碱胁迫应答基因GsARHP的克隆及转基因紫花苜蓿的耐碱性分析
2017-09-26陈冉冉朱娉慧贾博为宋雪薇王子君李佶娜李强丁晓东朱延明
陈冉冉,朱娉慧,贾博为,宋雪薇,王子君,李佶娜,李强,丁晓东,朱延明
(东北农业大学农业生物功能基因重点实验室,黑龙江 哈尔滨 150030)
碱胁迫应答基因GsARHP的克隆及转基因紫花苜蓿的耐碱性分析
陈冉冉**,朱娉慧**,贾博为,宋雪薇,王子君,李佶娜,李强,丁晓东,朱延明*
(东北农业大学农业生物功能基因重点实验室,黑龙江 哈尔滨 150030)
本研究基于实验室前期野生大豆碱胁迫转录组数据,筛选出一个碱胁迫下上调表达的假定蛋白基因,暂命名为GsARHP(alkali stress related hypothetical protein gene)。首先利用Real-time PCR方法验证了GsARHP基因受碱胁迫诱导表达。生物信息学分析表明,该基因编码一个含有130个氨基酸的亲水蛋白,含有信号肽但无跨膜结构域;构建了GsARHP植物超量表达载体,利用农杆菌介导的子叶节侵染法转化肇东紫花苜蓿,通过PCR,Southern Blot和RT-PCR方法检测获得了3个超量表达GsARHP基因的转基因株系,并对其耐碱性进行了分析。结果表明,在0,100和150 mmol/L NaHCO3处理14 d后,非转基因株系明显萎蔫、黄化甚至死亡,而转基因株系则长势良好;进一步分析其生理指标显示,相对质膜透性与丙二醛含量均显著低于非转基因株系(P<0.01),而叶绿素含量与CAT活性显著高于非转基因株系(P<0.01),说明GsARHP基因的超量表达可以增强紫花苜蓿的耐碱能力。
野生大豆;GsARHP基因;碱胁迫;紫花苜蓿;遗传转化
目前,世界各地主要的农作物减产是由非生物胁迫造成的[1]。植物长期处于不良环境中,会使植物体内产生相关生理代谢反应,致使植物机体产生结构和功能上的改变,严重时甚至会造成不可逆的损伤,导致整个植株死亡[2]。其中,盐碱胁迫是影响植物生长最主要的环境因素之一,大面积的盐碱化土地严重限制了农业可持续发展和生态环境的恢复,制约着全世界作物产量[3]。盐碱对土壤的危害是由于土壤中对作物有害的阴、阳离子富集所致,如CO32-、HCO3-、Cl-等阴离子和Na+、Ca2+等阳离子[4]。土壤内积累大量盐分会引起土壤肥力降低,有机质含量减少,理化性状改变等现象,研究表明土壤盐分的积累,会扰乱植物体内离子平衡,破坏植物正常的新陈代谢机能,阻碍蛋白质的合成与水解,引起可溶性盐离子在体内的积聚,从而产生离子毒害,危及植物的生长发育[5],导致植物代谢受阻,生长发育受到抑制,进一步萎蔫甚至死亡[6]。合理开发利用盐碱化土地成为推动现代农业发展、改善生态环境的重要任务之一[2-7]。目前,关于盐碱胁迫,研究较集中在中性盐(NaCl)对植物的危害[8-13],但是,碱土除了具有中性盐的胁迫特征外,还具有高pH特征,碱土高度离散,湿时膨胀,干时板结,通透性很差,严重妨碍作物的生长发育,比中性盐的危害更严重,因此对碱性盐的研究更具有现实意义。
紫花苜蓿(Medicagosativa)是一种世界性广泛种植的优良牧草,富含蛋白质、多种维生素和矿物质,是营养丰富的饲料植物[14]。紫花苜蓿具有较强的抗逆性,拥有抗寒、耐旱、耐盐碱等特性[1],是改良和开发盐碱地的重要作物之一。牧草作物一般具有发达的根系,在盐碱化土地上种植可以使土壤恶化的理化性质逐渐得到改善,减轻盐碱化土地对植物的危害,可以在培育牧草的过程中达到改善盐碱土地的目的[14-16]。但紫花苜蓿有限的耐碱能力[17],限制了它的栽种范围,所以亟需对其耐碱性状进行改良,培育耐碱性强的紫花苜蓿新品种,增强其在盐碱地区的生长能力,扩大种植范围,以期充分开发利用盐碱化土地资源。采用传统的育种方法,培育耐盐碱新品种进展缓慢耗时耗力。目前,利用基因工程技术向植物导入抗逆相关的目的基因,提高植物的耐盐碱性,已成为提高农牧业生产,培育抗逆新品种的重要途径。
根据实验室对东北野生大豆(Glycinesoja)Gs07256转录组测序数据,碱胁迫下对照组与不同时间点处理组的表达差异,从野生大豆根中筛选出3380个碱胁迫应答基因[18],在分析野生大豆盐碱胁迫应答的差异表达基因时,筛选出一个碱胁迫后上调表达的假定蛋白基因GsARHP,假定蛋白是指因功能未知而未被鉴定的新基因编码的蛋白[19],其在碱胁迫基因表达图谱中呈现上调表达[18],因此推测GsARHP很可能也参与了植物对碱胁迫的响应。
为确定该基因在碱胁迫应答中的作用,本研究以紫花苜蓿为受体材料,构建由强组成型启动子CaMV35S调控的超量表达GsARHP基因的植物表达载体,利用根癌农杆菌EHA105对紫花苜蓿进行遗传转化,获得转基因植株。通过PCR,Southern Blot,RT-PCR的方法鉴定转基因植株。分析转基因植株的生长发育状况和耐碱能力,初步确定该假定蛋白基因在碱胁迫应答中的功能。本研究为进一步研究假定蛋白基因GsARHP在碱胁迫应答调控中的作用机制奠定了重要的理论基础,对于建立紫花苜蓿分子育种技术体系,培育耐碱功能显著的转基因紫花苜蓿新株系,充分利用并改良我国盐碱地资源,具有重要的实际应用价值和广阔的开发前景。
1 材料与方法
1.1实验材料
野生大豆(Gs07256),采自吉林省白城市盐碱地。大肠杆菌DH5α、农杆菌EHA105菌株与植物表达载体pCAMBIA330035Su均由东北农业大学农业生物功能基因重点实验室保存;转化受体为肇东紫花苜蓿,由黑龙江省农业科学院草业研究所提供。
1.2GsARHP基因的克隆与表达模式分析
根据野生大豆与大豆的高度同源性,利用同源克隆技术,根据GsARHP在栽培大豆中的同源基因(Glyma17g07720)序列设计特异的PCR引物(GsARHP-S:5′-ATGGCCGCTACTATGAAG-3′;GsARHP-AS:5′-TTACTTTCCATTACCACCG-3′),以野生大豆cDNA作为模板,通过PCR扩增获得目的基因的完整CDS区。
选取长势一致的3周龄的野生大豆幼苗,置于50 mmol/L NaHCO3条件下处理0,1,3,6,12,24 h后迅速剪取根尖部分,提取总RNA,经反转录得到cDNA,以野生大豆GAPDH基因作为内参,结合基因特异性引物进行Real-time PCR,实验设置3次生物学重复以及3次技术重复,分析目的基因在碱胁迫处理下的表达模式。
1.3GsARHP基因生物信息学分析
通过ORF Finder(http://www.ncbi.nlm.nih.gov/gorf/orfig.cgi)查找序列的开放阅读框,最终确定基因GsARHP完整的全长序列。通过TMHMM Server v.2.0 (http://www.cbs.dtu.dk/services/TMHMM-2.0/)预测蛋白质亲疏水性与蛋白质跨膜结构域;经SignalP4.1(http://www.cbs.dtu.dk/services/SignalP/)预测基因产物信号肽;TargetP(http://www.cbs.dtu.dk/services/TargetP/)预测蛋白质亚细胞定位;分析蛋白一级结构。利用GOR4(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html)预测蛋白质二级结构,MotifScan(http://myhits.isb-sib.ch/cgi-bin/motif_scan)软件寻找蛋白质的结构域;确定该假定蛋白基本特性。
1.4植物表达载体的构建及转基因紫花苜蓿的培育
采用基于USERTM克隆技术的pCAMBIA330035Su载体作为植物超量表达载体。根据GsARHP基因序列设计的基因特异PCR扩增引物,在其上游引物中添加GGCTTAAU接头,在下游引物中添加GGTTTAAU接头。采用PfuTurbo Cx Hotstart DNA polymerase,PCR扩增GsARHP基因[20-21]。构建由烟草花叶病毒CaMV35S强启动子调控,Bar基因作为筛选标记基因的高水平组成型植物表达载体。利用冻融法将植物表达载体导入农杆菌EHA105,用于紫花苜蓿的遗传转化。
采用农杆菌侵染子叶节的方法进行紫花苜蓿的遗传转化[22]。种子经灭菌萌发后,将培养8 d的无菌苗从子叶节下部约1 mm膨大处剪取子叶节,经农杆菌侵染后,接种于含有100 μmol/L乙酰丁香酮的MS培养基上共培养3~5 d,除菌后,将外植体接种到添加草甘膦(0.5 mg/L)的固体筛选培养基上筛选培养,当抗性芽约2 cm长以后,进行生根处理并将其转入生根培养基中诱导生根,2周左右生根,驯化2~3 d后移栽。获得抗性再生植株。
1.5再生植株的分子生物学检测
再生植株的分子生物学检测分为基因水平的PCR检测,Southern Blot检测和转录水平的RT-PCR检测。提取抗性植株的总DNA,利用筛选标记基因Bar基因的特异引物进行PCR检测,阳性对照为含有目的基因的重组质粒DNA,阴性对照为未转基因植株的总DNA,初步确定目的基因已整合入再生植株的基因组中。对PCR阳性的再生植株进行Southern Blot检测,制备基因特异性探针,确定目的基因的整合及基因拷贝数。为检测已整合的目的基因在再生植株中转录水平的表达情况,对Southern Blot阳性植株进行RT-PCR半定量检测,选取Southern Blot阳性转基因植株,紫花苜蓿GAPDH基因作为内参基因(GeneBank登录号为MTR_4G131180),进行转录水平分析。
1.6碱胁迫下转基因植株生理指标检测
实验于2016年8—10月在东北农业大学园艺站温室内进行,设置3次重复。
由于紫花苜蓿异花授粉和生长周期长等特点,利用有性生殖进行扩繁存在耗时及不稳定等问题。因此,本研究采用扦插扩繁的方式来获得扦插苗,选取转基因植株和未转基因植株木质化程度较高且均匀一致的茎段进行扦插扩繁,剪取带有2~3个生长节点的茎段,采用生根粉浸泡2 h处理,待茎段生根后,移栽到含有蛭石∶珍珠岩=1∶1的小钵(直径5 cm,高6 cm,每钵1株)中,1000 lx光照,16 h/8 h的光照/黑暗周期,26 ℃/18 ℃昼夜温度的条件下培养4周后,挑选生长状态一致的转基因和未转基因的扦插苗,分别用含有0,100和150 mmol/L NaHCO3的1/8 Hoagland营养液处理,营养液每2 d更换一次,期间观察记录表型,14 d后取出植株,洗净珍珠岩,利用刻度尺测量植株根长与株高。
采用80%丙酮研磨提取叶绿素,比色法[23]检测叶绿素含量,采用电导法[24-25]检测相对质膜透性。利用硫代巴比妥酸(TBA)比色法[23]测定丙二醛(malondialdehyde,MDA)含量,采用过氧化氢法[26]检测过氧化氢酶(catalase,CAT)含量。
2 结果与分析
2.1GsARHP基因的筛选与克隆
2.1.1基于转录组数据的GsARHP筛选 根据实验室对东北野生大豆Gs07256转录组测序数据,从中选取在NaHCO3胁迫处理下根中表达量显著上调的基因GsARHP,结果见图1A。从图中可以看出,GsARHP在根中胁迫处理6 h时表达量达到最高值。采用Real-time PCR方法分析碱胁迫处理下不同时间点GsARHP基因在野生大豆中转录水平表达量的变化(图1B),以验证转录组测序数据。实时荧光定量PCR分析结果表明,在50 mmol/L NaHCO3胁迫处理下,GsARHP基因根中的表达量在12 h达到峰值。
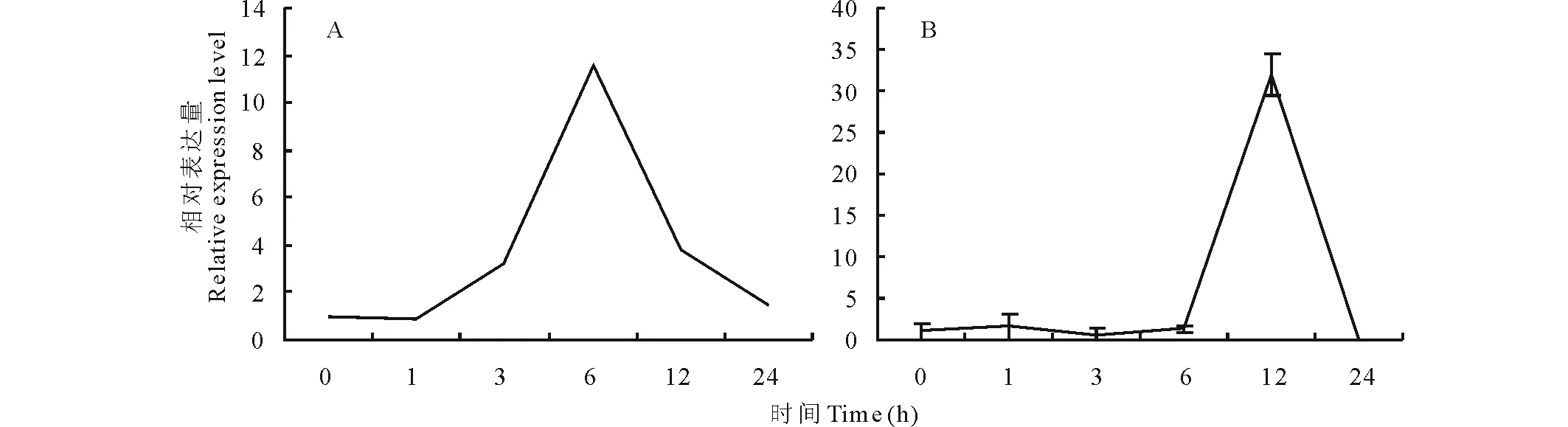
图1 碱胁迫下GsARHP基因表达特性分析Fig.1 GsARHP gene expression characteristics analysis under alkali stress A: GsARHP基因转录组测序结果RNA-seq result of GsARHP; B:GsARHP 基因Real-time PCR结果Real-time PCR result of GsARHP.
与转录组数据结果比较来看,虽然GsARHP基因
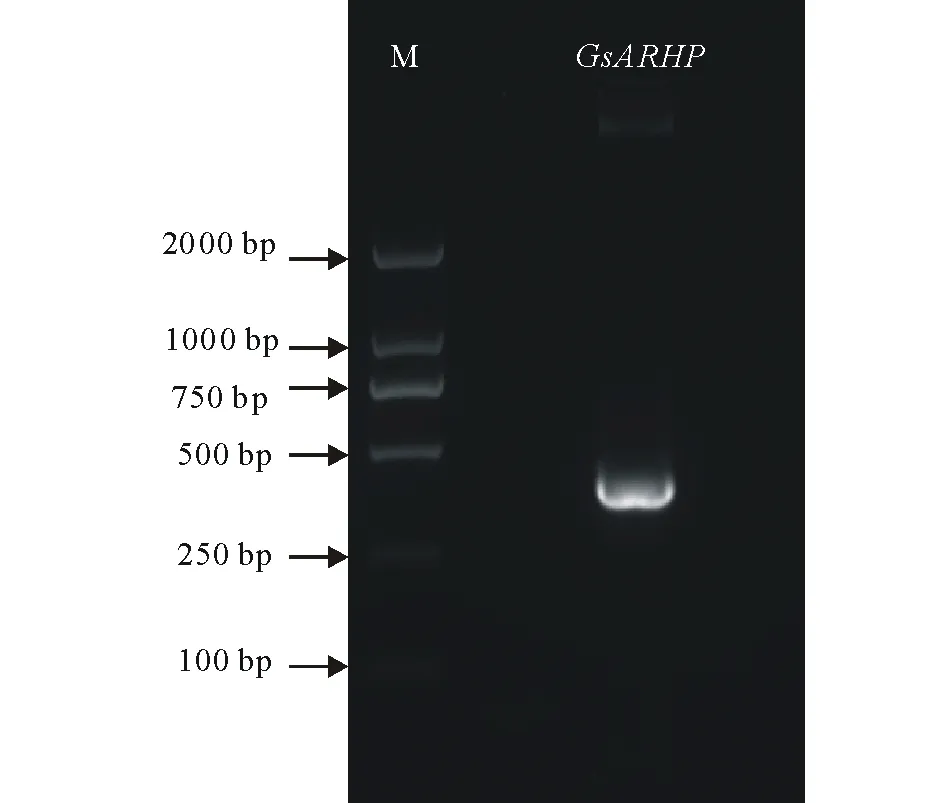
图2 GsARHP基因的PCR扩增Fig.2 PCR amplification of GsARHP gene
对胁迫响应的时间点有所推后,但变化趋势仍然基本一致。确定该基因为碱胁迫应答候选基因。
2.1.2GsARHP基因的克隆 提取野生大豆总RNA,反转录得到cDNA模板,利用目的基因特异性引物,进行PCR扩增。如图2所示,克隆得到长约390 bp的DNA条带(图2)。将回收得到的PCR产物连接到载体pCAMBIA330035Su上,转化大肠杆菌,采用PCR技术鉴定阳性转化子,将阳性克隆送交测序,测序结果表明序列正确,成功的克隆得到GsARHP基因。
2.2GsARHP基因生物信息学分析
2.2.1GsARHP基因序列分析 对GsARHP基因全长序列进行序列分析,根据Phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)上提供全序列分析GsARHP基因全长cDNA序列及推导的氨基酸序列,如图3所示,该基因全长为846 bp,包含127 bp 5′端非编码区序列(5′-UTR)与326 bp 3′端非编码区序列(3′-UTR),将测序所得到的序列在NCBI“ORF finder”预测开放阅读框(ORF),预测最长ORF为393 bp,编码130个氨基酸,与预期结果一致。因此,根据以上结果,认为此克隆序列已包含目的基因GsARHP的完整ORF。可以用于下一步实验。

图3 假定蛋白基因GsARHP全长cDNA序列及推导的氨基酸序列Fig.3 Full-length cDNA sequence and deduced amino acid sequence of GsARHP
2.2.2GsARHP蛋白结构预测 采用ProtParam软件预测蛋白质一级结构。假定蛋白基因共编码130个氨基酸,其中,甘氨酸的含量最高(29.2%),其次为丝氨酸(23.1%)、丙氨酸(14.6%)和亮氨酸(6.2%)。分子质量11.428 kD,等电点7.91,不稳定指数32.13,表明GsARHP为一个稳定蛋白。通过Protscale软件预测氨基酸的亲水性和疏水性,利用Hphob./Kyte&Doolittle算法得到如图4所示结果:该蛋白亲水性氨基酸均匀分布在整个肽链中且多于疏水性氨基酸。证明该基因编码蛋白是一种亲水性蛋白。
经GOR4预测,GsARHP蛋白二级结构由如下3个部分组成:无规卷曲共89个氨基酸,占68.46%,延伸片段共23个氨基酸,占17.69%,α螺旋共18个氨基酸,占13.85%。用TMHMM 2.0预测蛋白质的跨膜结构,表明该蛋白无跨膜结构域,用SignalIP 4.1预测信号肽及可能的切割位点,表明该多肽链含有信号肽,多肽链上信号肽的切割位点可能是第19~20位氨基酸残基。经TargetP预测,该蛋白可能是经分泌通路信号肽引导的胞外蛋白;用MotifScan软件寻找蛋白质的结构域,未发现功能结构域。
2.3GsARHP基因对紫花苜蓿的遗传转化
2.3.1植物表达载体构建 植物表达载体构建如图5,将送交测序并确定构建正确的重组质粒采用冻融法导入农杆菌EHA105中,对转化子进行菌落PCR鉴定,与阳性重组质粒相比,均扩增出约390 bp的片段,表明携带该目的基因的重组质粒已成功导入根癌农杆菌中,可用于下一步的农杆菌介导侵染子叶节法转化紫花苜蓿。
2.3.2GsARHP基因对紫花苜蓿的遗传转化 采用农杆菌介导的子叶节侵染方法对紫花苜蓿进行遗传转化(图6)。经过无菌苗培育、子叶节预培养、农杆菌侵染、共培养、不定芽诱导分化、伸长及草甘膦筛选培养(0.5 mg/L)、生根培养等组织培养阶段,最终获得抗性植株。共得到了37株抗性植株。
2.4转基因紫花苜蓿的分子生物学检测
2.4.1PCR检测与Southern Blot检测 利用筛选标记基因Bar基因引物对抗性植株进行PCR检测,部分PCR鉴定结果如图7所示,转基因植株扩增出约450 bp片段,与阳性对照一致,筛选得到14株PCR阳性植株,初步证明目的基因已整合到紫花苜蓿基因组中。对14株PCR阳性植株进行Southern Blot鉴定(图8),结果显示共获得3株Southern Blot阳性植株,其中#2为3个拷贝,#12为1个拷贝,#21为2个拷贝(图8)。证明目的基因已成功整合到紫花苜蓿基因组中。
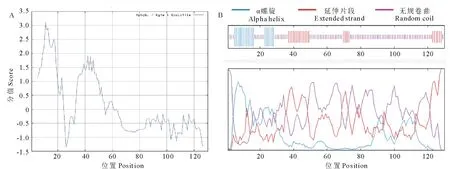
图4 GsARHP蛋白结构预测Fig.4 Prediction of GsARHP protein structure A:GsARHP蛋白疏水性、亲水性分析Hydrophilicity, hydrophobicity analyze of GsARHP;B:GsARHP蛋白二级结构预测 Secondary structure of prediction of GsARHP.
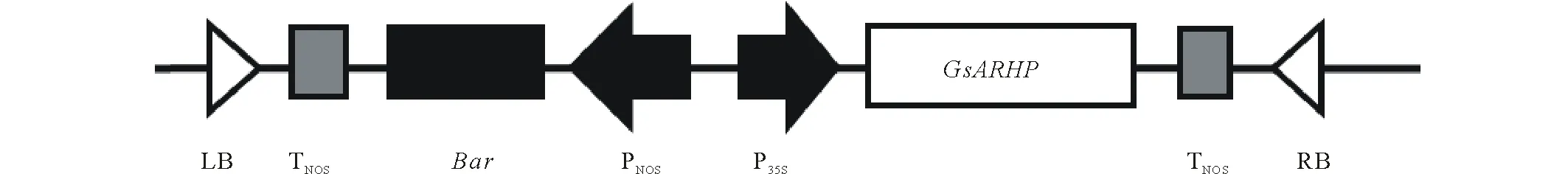
图5 GsARHP植物超量表达载体的构建Fig.5 Construction of GsARHP plant over expression vector LB:左界 Left border;TNOS:T-DNA复制原点 Ori of T-DNA;Bar:筛选标记基因 Selectable marker gene;PNOS:启动子复制原点Ori of promoter;P35S:35S强启动子35S promoter;RB:右边界 Right border.

图6 紫花苜蓿遗传转化及植株再生Fig.6 Genetic transformation and plant regeneration of alfalfa A:8日龄无菌苗Germination 8 days of aseptic seedling;B:农杆菌侵染子叶节Infection;C:共培养Co-cultivation;D:不定芽诱导Adventitious bud induction;E:抗性芽伸长Resistant shoot elongation;F:抗性苗生根驯化Root plantlets and domestication.
2.4.2RT-PCR检测 为确定转基因植株在转录水平的表达情况,对筛选得到的3株Southern Blot阳性植株进行转录水平分析。半定量RT-PCR分析结果显示(图9),3株Southern Blot阳性植株在转录水平均可正常表达。
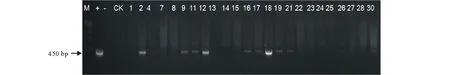
图7 转GsARHP基因抗性植株PCR检测 Fig.7 The PCR identification of resistant plants with GsARHP M:DL 15000;+:阳性对照Positive control;-:水对照Water control;CK:非转基因对照Control check;1~30:转GsARHP基因抗性植株PCR扩增产物 PCR amplification products of transgenic plants with GsARHP gene.

图8 转基因植株的Southern Blot鉴定Fig.8 Southern Blot identification of the transgenic alfalfa
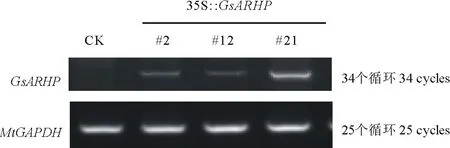
图9 转基因植株的半定量RT-PCR鉴定Fig.9 Semi-RT PCR of transgenic resistant plants with GsARHP
2.5转基因紫花苜蓿碱胁迫生物学分析

图10 转GsARHP基因紫花苜蓿碱胁迫表型分析Fig.10 Alkali stress phenotype analysis of GsARHP transgenic alfalfa
2.5.1碱胁迫下转基因植株的表型分析 碱胁迫会导致植物产生一系列生理响应,抑制其各器官及组织的生长,从而抑制植物的生长发育,对植物造成伤害。为研究碱胁迫对转基因紫花苜蓿生长发育的影响,本研究选取Southern Blot检测呈阳性的2个转基因植株#2和#21及未转基因(CK)紫花苜蓿进行扦插扩繁,获得扦插苗后,以100和150 mmol/L NaHCO3作为胁迫浓度,模拟碱胁迫条件,对转基因与未转基因株系进行耐碱性分析。结果显示在正常未处理条件下,转GsARHP基因紫花苜蓿与非转基因紫花苜蓿的长势良好,无明显差异(图10)。在经150 mmol/L NaHCO3处理14 d后,与明显发生萎蔫、黄化的非转基因对照株系相比,转基因株系#2和#21生长良好且能维持正常的生物学形态(图10)。对转基因与未转基因株系的生长发育情况进行分析得知,随着碱胁迫处理浓度的增加,所有株系的株高与根长均呈现不同程度的下降趋势;经统计,在150 mmol/L NaHCO3处理条件下,2个转基因株系的株高和根长均显著高于非转基因株系(P<0.01)(图11)。由此可以说明与非转基因株系相比,转GsARHP基因紫花苜蓿在碱胁迫下能更好地维持生长发育,抵御碱胁迫造成的危害。
2.5.2碱胁迫下转基因植株的生理指标检测 通过检测转基因与对照非转基因株系的叶绿素含量,如图12A所示,发现随着NaHCO3胁迫处理浓度的增加,转GsARHP基因紫花苜蓿株系和对照株系的叶绿素含量均呈现出不同程度的降低趋势,但在相同浓度的NaHCO3胁迫处理条件下,转基因株系的叶绿素含量高于对照株系,尤其在150 mmol/L NaHCO3胁迫处理条件下,与对照株系相比,转基因株系#21的叶绿素含量显著升高(P<0.01)(图12A),显然转GsARHP基因紫花苜蓿株系在碱胁迫条件下能更好维持叶绿体的稳定性,从而维持植株正常的生长发育。
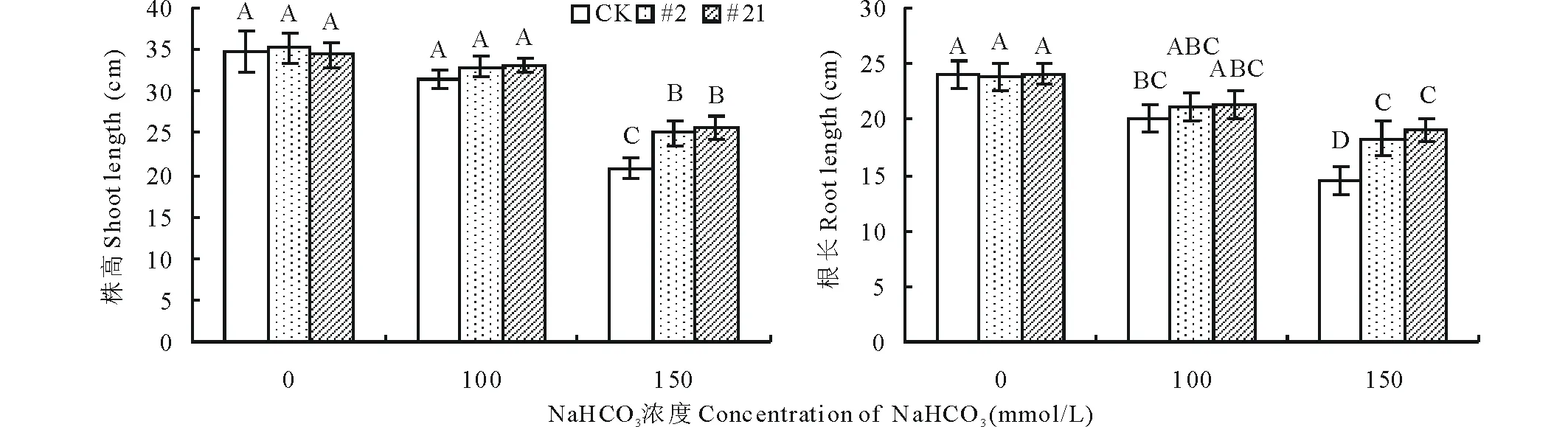
图11 碱胁迫对紫花苜蓿株高和根长的分析Fig.11 Shoot and root length analysis of transgenic alfalfa with GsARHP under alkali stress 图柱上方不同大写字母表示在 P<0.01水平上差异显著。下同。Bars with different capital letters are significant differences at P<0.01.The same below.
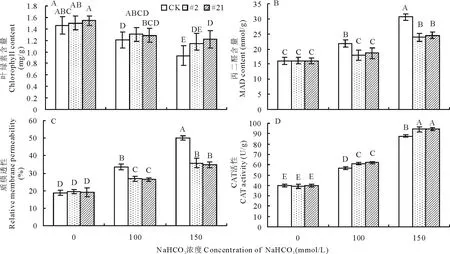
图12 碱胁迫下转GsARHP基因紫花苜蓿生理指标检测Fig.12 Physiological indices analysis of transgenic alfalfa with GsARHP under alkali stress
通过检测转基因与未转基因株系MDA含量(图12B)发现,随NaHCO3胁迫处理浓度的增加,2个转GsARHP基因紫花苜蓿株系的MDA含量略有上升,但由数据分析可知,非转基因株系MDA含量显著高于转基因株系(P<0.01),从而说明超量表达GsARHP基因能降低NaHCO3胁迫对细胞膜造成的脂质过氧化伤害,保持细胞膜正常生理功能。
检测转基因与对照非转基因株系的相对质膜透性,由图12C所示,在NaHCO3胁迫处理下,相对质膜透性在转GsARHP基因植株和对照植株中均有升高,但2个转基因株系相对质膜透性的增幅均显著低于对照株系(P<0.01),说明在NaHCO3胁迫处理条件下,超量表达GsARHP基因可降低碱胁迫对细胞膜的损伤,维持细胞膜完整性从而提升耐碱能力。
CAT活性的测定如图12D所示,在NaHCO3胁迫处理下,转基因和非转基因植株的CAT活性均提高,而在2个转基因植株中CAT活性显著高于非转基因植株(P<0.01),由此可见,GsARHP基因超量表达增强了转基因植株对自由基清除能力,从而降低了碱胁迫下自由基的积累,减少了自由基对植株核酸、蛋白质以及细胞膜的伤害。
3 讨论
盐碱胁迫严重制约着作物的生长发育,影响其产量及质量,Webb[27]首次由农杆菌介导的紫花苜蓿转化获得成功之后,在以后的十多年时间里,紫花苜蓿转基因取得了很大的进展[28],Winicov等[29]将ALfinl基因转入苜蓿中提高了盐诱导基因MsPRP2的表达,提高了转基因植株的耐盐性,Liu等[30]将ScNHX1基因和ScVP基因(液泡无机焦磷酸酶)转入苜蓿,得到了具有很强耐盐碱性的转基因苜蓿。Wang等[31]将GsGST14在紫花苜蓿中超量表达,提升了转基因植株中GST的酶活力,增强了转基因植株对碱的抗性。盛慧等[32]将抗渗透胁迫信号传导过程中的关键基因DREB2A通过农杆菌介导法对紫花苜蓿进行遗传转化,获得了大量转基因植株。目前,利用转基因技术向苜蓿中导入具有优良性状的目的基因来提升转基因苜蓿的耐盐碱性状,已成为改良农业和畜牧业生产的重要途径[33]。本研究选用从野生大豆中筛选出的碱胁迫应答候选基因GsARHP,转化肇东紫花苜蓿,期望提高转基因紫花苜蓿的耐碱能力。
野生大豆生长在极其恶劣的生态环境中,具有耐盐碱,抗寒,抗病等优良特性,含有丰富的耐逆基因,是我国珍贵的种质资源,本研究从野生大豆碱胁迫转录组数据中筛选得到一个碱胁迫下上调表达的假定蛋白基因GsARHP,假定蛋白是经测序翻译后得到的功能未知的蛋白,推测可能与野生大豆抗盐碱的生理活动有关,因此对该蛋白进行了生物信息学分析,发现该多肽链不具备跨膜结构域,但含有信号肽,可能是经分泌通路信号肽引导的胞外亲水蛋白,为稳定蛋白;但是并未发现该蛋白的功能结构域。为该基因的分子机制研究奠定了重要的理论基础。进一步在肇东紫花苜蓿中超量表达GsARHP,验证了该基因在植物碱胁迫应答反应中的功能。在37株再生植株中经检测得到了3株GsARHP基因在转录水平超量表达的植株。
在对转基因植株进行检测方面,PCR检测法是现在使用最普遍也是最简便的实验检测方法[34],研究表明,利用常规PCR法检测重组载体上标记基因如GUS基因[35],35S启动子[36]等部分片段,最终证明整合了GUS基因,35S启动子的转基因植株中可能不含有目的基因片段[37-38],这可能与外源基因的整合机制相关,如染色体的重组、异位等现象的发生致使目的基因片段的丢失[39-40],本实验结果也证实了这一说法;本研究采取Southern Blot方法设计可与目的基因特异性结合的特异性探针,保证了转基因阳性植株的正确筛选,同时对整合目的基因的阳性植株进行转录水平的检测,确定已整合的目的基因在转录水平上成功表达可用于进一步基因功能验证。
碱胁迫条件下,植物会在细胞内产生并积累大量的活性氧,破坏了植物体内活性氧代谢平衡,从而对核酸、蛋白质等生物大分子产生不利影响;导致光合作用相关元件损伤,影响植物光合作用,从而导致植物失绿甚至死亡;致使膜系统发生过氧化现象,改变质膜透性,产生丙二醛(MDA),但植物本身存在的活性氧清除保护机制可在一定程度上清除活性氧自由基,维持细胞内活性氧代谢平衡。以上生理指标均可用来指示植物在碱胁迫下受到的损伤情况[41-42]。本研究结果表明GsARHP基因超量表达的紫花苜蓿,在碱胁迫条件下,能够保持叶绿素含量,维持光合作用积累有机物来保证植株正常的生长发育;维持细胞膜的选择透过性等生理活性,减少丙二醛等有害物质的产生,增强了转基因紫花苜蓿对活性氧自由基的清除能力,保护细胞避免因H2O2累积而产生伤害,从而维持了碱胁迫下转基因植株的氧化还原平衡。说明GsARHP基因的超量表达可以提高紫花苜蓿的耐碱能力,具体分子机制还需进一步实验验证。本研究为深入研究GsARHP基因的功能和培养紫花苜蓿耐盐碱新品种奠定了基础。
References:
[1] Qi Z Q, Yu Y X, Hu Y G,etal. Current status and future tasks of theMedicagosativaindustry in China. Acta Prataculturae Sinica, 2008, 17(1): 107-113. 戚志强, 玉永雄, 胡跃高, 等. 当前我国苜蓿产业发展的形势与任务. 草业学报, 2008, 17(1): 107-113.
[2] Hua Y, Cai H, Bai X,etal. Research progress on plant salt stress tolerance genetic engineering. Journal of Northeast Agricultural University, 2010, 41(10): 150-156. 化烨, 才华, 柏锡, 等. 植物耐盐基因工程研究进展. 东北农业大学学报, 2010, 41(10): 150-156.
[3] Peng J Q, Liu Y Q, Yang Y F,etal. Importance and technology progress in containing the soil salinization and desertification. Tianjin Agricultural Sciences, 2008, 14(4): 26-29. 彭津琴, 刘永强, 杨玉芳, 等. 遏制土壤盐碱化、荒漠化的必要性及技术进展. 天津农业科学, 2008, 14(4): 26-29.
[4] Tian C X, Zhang Y M, Wang K,etal. The anatomical structure responses in alfalfa to salinity-alkalinity stress of NaHCO3. Acta Prataculturae Sinica, 2014, 23(5): 133-142. 田晨霞, 张咏梅, 王凯, 等. 紫花苜蓿组织解剖结构对NaHCO3盐碱胁迫的响应. 草业学报, 2014, 23(5): 133-142.
[5] Hasegawa P M, Bressan R A, Zhu J K,etal. Plant cellular and molecular responses to high salinity. Plant Biology, 2000, 51(51): 463-499.
[6] Yang X Y, Yang J S, Liu G M,etal. Changes in soil fertilities and crop growth after transferring paddy soil to upland soil. Chinese Journal of Soil Science, 2006, 37(4): 675-679. 杨晓英, 杨劲松, 刘广明, 等. 盐碱地稻田旱作后土壤肥力变化及其对作物生长的影响. 土壤通报, 2006, 37(4): 675-679.
[7] Li H W, Ma D M, Xu X,etal. Advance and prospects on salt-tolerant transgenic alfalfa. Northern Horticulture, 2012, (19): 184-188. 李会文, 麻冬梅, 许兴, 等. 苜蓿耐盐碱转基因研究进展与展望. 北方园艺, 2012, (19): 184-188.
[8] Das-Chatterjee A, Goswami L, Maitra S,etal. Introgression of a novel salt-tolerant L-myo-inositol 1-phosphate synthase fromPorteresiacoarctata(Roxb.) Tateoka (PcINO1) confers salt tolerance to evolutionary diverse organisms. FEBS Letters, 2006, 580(16): 3980-3988.
[9] Jing Y X, Yuan Q H. Effects of salt stress on seedling growth of alfalfa (Medicagosativa) and ion distribution in different alfalfa organs. Acta Prataculturae Sinica, 2011, 20(2): 134-139. 景艳霞, 袁庆华. NaCl胁迫对苜蓿幼苗生长及不同器官中盐离子分布的影响. 草业学报, 2011, 20(2): 134-139.
[10] Jin T, Chang Q, Li W,etal. Stress-inducible expression of GmDREB1 conferred salt tolerance in transgenic alfalfa. Plant Cell, Tissue and Organ Culture (PCTOC), 2010, 100(2): 219-227.
[11] Liu J, Cai H, Liu Y,etal. A study on physiological characteristics and comparison of salt tolerance of twoMedicagosativaat the seedling stage. Acta Prataculturae Sinica, 2013, 22(2): 250-256. 刘晶, 才华, 刘莹, 等. 两种紫花苜蓿苗期耐盐生理特性的初步研究及其耐盐性比较. 草业学报, 2013, 22(2): 250-256.
[12] Zhang L Q, Zhang F Y, Hasi Agula. Research progress on alfalfa salt tolerance. Acta Prataculturae Sinica, 2012, 21(6): 296-305. 张立全, 张凤英, 哈斯阿古拉. 紫花苜蓿耐盐性研究进展. 草业学报, 2012, 21(6): 296-305.
[13] Jiang J, Yang B L, Xia T,etal. Genetic diversity analysis of salt tolerant germplasm resources of alfalfa. Acta Prataculturae Sinica, 2011, 20(5): 119-125. 姜健, 杨宝灵, 夏彤, 等. 紫花苜蓿耐盐种质资源的遗传多样性分析. 草业学报, 2011, 20(5): 119-125.
[14] Wang J F, Li Y Q, Guo Y Q. Effects of saline alkali soil on saline alkali soil. Shangdong Journal of Animal Science and Veterinary Medicine, 1999, (5): 20-21. 王金芬, 李玉芹, 郭玉泉. 耐盐碱牧草改良盐碱土的效果. 山东畜牧兽医, 1999, (5): 20-21.
[15] Wang X P, Li W Q. A report on the quality inspection and analysis of several different alfalfa varieties. Contemporary Animal Husbandry, 2004, (5): 39. 王学鹏, 李文全. 几个不同品种紫花苜蓿质量检验分析报告. 当代畜牧, 2004, (5): 39.
[16] Tian F P, Wang S M, Guo Z G,etal. Relationship between proline content and water content, single plant dry matter, and drought resistance of alfalfa. Pratacultural Science, 2004, 21(1): 3-6. 田福平, 王锁民, 郭正刚, 等. 苜蓿脯氨酸含量和含水量、单株干质量与抗旱性的相关性研究. 草业科学, 2004, 21(1): 3-6.
[17] Zhang H N, Li X J, Li C D,etal. Effects of overexpression of wheat superoxide dismutase (SOD) genes on salt tolerant capability in Tobacco. Acta Agronomica Sinica, 2008, 34(8): 1403-1408. 张海娜, 李小娟, 李存东, 等. 过量表达小麦超氧化物歧化酶(SOD)基因对烟草耐盐能力的影响. 作物学报, 2008, 34(8):1403-1408.
[18] Duanmu H, Wang Y, Bai X,etal. Wild soybean roots depend on specific transcription factors and oxidation reduction related genes in response to alkaline stress. Functional & Integrative Genomics, 2015, 15(6): 651-660.
[19] Wu P, Liu T, Qin G X,etal. Full-length cDNA cloning and characterization of a hypothetic protein gene responsive to the infection of bombyxmoricytoplasmic polyhedrosis virus. Science of Sericulture, 2010, 36(6): 937-943. 吴萍, 刘挺, 覃光星, 等. 家蚕质型多角体病毒感染应答的假定蛋白基因全长cDNA克隆与表达特征分析. 蚕业科学, 2010, 36(6): 937-943.
[20] Bitinaite J. USERTMfriendly DNA engineering and cloning method by uracil excision. Nucleic Acids Research, 2007, 35(6): 1992-2002.
[21] Geuflores F, Noureldin H H, Nielsen M T,etal. USER fusion: a rapid and efficient method for simultaneous fusion and cloning of multiple PCR products. Nucleic Acids Research, 2007, 35(7): e55.
[22] Jones H D, Wilkinson M, Doherty A,etal. High throughputAgrobacteriumtransformation of wheat: a tool for functional genomics[C]//Wheat Production in Stressed Environments. Proceedings of the 7th International Wheat Conference, 2007: 693-699.
[23] Cortazar B, Koydemir H C, Tseng D,etal. Field quantification of plant chlorophyll content using Google Glass[C]//SPIE BiOS. International Society for Optics and Photonics, 2015.
[24] Liu T Z, Zhao X P. Study of apricot leaves plasma membrane permeability by electric conductivity method. Journal of Hebei Agricultural Sciences, 2008, 12(1): 33-34. 刘铁铮, 赵习平. 电导法测定杏叶片细胞质膜相对透性的研究. 河北农业科学, 2008, 12(1): 33-34.
[25] Zhang T X, Lin Z J, Cao M H,etal. Study on determination of pepper leaf cell membrane permeability of the electrical conductivity method. Fujian Science & Technology of Tropical Crops, 2013, (1): 19-21. 张天翔, 林宗铿, 曹明华, 等. 电导率法测定甜椒叶片细胞质膜相对透性的研究. 福建热作科技, 2013, (1): 19-21.
[26] Alt D. The cat-method for the chemical analysis of horticultural substrates (refereed). Acta Horticulturae, 1997, 450(450): 87-96.
[27] Webb B L. Response of seedling alfalfa (Medicagosativa) to four postemergence herbicides. Weed Technology, 1991, 5(4): 736-738.
[28] Zhang Y, Liu J, Zhou Y,etal. Enhanced phytoremediation of mixed heavy metal (mercury)-organic pollutants (trichloroethylene) with transgenic alfalfa co-expressing glutathione S-transferase and human. Journal of Hazardous Materials, 2013, 260(6): 1100-1107.
[29] Winicov I, Bastola D R. Transgenic overexpression of the transcription factor alfin1 enhances expression of the endogenousMsPRP2 gene in alfalfa and improves salinity tolerance of the plants. Plant Physiology, 1999, 120(2): 473-480.
[30] Liu L, Fan X D, Wang F W,etal. Coexpression ofScNHX1 andScVPin transgenic hybrids improves salt and saline-alkali tolerance in alfalfa (MedicagosativaL.). Journal of Plant Growth Regulation, 2013, 32(1): 1-8.
[31] Wang Z Y, Song F B, Cai H,etal. Over-expressing GsGST14 fromGlycinesojaenhances alkaline tolerance of transgenicMedicagosativa. Biologia Plantarum, 2012, 56(3): 1-5.
[32] Sheng H, Zhu Y M, Li J,etal. Genetic transformation ofDREB2Agene into alfalfa. Pratacultural Science, 2007, 24(3): 40-45. 盛慧, 朱延明, 李杰, 等.DREB2A基因对紫花苜蓿遗传转化的研究. 草业科学, 2007, 24(3): 40-45.
[33] Yang Z, Zhu Y M, Bai X,etal. Over-expressing GsCBRLK/SCMRP enhances alkaline tolerance and methionine content in transgenicMedicagosativa. Acta Agronomica Sinica, 2014, 40(3): 431.
[34] Yue Y F, Wu G, Wu Y H,etal. Development of qualitative PCR method targeting marker genes in transgenic plants. Chinese Journal of Oil Crop Sciences, 2011, 33(3): 280-289. 岳运锋, 吴刚, 武玉花, 等. 转基因植物中标记基因定性PCR检测方法研究. 中国油料作物学报, 2011, 33(3): 280-289.
[35] Wang M, Zhang Q, Liu F C,etal. Family-wide expression characterization ofArabidopsisbeta-carbonic anhydrase genes using qRT-PCR and Promoter::GUS fusions. Biochimie, 2014, 97(1): 219-227.
[36] Shang Z M, Gao H W, Wang X L,etal. Extraction of bean DNA and detection of transgenic CaMV35S by PCR method. Journal of Heze University, 2006, 28(2): 73-75. 尚智美, 高宏伟, 王学亮, 等. 大豆样品中DNA的提取与外源基因CaMV35S的PCR检测. 菏泽学院学报, 2006, 28(2): 73-75.
[37] García-Pérez R D, Houdt H V, Depicker A. Spreading of post-transcriptional gene silencing along the target gene promotes systemic silencing. Plant Journal, 2004, 38(4): 594-602.
[38] Wang H H, Wu S J, Li F F,etal. Transgene silencing caused by 35S promoter methylation in upland cotton (Gossypiumhirsutum). Cotton Science, 2008, 20(4): 274-280. 王海海, 吴慎杰, 李飞飞, 等. 35S启动子甲基化引起棉花转基因沉默. 棉花学报, 2008, 20(4): 274-280.
[39] Peng H Y, Li-Jie Y U. Integration pattern and character of exogenous genes in transgenic plants. Heilongjiang Agricultural Sciences, 2011, 95(2): 509-513.
[40] Atsuo K, Yu H, Satoshi O,etal. Site-specific integration of exogenous genes using genome editing technologies in zebrafish. International Journal of Molecular Sciences, 2016, 17(5): 727.
[41] Tang L, Cai H, Ji W,etal. Overexpression of GsZFP1 enhances salt and drought tolerance in transgenic alfalfa (MedicagosativaL.). Plant Physiology & Biochemistry, 2013, 71(71C): 22-30.
[42] Sun M, Sun X, Zhao Y,etal. Ectopic expression of GsPPCK3 and SCMRP inMedicagosativaenhances plant alkaline stress tolerance and methionine content. Plos One, 2013, 9(2): e89578.
IsolationofGsARHPfromGlycinesojaanditsfunctionalanalysisintransgenicalfalfaunderalkalinestress
CHEN Ran-Ran**, ZHU Ping-Hui**, JIA Bo-Wei, SONG Xue-Wei, WANG Zi-Jun, LI Ji-Na, LI Qiang, DING Xiao-Dong, ZHU Yan-Ming*
KeyLaboratoryofAgriculturalBiologicalFunctionalGenes,NortheastAgriculturalUniversity,Harbin150030,China
GsARHP, which encodes hypothetical protein up-regulated by alkaline stress, was identified from RNA-seq data of wild soybean (Glycinesoja) at the early stage of the salt stress response. Real-time PCR analyses showed thatGsARHPwas induced by alkali stress. A bioinformatics analysis showed that the gene encodes a hydrophilic protein consisting of 130 amino acids, with a signal peptide but no transmembrane domain. A plant overexpression vector was constructed and transformed into alfalfa byAgrobacteriumtumefaciens-mediated cotyledonary node infection. Three transgenic lines were identified by PCR, Southern blotting, and RT-PCR analyses. The non-transgenic lines became wilted and yellow or even died under 0, 100, and 150 mmol/L NaHCO3treatment for 14 days, while the transgenic lines grew well under the same conditions. Further analyses showed that the transgenic lines had significantly lower relative plasma membrane permeability and malondialdehyde (MDA) content than did non-transgenic lines (P<0.01), and significantly higher chlorophyll content and CAT activity (P<0.01). These results indicated that overexpression ofGsARHPcould enhance the alkali resistance of alfalfa.
Glycinesoja;GsARHPgene; alkali stress; alfalfa; genetic transformation
10.11686/cyxb2016464
http://cyxb.lzu.edu.cn
陈冉冉, 朱娉慧, 贾博为, 宋雪薇, 王子君, 李佶娜, 李强, 丁晓东, 朱延明. 碱胁迫应答基因GsARHP的克隆及转基因紫花苜蓿的耐碱性分析. 草业学报, 2017, 26(9): 92-103.
CHEN Ran-Ran, ZHU Ping-Hui, JIA Bo-Wei, SONG Xue-Wei, WANG Zi-Jun, LI Ji-Na, LI Qiang, DING Xiao-Dong, ZHU Yan-Ming. Isolation ofGsARHPfromGlycinesojaand its functional analysis in transgenic alfalfa under alkaline stress. Acta Prataculturae Sinica, 2017, 26(9): 92-103.
2016-12-08;改回日期:2017-02-11
国家自然科学
基金项目(31171578),黑龙江省高校科技创新团队建设计划项目(2011TD005)和东北农业大学学科团队建设项目—团队1资助。
陈冉冉(1991-),女,河北邯郸人,在读硕士。E-mail:crr0620@126.com。朱娉慧(1990-),女,黑龙江齐齐哈尔人,在读硕士。E-mail:zhupinghui@outlook.com。**共同第一作者These authors contributed equally to this work.*通信作者Corresponding author. E-mail: ymzhu2001@neau.edu.cn
