交配对广聚萤叶甲成虫耐饥与耐热能力的影响
2017-09-16赵晨晨万方浩周忠实
赵晨晨,马 超,张 燕,2,万方浩,周忠实*
(1.中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193;2.广西农业环境与农产品安全重点实验室培育基地,广西530005)
交配对广聚萤叶甲成虫耐饥与耐热能力的影响
赵晨晨1,马 超1,张 燕1,2,万方浩1,周忠实1*
(1.中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193;2.广西农业环境与农产品安全重点实验室培育基地,广西530005)
昆虫交配是一个消耗能量的过程,因此交配后可能对其抗逆能力造成一定的影响。广聚萤叶甲是恶性杂草-豚草的专一性天敌,正常情况下,成虫耐饥饿和耐热能力较强。为明确交配对广聚萤叶甲成虫抗逆能力的影响,本研究观察在一次交配处理后,广聚萤叶甲在饥饿条件下或高温条件下继续存活时间。结果发现,在不提供食物的情况下,交配对雄虫的寿命影响不大,但雌虫的寿命(8.4 d±0.1 d)显著短于对照雌虫(10.8 d±0.1 d)。在高温状态下,无论取食与否,交配都会显著增加广聚萤叶甲雌雄成虫的耐热能力,尤其以雌虫表现更为明显(处理雌虫和对照雌虫存活时间分别为:取食(6.15 d±0.14 d,4.95 d±0.13 d)、不取食(5.55 d±0.13 d,4.81 d±0.11 d)。可见,交配降低广聚叶甲雌虫的饥饿能力,但成虫的耐热能力显著加强。
交配;广聚萤叶甲;耐饥;耐热
豚草AmbrosiaartemisiifoliaL.,菊科Asteraceae,豚草属AmbrosiaL,又称普通豚草(Crockett, 1977)、美洲艾等,一年生草本植物,原产自北美中北部(Kovalev, 1989),是一种世界性恶性杂草。自30年代豚草入侵我国以来(Duan and Chen, 2000),已扩繁至我国大部分省市(EPPO, 2014)。豚草素有“植物杀手”之称,其株系高大、根系发达、营养体巨大,可与农作物争光和争夺水肥,且植株体内存在化感物质,具有极强的化感作用,抑制本地植物与农作物生长(Bruckneretal., 2001),因此,对农牧业造成巨大的经济损失(Clewisetal., 2001)。豚草花粉是引起人体一系列过敏性变态症状-枯草热的主要病原,每年开花时节,过敏体质者多会出现喷嚏、哮喘、呼吸急促等过敏症状(Dechamp, 1999),严重者常会引发肺气肿而导致死亡(Gerberetal., 2011)。目前,豚草生物防治被认为是一种最为理想和有效的途径(周忠实等,2008)。在众多天敌,广聚萤叶甲Ophraellacommuna是一种最为有效,且非常安全的天敌,其幼虫和成虫“火烧状”蚕食豚草,对豚草控制效果非常显著。
在自然界,食物和温度是影响天敌种群建立的重要因子。温度决定了昆虫的存活、种群动态和分布(Nicolson and Chown, 2004; Jolivetetal., 2004; Angilletta, 2009)。环境中温度的变化会对昆虫的耐热生理(Renaultetal., 2004; Bozinovicetal., 2011; Marshall and Sinclair, 2012)、传粉(Charabidzeetal., 2008)和生物防治(Butler and Trumble, 2010; Colinet and Hance, 2010)等行为造成影响。一般而言,昆虫中食物缺乏时,常会表现出一定的耐饥饿能力,使其种群度过食物匮乏时期(Rion and Kawecki, 2007)。昆虫交配是两性昆虫繁殖后代必不可少的环节,是一个消耗能量的行为过程,因此交配后,昆虫常会表现出抗逆能力下降的现象(Service, 1989),但是有些昆虫却表现除了更高的抗逆能力(Rushetal., 2007)。
鉴于此,本文旨在了解广聚萤叶甲成虫交配后,其成虫耐饥饿和耐热能力是否受到影响?这一科学问题的解答,有助于解答夏季高温以及食物短缺条件下,交配是否影响广聚萤叶甲成虫种群的发展,为其更好的利用提供科学依据。
1 材料与方法
1.1 供试昆虫及饲养方法
供试寄主:豚草种子于2015年10月收集于河北省廊坊市中国农业科学院科研中试基地,并于2016年6月-11月于中国农业科学院植物保护研究所温室(26℃±1℃、L ∶D=14 h ∶10 h)内分批播种干营养钵中,单株生长至10 cm时,带土分别移栽至方形塑料花盆1(长12 cm、高10 cm)、待植物生长至40-50 cm时放置于养虫笼内用于昆虫饲养。
供试虫源:广聚萤叶甲成虫采集于2016年7月河北省廊坊市中国农业科学院科研中试基地,经体视解剖镜辨别雌雄后饲养于养虫笼(40 cm×60 cm×60 cm)笼罩的豚草植株上(每个笼子6株豚草,每株豚草3对成虫)。待成虫产卵后,移出带卵豚草。每株豚草保留30粒左右卵,放置新的养虫笼内,将卵饲养至成虫。饲养环境26℃±1℃,RH 70±10%,L ∶D=14 h ∶10 h。当成虫生长至2日龄时,选择个体大小均一的成虫用于实验。
1.2 试验方法
1.2.1交配对成虫耐饥能力的影响
上述成虫(见1.1)提供豚草取食24 h后,进行配对,当成虫交配时,记录交配时间,给其统一交配40 min(不到40 min的个体剔除,够40 min将其分开),交配后的雌雄成虫分开,分别放入养虫盒(9 cm×15 cm×25 cm,盒顶部一个长方形气窗5 cm×12.5 cm、盒盖三个圆形气窗半径为 7 cm,由40目纱网制作)内,用棉球提供清水;实验以未交配同日龄的成虫作为对照。然后,逐日观察交配和未交配雌虫和雄虫的存活数量,并逐日记录成虫寿命和产卵数量。每处理设5次重复,每重复分别处理20头雌虫和20头雄虫。
1.2.2交配对成虫耐热能力的影响
上述成虫(见1.1)提供豚草取食24 h后,进行配对,当成虫交配时,记录交配时间,给其统一交配40 min(不到40 min的个体剔除,够40 min把其分开),交配后的雌雄成虫分开,分别放入养虫盒内,用棉球提供清水;实验以未交配同日龄的成虫做为对照。同时设置另一组试验,广聚萤叶甲在养虫盒内提供新鲜豚草;试验以未交配同日龄同样喂食的成虫作为对照。处理后第 5天,交配、未交配雌虫和雄虫分别放到40℃人工气候箱内处理,逐日观察记录死亡数量,直至全部成虫死亡。每处理设5次重复,每重复分别处理40头雌虫和40头雄虫。
1.2.3数据分析和图像处理
数据采用SAS 9.4统计软件(SAS Institute)中2*2*2复因子试验统计分析方法(LSD法)和SPSS 18.0中的probit进行1统计分析。图像使用originPro 2015进行处理分析。
2 结果与分析
2.1 交配对成虫耐饥能力的影响
从存活时间可以看出,雌雄和饥饿与否组合处理显著影响广聚萤叶甲成虫的耐饥能力(F=34.95,df=7,P<0.0001),未交配的雌成虫耐饥力最强,其次为交配和未交配雄虫,耐饥力最弱的是交配雌虫(表1)。在饥饿条件下,无论交配与否,雌雄成虫的最短存活时间相同,但是对照雄虫的最长存活时间稍长于其他处理。由此可见,雌虫的耐饥力强于雄虫;交配不影响雄虫的耐饥力,但显著降低了雌虫的耐饥力。

表1 广聚萤叶甲成虫耐饥饿时间及饥饿半致死时间(T50)和饥饿致死时间(T95)
注:不同小写字母表示差异显著(P<0.05),下表同。Note:The different letter in same line means significant difference (P<0.05).The same below.
从试验结果可以看出,广聚萤叶甲成虫的完全饥饿状态下的存活率都随着饥饿时间的增加而降低,除交配雌虫的耐饥饿曲线呈直线下降外,其他处理的耐饥饿曲线均呈现出弓形(图1)。饥饿状态下,交配雌虫在第7天、第10天出现较高的死亡率(23.50%、30.50%),其他处理存活率下降平缓,交配雄虫在第10和第11天出现较高的阶段死亡率(28.00%、33.50%),对照雌虫在第11、12天出现较高的阶段死亡率(32.00%、33.00%),对照雄虫在第10至第12天的阶段死亡率较高(29.00%、20.00%、25.00%)(图1)。
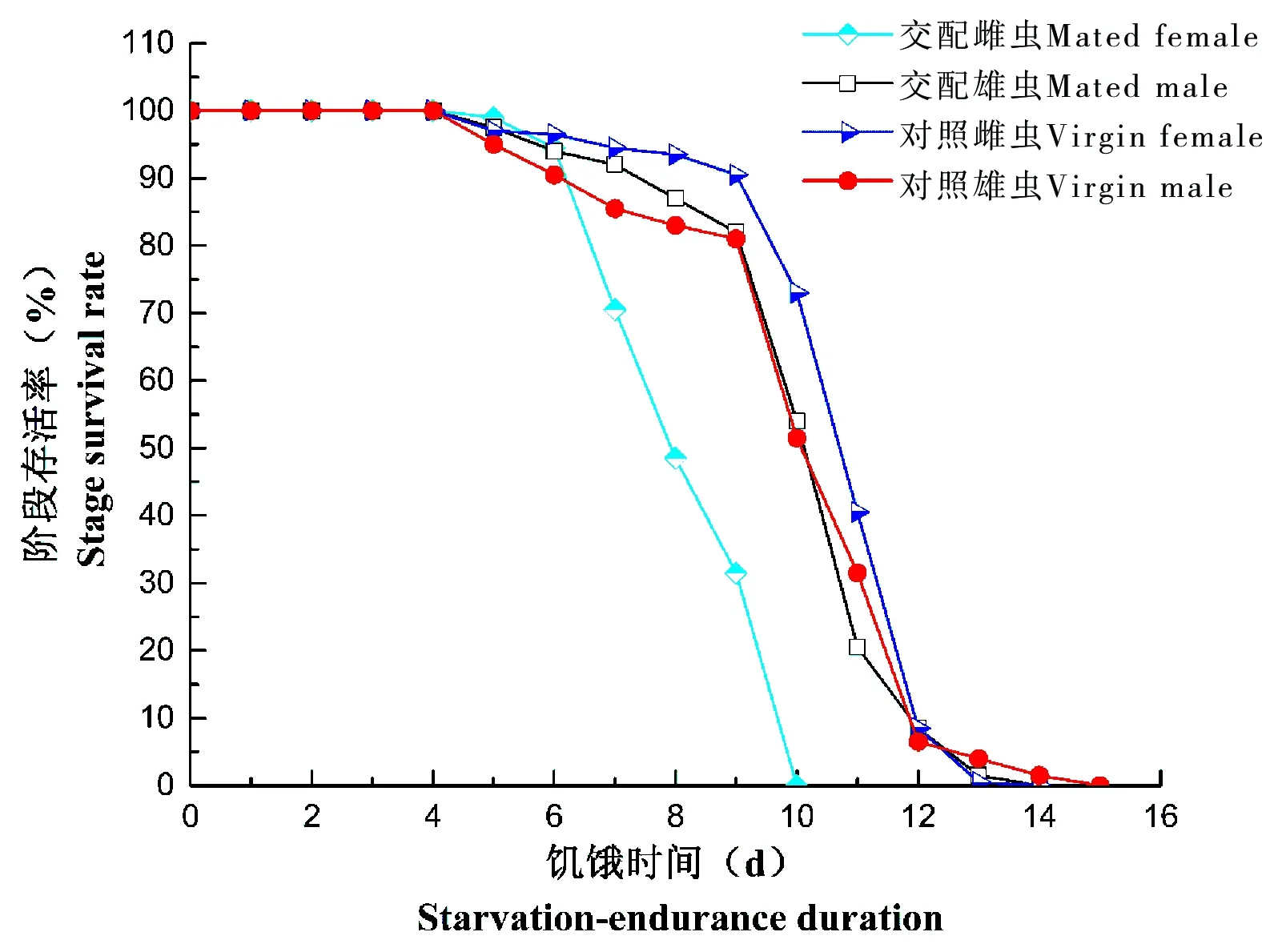
图1 广聚萤叶甲成虫饥饿条件下存活率Fig.1 The adults’ survival rate under starvation condition in Ophraella communa
用二次方程对不同处理的广聚萤叶甲成虫的阶段存活率进行拟合,得出;
YMF=93.56+9.91t-1.89t2(r2=0.975,P<0.0001)
YMM=96.68+5.15t-094t2(r2=0.938,P<0.0001)
YUF=92.45+7.66t-1.07t2(r2=0.927,P<0.0001)
YUM=101.22+1.81t-0.65t2(r2=0.933,P<0.0001)
以上各式中,t为广聚萤叶甲成虫饥饿时间(d)。各个处理的阶段存活率(y)和饥饿时间(t)之间均存在极显著的二次多项式相关性,可以用各自的拟合方程对存活率进行较准确的预测。
广聚萤叶甲雌虫交配后,在饥饿条件下,可以继续产卵,但是第一次产卵为交配后第5天,取食情况下,第一次往往在交配后第2天和第3天(图2)。交配雌虫在饥饿第7天和第10天,存活率急速下滑,雌虫的年龄-特征繁殖力(fx)表现为先增长后下降,最高峰为第6天,而年龄-阶段生殖能力(vxy)的曲线表示,雌虫的种群贡献在第5天和第6天最高(图2)。
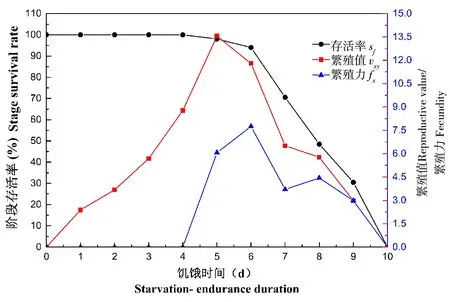
图2 广聚萤叶甲雌虫饥饿条件下繁殖力Fig.2 The female’ age-specific fecundity under starvation condition in Ophraella communa
2.2 交配对广聚萤叶甲成虫耐高温的影响
2.2.1饲喂豚草
经热处理后,在提供食物情况下,不同广聚萤叶甲成虫最短存活时间相同,但平均存活时间差异显著(F=36.53,df=7,P<0.0001)。交配显著影响广聚萤叶甲成虫耐热时间(P<0.0001),且最长存活时间显著高于未交配成虫。不考虑交配情况,雌雄虫耐热能力差异不显著(P=0.6213),交配和雌雄的交互作用差异显著(P=0.0003),其中交配雌虫在热处理后,存活时间显著高于其他处理,半致死时间和致死时间也显著长于其他处理。其中以对照雌虫的存活时间最短,显著低于其他处理。
在热处理条件下,提供豚草,各个处理的广聚萤叶甲成虫存活率曲线均成弓形,可以看出交配雌虫的存活率曲线下降缓慢,第7天死亡率最高(57.50%),交配雄虫在第5天至第7天死亡率较高(17.50%-36.67%)。而对照处理中,从第3天到第7天,成虫一直保持较高的死亡率。
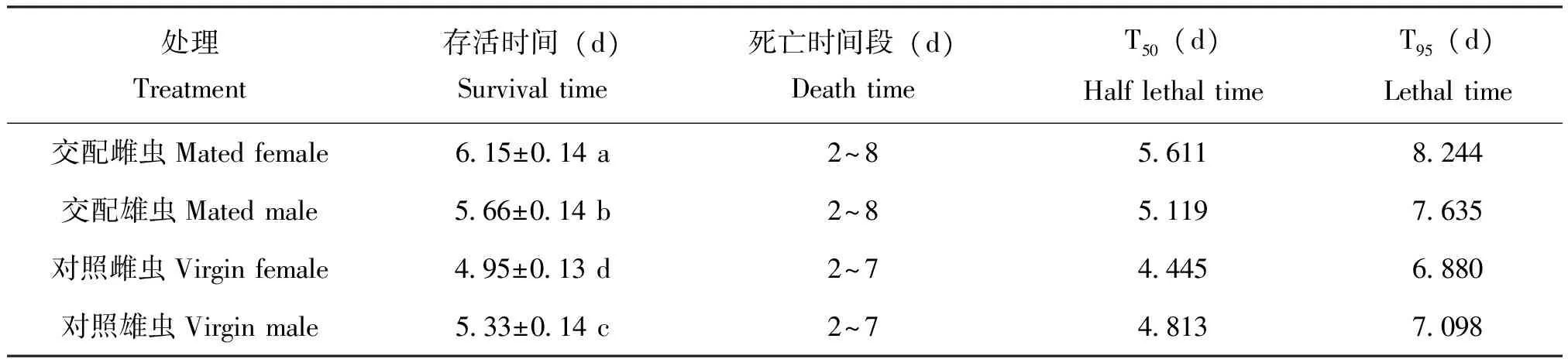
表2 广聚萤叶甲成虫耐热时间及热处理半致死时间(T50)和热处理致死时间(T95)(仅提供豚草)
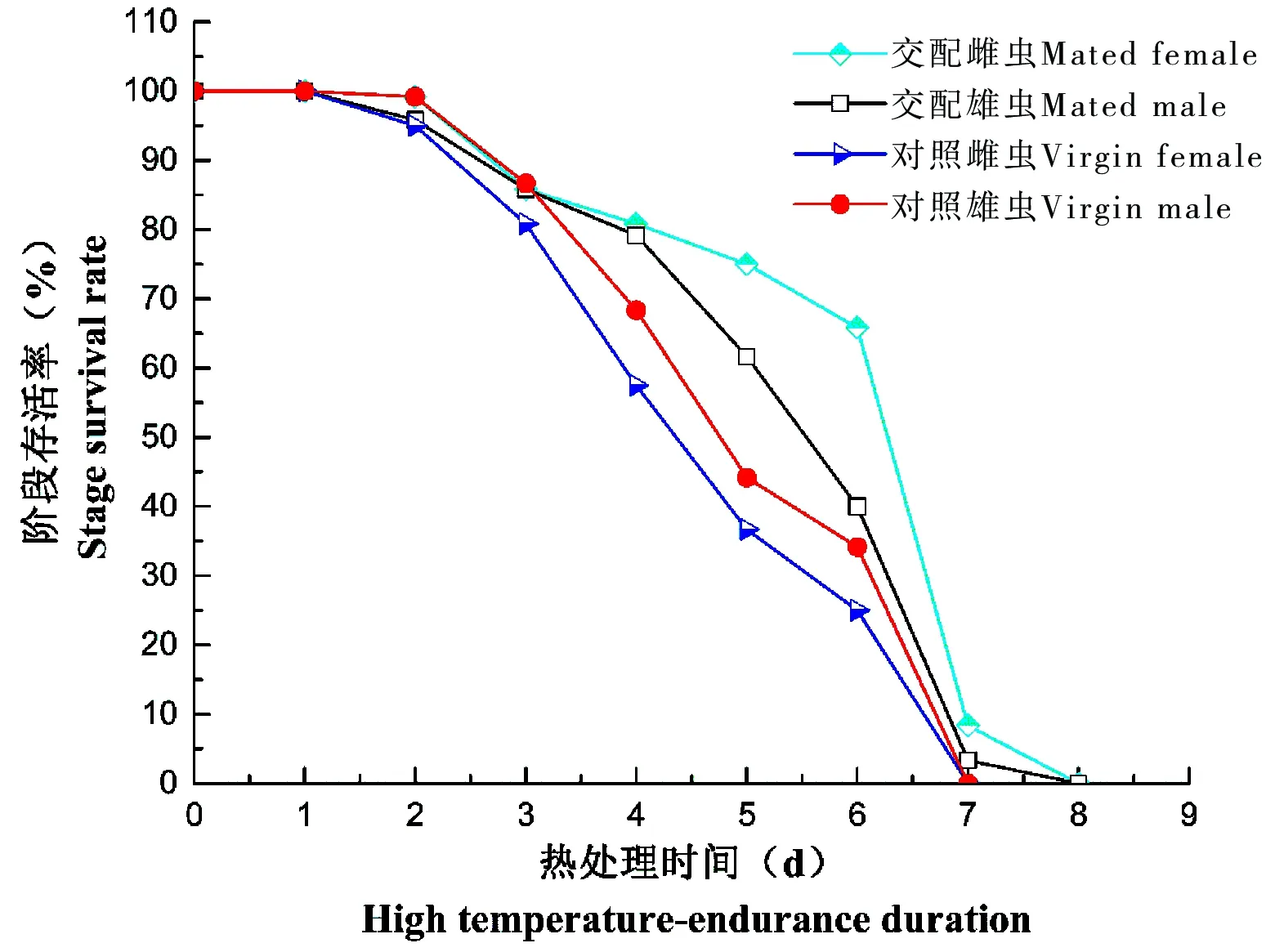
图3 广聚萤叶甲成虫在热处理条件下存活率(仅提供豚草)Fig.3 The adults’ survival rate under high temperature in Ophraella communa(Provide regweed)
2.2.2仅提供清水
在仅提供清水时,不同状态的广聚萤叶甲成虫耐热力差异显著(F=14.01,df=7,P<0.0001)。交配显著影响广聚萤叶甲成虫在热处理下的存活时间(P<0.0001),交配雌虫和雄虫的存活时间分别远远高于对照成虫,致死时间和半致死时间也远远长于对照成虫。其中交配雌虫的存活时间、存活周期、半致死时间、致死时间都最长。雌雄虫受热处理影响差异显著(P<0.0001),雌成虫在热处理下的存活时间远远长于雄成虫。
试验得出,广聚萤叶甲成虫在热处理后,交配成虫的存活率曲线下降比对照成虫下降缓慢,其中以对照雄虫存活率下降迅速,曲线呈凹形。对照雌虫总体生存时间较久,在热处理第7天存活率急剧下降,死亡率为35.83%,对照雌虫在热处理第4天至第7天,死亡率均超过22.5%,交配雄虫在热处理第3天至第5天,保持较高死亡率(20.83%、19.17%、31.67%),对照雌虫高死亡率出现在热处理第3天和第4天(30.83%、38.33%)。
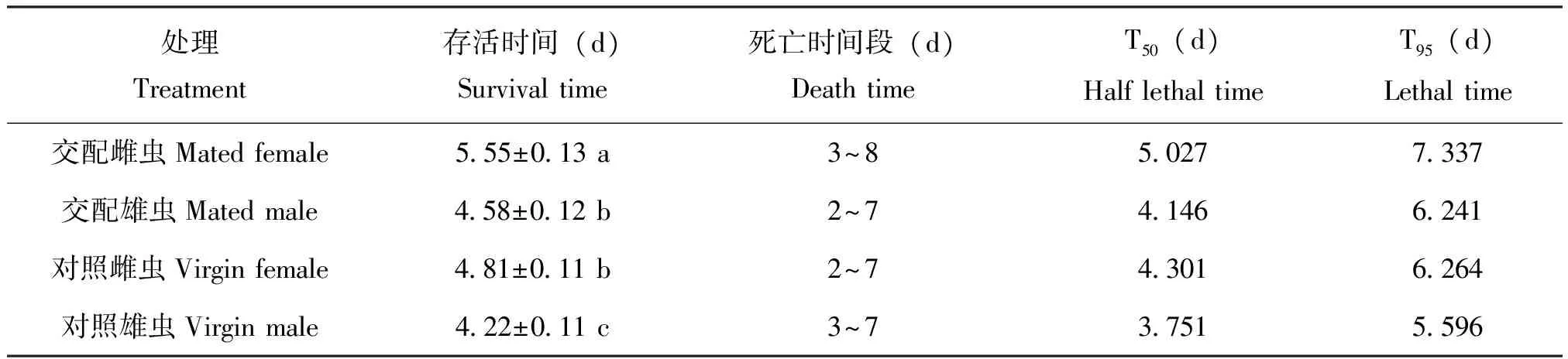
表3 广聚萤叶甲成虫耐热时间及热处理半致死时间(T50)和热处理致死时间(T95)(仅提供清水)
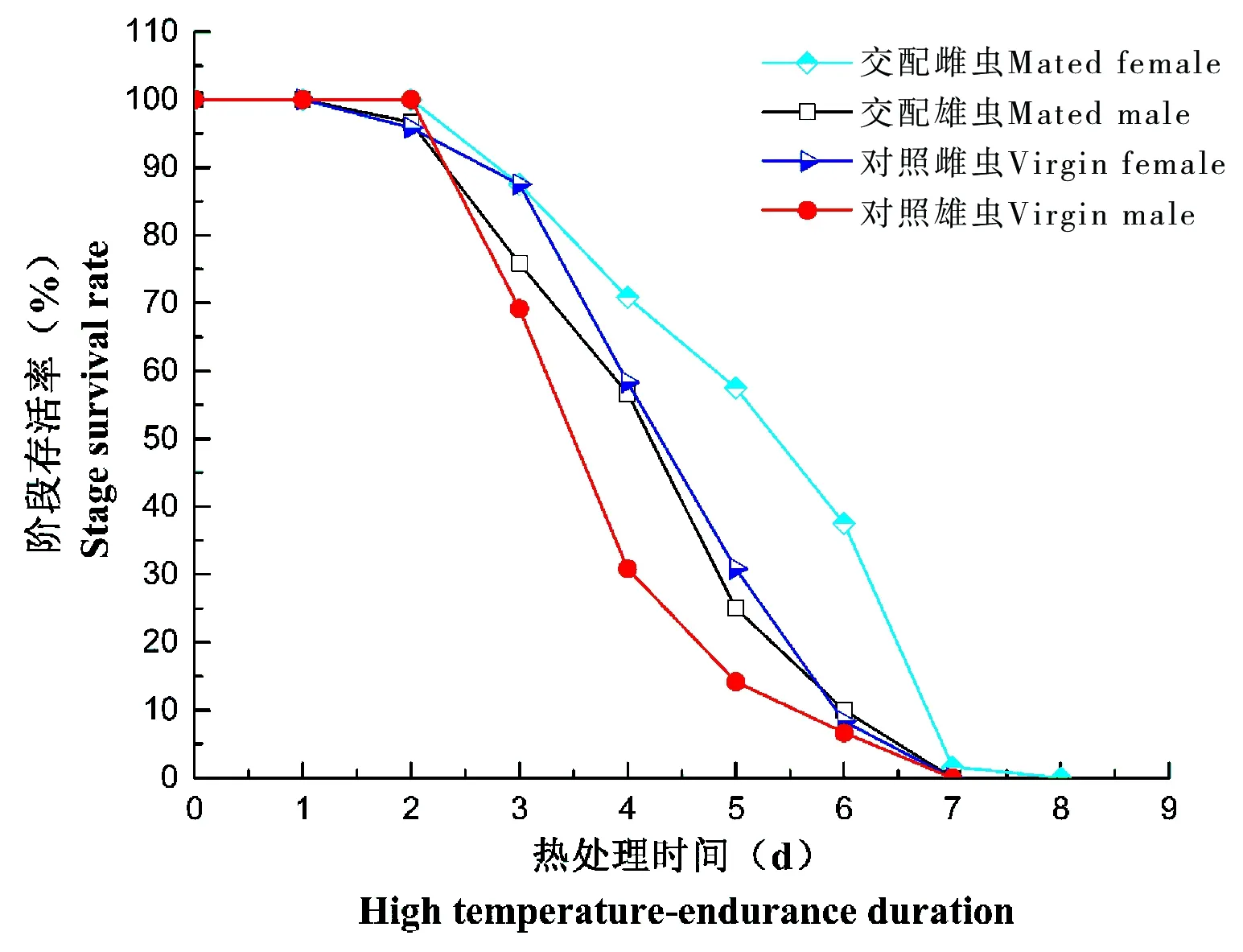
图4 广聚萤叶甲成虫在热处理条件下的存活率(仅提供清水)Fig.4 The adults’ survival rate under high temperature in Ophraella communa(Provide water)
3 结论与讨论
昆虫的耐饥饿和耐热能力关系到种群的生存能力,较强的耐饥饿和耐热能力可更好地适应恶劣环境。在长期进化适应自然界的过程中,昆虫不断提高对饥饿和高温的耐受性。不同状态下,昆虫对高温和饥饿的忍受程度存在差异,且昆虫会根据环境的变化进行行为学或生理学上的调整,例如烟草天蛾Manducasexta会减少蛹的大小(Kingsolveretal., 2009),果蝇Drosophilids中,胸、翅的大小和体重(Petavyetal., 2004; Czarnoleskietal., 2013)来适应温度的变化。对于天敌昆虫而言,其种群会因为食物短缺出现断代现象(陈洪凡等,2010),而前期研究发现广聚萤叶甲成虫会在低温下进行交配来降低的半致死低温(岳雷,2014)。
昆虫交配是一个消耗能量的过程,其交配后往往会出现昆虫抗逆性下降,寿命缩短的现象(Service, 1989; Avilaetal., 2011)。本研究发现,在不提供食物的情况下,交配一次后,广聚萤叶甲雄虫的寿命影响不大,但雌虫的寿命显著缩短。在高温状态下,无论取食与否,交配都会显著增加广聚萤叶甲雌雄成虫的耐热能力,尤其以雌虫表现更为明显(此时,雌虫不产卵)。同时,也观察到仅在饥饿状态下,广聚萤叶甲雌成虫交配后可持续产卵,但是寿命显著短于对照组,这一现象与泽兰实蝇Procecidocharesutilis和家蝇Muscadomestica的情况一致。交配后,泽兰实蝇雌虫的寿命会显著缩短(马沙等,2012)。在家蝇中,仅提供清水的情况下,交配也会显著降低雌虫的寿命(Hicksetal., 2004)。交配后雌虫寿命缩短被认为是繁殖成本消耗的效应(Renznick, 1985; Kirkwoodetal., 2005; Rush,etal., 2007),这也可能是广聚萤叶甲雌虫寿命缩短的主要原因。在昆虫两性交配过程中,雄虫会向雌虫体内转移精液蛋白(Seminal Fluid Proteins, SFPs)(Avilaetal., 2011)。对于某些物种的雌虫来说,交配的代价是昂贵的,因为雄虫精液中会含有毒物质,导致寿命缩短。但对于另一些物种来说,雌虫可以从交配中获益,引起寿命延长(Williametal., 2001)。在交配后一定时间内,雄虫可以自我补充精液蛋白(Sirotetal., 2009)。实验证明,广聚萤叶甲属于交配受益昆虫,雌雄成虫寿命均显著高于对照,交配提高了成虫抗逆性。
该叶甲成虫一生可多次交配,甚至一天也能交配很多次(孟玲和李保平,2006),仅一次交配对雄虫的能量消耗较少,而交配可以增强自身的活力,从而抗逆能力得以增强。至于给其多次交配后,广聚萤叶甲成虫的耐饥饿和耐热能力如何?还需要进一步进行验证与探讨。
References)
Angilletta MJJ. Thermal Adaptation: A Theoretical and Empirical Synthesis [M]. Oxford: Oxford University Press, 2009: 289.
Avila FW, Sirot LK, LaFlamme BA,etal. Insect seminal fluid proteins: Identification and function [J].AnnualReviewofEntomology, 2011, 56 (56): 21-40.
Bozinovic F, Bastías DA, Boher F,etal. The mean and variance of environmental temperature interact to determine physiological tolerance and fitness [J].PhysiologicalandBiochemicalZoology, 2011, 84 (6): 543.
Bruckner DJ, Lepossa A, Herpai Z. Ragweed allelopathay:Indirect interactions [J].Novenytermeles, 2001, 50 (2-3): 231-236.
Butler CD, Trumble JT. Predicting population dynamics of the parasitoidCotesiamarginiventris(Hymenoptera: Braconidae)resulting from novel interactions of temperature and selenium [J].BiocontrolScienceandTechnology, 2010, 20 (4): 391-406.
Charabidze D, Morvan G, Dupont D,etal. Forenseek: A simulation program of carrion-feeding insects development dedicated to forensic entomology [J].Annales-SocieteEntomologiquedeFrance, 2008, 44 (3): 385-392.
Clewis SB, Askew SD, Wilcut JW. Common ragweed interference in peanut [J].WeedScience, 2001, 49 (6): 768-772.
Colinet H, Hance T. Interspecific variation in the response to low temperature storage in different aphid parasitoids [J].AnnalsofAppliedBiology, 2010, 156 (1): 147-156.
Crockett LJ. Wildly Successful Plants: A Handbook of North American Weeds [M]. New York: Macmillan, 1977: 268.
Czarnoleski M, Cooper BS, Kierat J,etal. Flies developed small bodies and small cells in warm and in thermally fluctuating environments [J].JournalofExperimentalBiology, 2013, 216 (15): 2896-2901.
Dechamp C. Ragweed, a biological pollutant:Current and desirable legal implications in France and Europe [J].RevueFrancaiseDallergologieetDimmunologieClinique, 1999, 39 (4): 289-294.
Duan H, Chen B. Biological characters, encroaching habit and control strategy of common ragweed in Shanghai area [J].ActaAgriculturaeShanghai, 2000, 16 (3): 73-77.
EPPO. PQR database. Paris, France: European and Mediterranean Plant Protection Organization.2014, http://www.eppo.int/DATABASES/pqr/pqr.htm.
Gerber E, Schaffner U, Gassmann A,etal. Prospects for biological control ofAmbrosiaartemisiifoliain Europe: Learning from the past [J].WeedResearch, 2011, 51 (6): 559-573.
Hicks SK, Hagenbuch KL, Meffert LM. Variable costs of mating, longevity, and starvation resistance inMuscadomestica(Diptera: Muscidae)[J].EnvironmentalEntomology, 2004, 33 (3): 779-786.
Jolivet P, Santiago-Blay JA, Schmitt M. NewDevelopments in the Biology of Chrysomelidae [M]. The Hague, the Netherlands: SPB Academic Pub. 2004: 803.
Kingsolver JG, Ragland GJ, Diamond SE. Evolution in a constant environment:Thermal fluctuations and thermal sensitivity of laboratory and field populations ofManducasexta[J].Evolution, 2009, 63 (2): 537-541.
Kirkwood TB. Understanding the odd science of aging [J].Cell, 2005, 120: 437-447.
Kovalev OV. Spread of adventitious plants of thetribeAmbrosiain Eurasia and methods of biological control of weeds of the genusAmbrosiaL. (Ambrosieae, Asteraceae)[J].TrudyZoologicheskii,InstitutAkademiiNaukSSSR, 1989, 189: 7-23.
Marshall KE, Sinclair BJ. The impacts of repeated cold exposure on insects [J].JournalofExperimentalBiology, 2012, 215 (10): 1607-1613.
Nicolson SW, Chown S. Insect Physiological Ecology: Mechanisms and Patterns [M]. Oxford: Oxford University Press, 2004: 243.
Petavy G, David JR, Debat V,etal. Specific effects of cycling stressful temperatures upon phenotypic and genetic variability of size traits inDrosophilamelanogaster[J].EvolutionaryEcologyResearch, 2004, 6 (6): 873-890.
Renault D, Nedved O, Hervant F,etal. The importance of fluctuating thermal regimes for repairing chill injuries in the tropical beetleAlphitobiusdiaperinus(Coleoptera: Tenebrionidae)during exposure to low temperature [J].PhysiologicalEntomology, 2004, 29 (2): 139-145.
Renznick D. Costs of reproduction: An evaluation of the empirical evidence [J].Oikos, 1985, 44 (2): 257-267.
Rion S, Kawecki TJ. Evolutionary biology of starvation resistance:What we have learned fromDrosophila[J].JournalofEvolutionaryBiology, 2007, 20 (5): 1655-1664.
Rush B, Sandver S, Bruer J,etal. Mating increases starvation resistance and decreases oxidative stress resistance inDrosophilamelanogasterfemales [J].AgingCell, 2007, 6 (5): 723.
Service PM.The effect of mating status on lifespan, egg laying, and starvation resistance inDrosophilamelanogasterin relation to selection on longevity [J].JournalofInsectPhysiology, 1989, 35 (5): 447-452.
Sirot LK, Buehner NA, Fiumera AC,etal. Seminal fluid protein depletion and replenishment in the fruit fly,Drosophilamelanogaster: An ELISA-based method for tracking individual ejaculates [J].Behav.Ecol.Sociobiol., 2009, 63 (10): 1505-1530.
William EW, Jr RJ, Kelley KR,etal. Females receive a life-span benefit from male ejaculates in a field cricket [J].Evolution, 2001, 5 (5): 994-1001.
Chen HF, Huang SS, Zhang YZ,etal. Control efficacy ofTrichogrammajaponicumagainstChilosuppressalisandChilaraeaauricilia[J].TheJournalofAppliedEcology, 2010, 21 (3): 743-748. [陈洪凡, 黄寿山, 张玉烛, 等. 稻螟赤眼蜂对二化螟和台湾稻螟的控制潜能评价[J]. 应用生态学报, 2010, 21 (3): 743-748]
Ma S, Wu GX, Zhu JY,etal. Effects of supplying hydromel and mating behavior on life-span ofProcecidocharesutilisadults [J].ActaAgriculturaeJiangxi, 2012, 2: 83-85. [马沙, 吴国星, 朱家颖, 等. 补充蜂蜜水及交配行为对泽兰实蝇成虫寿命的影响[J]. 江西农业学报, 2012, 2: 83-85]
Meng L, Li BP. The mating and spawning behavior of theOphraellacommuna[J].AppliedEntomology, 2006, 4 (40): 11-13. [孟玲, 李保平. 广聚萤叶甲的交配和产卵行为[J]. 应用昆虫学报, 2006, 4 (40): 11-13]
Yue L, Zhou ZS, Liu ZB,etal. Effect of rapid cold hardening in different intensities on the physiological indices related to cold tolerance in adult ofOphraellacommuna(Coleoptera: Chrysomelidae)[J].ActaEntomologicaSinica, 2014: 631-638. [岳雷, 周忠实, 刘志邦, 等. 不同强度快速冷驯化对广聚萤叶甲成虫耐寒性生理指标的影响[J]. 昆虫学报, 2014: 631-638]
Zhou ZS, Guo JY, Wan FH,etal. Comprehensive assessment on control measures ofAmbrosiaartemisiifoliaL. [J].ChineseJournalofAppliedEcology, 2008, 19 (9): 1917-1924. [周忠实, 郭建英, 万方浩, 等. 豚草防治措施综合评价[J]. 应用生态学报, 2008, 19 (9): 1917-1924]
EffectofmatingonstarvationenduranceandthermaltoleranceofOphraellacommuna
ZHAO Chen-Chen1, MA Chao1, ZHANG Yan1,2, WAN Fang-Hao1, ZHOU Zhong-Shi1*
(1. Key Laboratory for Biology of Plant Disease and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China; 2. Guangxi Agricultural Environment and Agricultural Product Safety Key Laboratory Breeding Base, Guangxi 530005, China)
Insect mating is a time consuming process, so mating may have an adverse effect on its resistance to stresses.Ophraellacommunais a specific predator of ragweed, under normal circumstances, adults had stronger ability of bearing starvation and high temperature. In order to clear the effect of mating on the tolerance to environmental stresses inO.communaadults, the effects of mating on the resistance to starvation and thermal tolerance were studied. The result showed that in the absence of food, mating only once, the mating has strong effect on female’s longevity, but females (8.4 d±0.1 d)have significantly lower resistance to starvation than age-matched virgin females (10.8 d±0.1 d). At high temperature, regardless of feeding or not, mating will obviously enhance the heat resistance of male and female adults, especially for females (The survival time will be 6.15 d±0.14 d, 4.95 d±0.13 d for mated females and virgin females respectively when ragweed was provided; without food, the ratios will be 5.55 d±0.13 d, 4.81 d±0.11 d for mated females and virgin females respectively).Therefore,the starvation tolerance ofO.communafemales reduced after they copuloted,but copuation enhanced heat resistance of the beetle adults significant.
Mating; starvation endurance; thermal tolerance
国家自然科学基金项目(31672089)
赵晨晨,女,1991年生,硕士研究生,研究方向为昆虫生态学,E-mail:zhaochen06166@163.com
*通讯作者Author for correspondence, E-mail: zs.zh@126.com
Received: 2017-05-25; 接受日期Accepted: 2017-06-20
Q963;S433.5
:A
1674-0858(2017)04-905-07
赵晨晨,马超,张燕,等.交配对广聚萤叶甲成虫耐饥与耐热能力的影响[J].环境昆虫学报,2017,39(4):905-911.
