou Z N. Effects of exogenous γ-aminobutyric acid on growth and reactive oxygen species metabolism of cucumber seedlings under NaCl stress. Acta Bot Boreali-Occident Sin, 2007, 27: 2046–2051 (in Chinese with English abstract)
[19]徐中儒. 回归分析与试验设计. 北京: 中国农业出版社, 1998.pp 124–161 Xu Z R. Regression Analysis and Experimental Design. Beijing:China Agriculture Press, 1998. pp 124–161 (in Chinese)
[20]Maas E V, Grattan S R. Crop yields as affected by salinity. In: van Schilfgaarde J, Skaggs R W, eds. Agricultural Drainage. Madison,WI: ASA, CSSA, SSA, 1999. pp 55–108
[21]Grattan S R, Zeng L, Shannon M C, Roberts S R. Rice is more sensitive to salinity than previously thought. Calif Agric, 2002,56(6): 189–198
[22]Esan A M, Olaiya C O. Effect of salicylic acid (SA) seeds soaking on the NaCl salt stress induced changes in soluble sugar and protein accumulation in organs of two genotypes of okra plants.Afr J Plant Sci, 2016, 10(6): 105–110
[23]Farhangi-Abriz S, Ghassemi-Golezani K. Improving amino acid composition of soybean under salt stress by salicylic acid and jasmonic acid. J Appl Bot Food Qual, 2016, 89: 243–248
[24]Qados A M S A. Effects of salicylic acid on growth, yield and chemical contents of pepper (Capsicum annuum L.) plants grown under salt stress conditions. Int J Agric Crop Sci, 2015, 8: 107–113
[25]Agami R A. Applications of ascorbic acid or proline increase resistance to salt stress in barley seedlings. Biol Plant, 2014, 58:341–347
[26]Teh C Y, Shaharuddin N A, Ho C L, Mahmood M. Exogenous proline significantly affects the plant growth and nitrogen assimilation enzymes activities in rice (Oryza sativa) under salt stress. Acta Physiol Plant, 2016, 38(6): 1–10
[27]Wutipraditkul N, Wongwean P, Buaboocha T. Alleviation of salt-induced oxidative stress in rice seedlings by proline and/or glycinebetaine. Biol Plant, 2015, 59: 547–553
[28]Ziogas V, Tanou G, Belghazi M, Diamantidis G, Molassiotis A.Characterization of β-amino- and γ-amino butyric acid-induced citrus seeds germination under salinity using nanoLC-MS/MS analysis. Plant Cell Rep, 2017, 36: 787–789
[29]Xiang L, Hu L, Xu W, Zhen A, Zhang L, Hu X. Exogenous γ-aminobutyric acid improves the structure and function of photosystem II in muskmelon seedlings exposed to salinity-alkalinity stress. PLoS One, 2016, 11(10): e0164847
[30]Nazar R, Iqbal N, Syeed S, Khan N A. Salicylic acid alleviates decreases in photosynthesis under salt stress by enhancing nitrogen and sulfur assimilation and antioxidant metabolism differentially in two mungbean cultivars. J Plant Physiol, 2011, 168:807–815
[31]Wani A S, Ahmad A, Hayat S, Tahir I. Is foliar spray of proline sufficient for mitigation of salt stress in Brassica juncea. Environ Sci Pollut Res, 2016, 23: 13413–13423
[32]赵九洲, 胡立盼, 徐志然, 申璐, 胡晓辉. 甜瓜幼苗耐盐碱性及缓解盐碱胁迫γ-氨基丁酸浓度的筛选. 北方园艺, 2014, (9):1–7 Zhao J Z, Hu L P, Xu Z R, Shen L, Hu X H. The screening of melon (Cucumis melo L.) seedling cultivar for salt-alkaline tolerance and the concentration of γ–aminobutyric acid alleviating salt-alkaline stress. North Hortic, 2014, (9): 1–7 (in Chinese with English abstract)
[33]Akhtar J, Ahmad R, Ashraf M Y, Tanveer A, Warrich E A, Oraby H. Influence of exogenous application of salicylic acid on saltstressed mungbean (Vigna radiata): growth and nitrogen metabolism. Pak J Bot, 2013, 45: 119–125
[34]马敬坤, 袁永泽, 欧吉权, 欧阳敏, 鲍世颖, 张楚富. 外源水杨酸对水稻(Oryza sativa L.)幼苗根的NaCl胁迫缓解效应. 武汉大学学报(理学版), 2006, 52: 471–474 Ma J K, Yuan Y Z, Ou J Q, Ou-Yang M, Bao S Y, Zhang C F. Relieving effect of exogenous salicylic acid on rice (Oryza sativa L.)seedling roots under NaCl stress. J Wuhan Univ (Nat Sci Edn), 52:471–474 (in Chinese with English abstract)
[35]Li Z, Yu J, Peng Y, Huang B. Metabolic pathways regulated by abscisic acid, salicylic acid and gamma-aminobutyric acid in association with improved drought tolerance in creeping bentgrass (Agrostis stolonifera). Physiol Plant, 2017, 159: 42–58
[36]Wu L, Hu X, Wang S, Tian L, Pang Y, Han Z, Wu L, Chen Y.Quantitative analysis of changes in the phosphoproteome of maize induced by the plant hormone salicylic acid. Sci Rep-UK,2015, 5: 18155
Effect of Exogenous Salicylic Acid, Proline, and γ-Aminobutyric Acid on Yield of Rice under Salt Stress
SHA Han-*Jing, HU Wen-Cheng, JIA Yan, WANG Xin-Peng, TIAN Xue-Fei, YU Mei-Fang, and ZHAO Hong-Wei
Rice Research Institute, Northeast Agricultural University, Harbin 150030, China
10.3724/SP.J.1006.2017.01677
本研究由国家重点研发计划项目(2016YFD0300104)和黑龙江省重大科技招标项目(GA14B102-02)资助。
This study was supported by the National Key R&D Program (2016YFD0300104) and the Science and Technology Tender Program of Heilongjiang Province (GA14B102-02).
*通讯作者(Corresponding author): 赵宏伟, E-mail: hongweizhao_cool@126.com, Tel: 0451-55190292
联系方式: E-mail: nkxshahanjing@163.com
): 2017-02-16; Accepted(接受日期): 2017-07-23; Published online(网络出版日期): 2017-08-01.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20170801.1839.010.html
