花前氮亏缺对水稻叶片氮代谢酶活性的影响
2017-09-03武辉向镜陈惠哲张玉屏张义凯姬广梅王亚梁史鸿志朱德峰
武辉 向镜 陈惠哲 张玉屏 张义凯 姬广梅 王亚梁 史鸿志 朱德峰
(中国水稻研究所/水稻生物学国家重点实验室,杭州310006;第一作者:wuhuiscience@163.com;*通讯作者:cnrice@qq.com)
花前氮亏缺对水稻叶片氮代谢酶活性的影响
武辉 向镜 陈惠哲 张玉屏 张义凯 姬广梅 王亚梁 史鸿志 朱德峰*
(中国水稻研究所/水稻生物学国家重点实验室,杭州310006;第一作者:wuhuiscience@163.com;*通讯作者:cnrice@qq.com)
以籼型杂交稻中浙优1号和籼粳型杂交稻甬优12为材料,试验分析了花前不同时期氮亏缺处理对水稻叶片氮代谢酶活性的影响。结果表明,花前氮亏缺导致植株上3叶氮浓度、NR、GS、GOGAT、GOT和GPT酶活性大幅下降,GDH活性显著增长,且各叶位对花前氮亏缺敏感度总体上表现为剑叶>倒2叶>倒3叶;破口期亏氮对籼型杂交稻中浙优1号上3叶氮同化酶活性的影响远小于减数分裂期处理,与之相比,籼粳杂交稻甬优12对破口期亏氮胁迫仍较敏感,表明中浙优1号植株氮代谢酶的亏氮敏感性由减数分裂期至破口期逐步下降,对土壤速效氮的需求同步降低,甬优12则对土壤供氮存在更高需求。
水稻;氮亏缺;花前;氮代谢;减数分裂期;破口期
我国水稻栽培普遍存在氮肥用量过高、施用方式不合理、养分损失严重等问题[1-2],导致氮肥利用率低[3]、地下水污染、江河湖泊水体富营养化、温室气体的增加等问题[4-6]。实现农业节氮减氮、提高氮肥利用率的同时做到稳产增产,已成为农业生产和环境和谐共存、可持续发展的必然趋势。研究表明,拔节至抽穗期是水稻吸氮高峰期[7-8],穗分化期特别是减数分裂期至始穗期对氮的响应非常敏感[9],此时供氮或亏氮,会对颖花退化、功能叶片氮代谢生理[10]、源库器官NSC积累[11]产生重要影响。硝酸还原酶(NR)、谷氨酰胺合成酶(GS)、谷氨酸合酶(GOGAT)、谷氨酸脱氢酶(GDH)、谷草转氨酶(GOT)和谷丙转氨酶(GPT)是作物氮素吸收、同化和循环过程中的关键酶,GS/GOGAT偶联形成的循环是高等植物氨同化的主要途径,在无机氮转化为有机氮的过程中起关键作用[10-13],全生育期或某时期缺氮均显著影响植株氮代谢酶活性[14-16],进而影响氮的转化和利用。本试验通过一种简易、有效的土壤速效氮淋洗减氮方法,并分别于减数分裂期和破口期进行淋洗处理,探讨土壤氮亏缺对籼型杂交稻和籼粳型杂交稻主功能叶氮代谢酶活性的影响,以期为阐明水稻氮素积累及转运特征、大田生产减氮技术应用提供研究基础。
1 材料与方法
1.1 试验材料
试验于2016年在中国水稻研究所富阳试验基地进行。供试材料选用籼粳型杂交稻品种甬优12和籼型杂交稻品种中浙优1号。试验采用自行设计的盆栽土壤速效氮淋洗结构(图1)进行水稻栽植,盆栽土壤理化性质:pH值5.80,有机质42.57 g/kg,全氮0.19%,碱解氮73.54 mg/kg,有效磷54.30 mg/kg,速效钾44.22 mg/kg。采用162穴大钵盘育秧,水稻机插专用育秧基质育秧,催芽种2粒/穴,移栽秧龄21~22 d,叶龄5.1~5.5,1丛/盆。全生育期施尿素3 g/盆,按基肥∶分蘖肥∶穗肥=4∶3∶3的比例施用。基肥于移栽前1 d施入,分蘖肥于移栽后7 d施入,穗肥于倒3~4叶施入。过磷酸钙(含P2O512%)3 g,随基肥一次性施入。氯化钾(含K2O 40%)2 g,按基肥和穗肥分2次等量施入。主要生育期情况见表1。
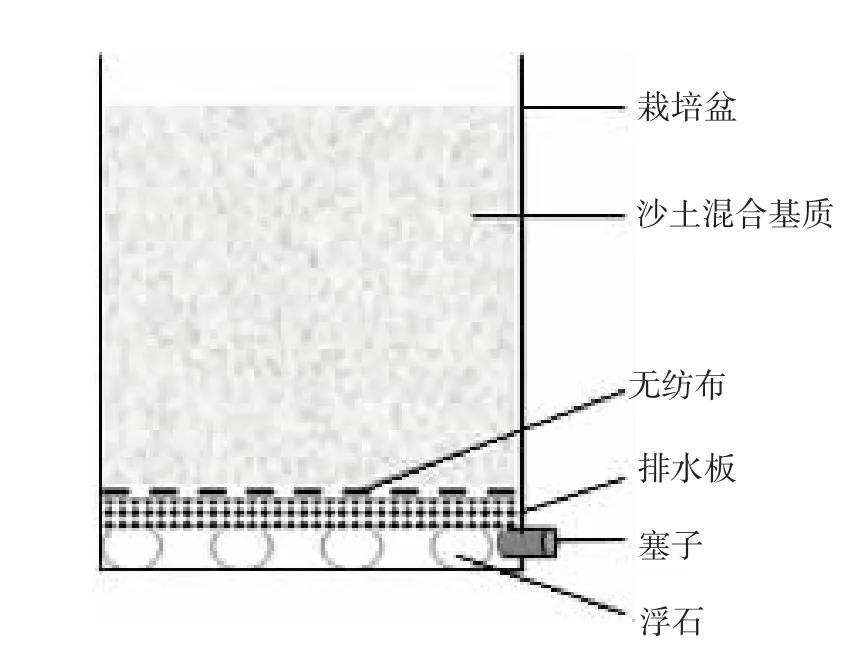
图1 盆栽土壤淋洗结构
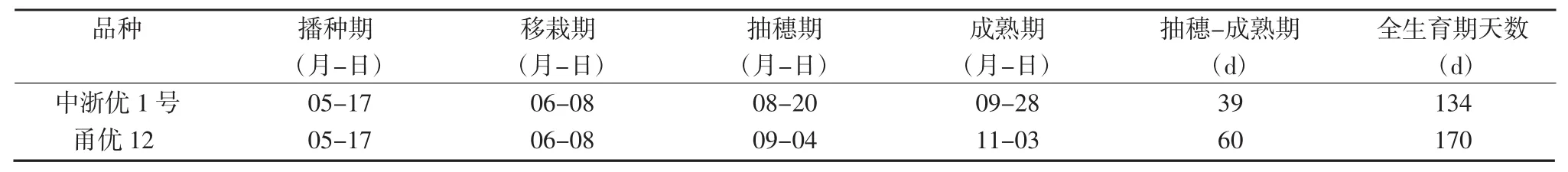
表1 参试品种主要生育期以及生育阶段天数
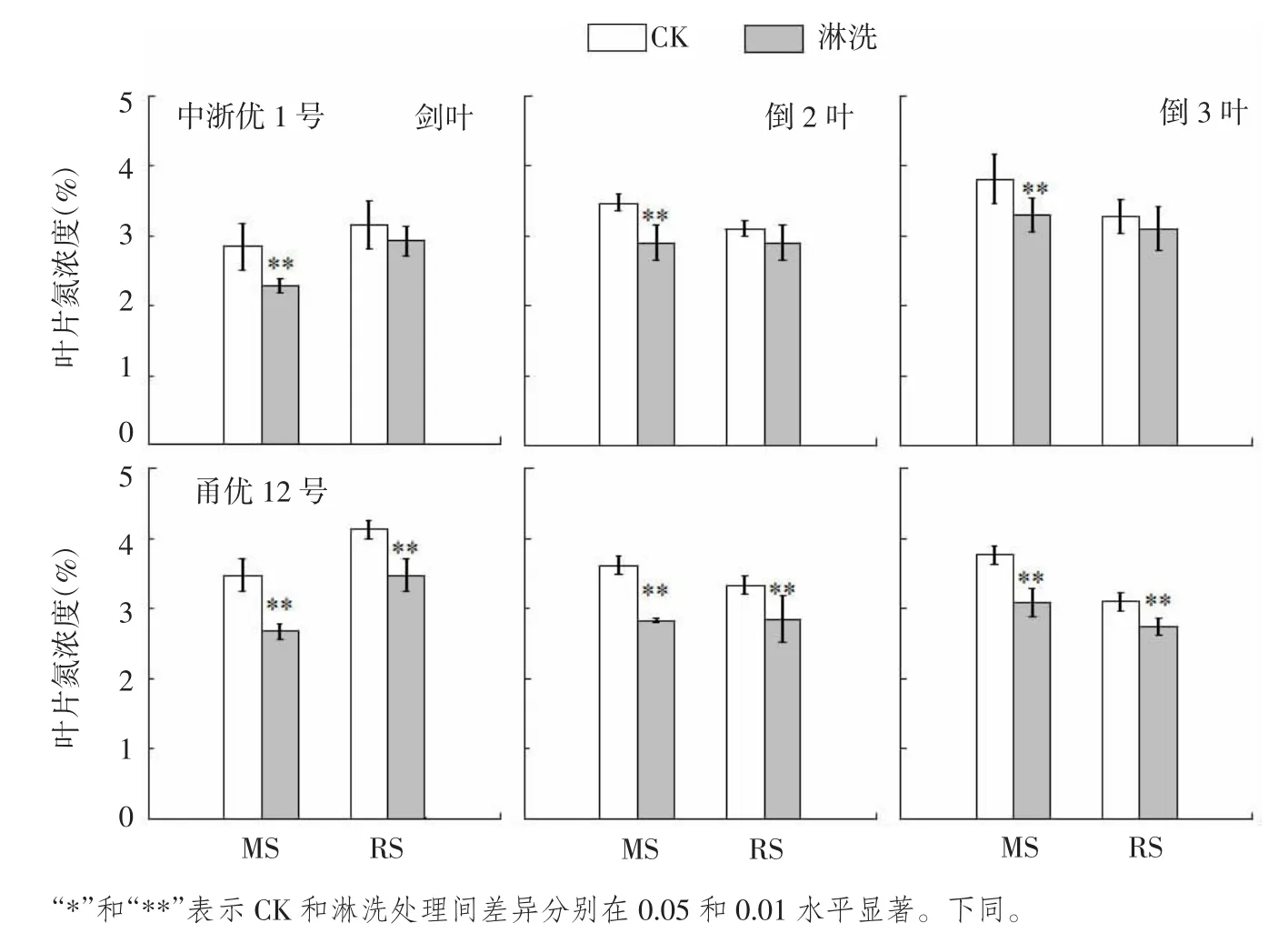
图2 水稻不同生长时期淋洗对上3叶氮浓度的影响
1.2 土壤速效氮淋洗结构
土壤淋洗结构如图1,栽培盆内从下到上依次为:浮石、带孔排水板(PVC材质)、无纺布(40 g/m2)、沙土混合基质。栽培盆内径为25.5 cm×20.0 cm×26.5 cm(长×宽×高),均装入沙土混合基质10±0.2 kg,土表距盆口约4 cm。栽培盆侧下方开有排水孔1个,并用橡胶塞堵塞。盆底部放置若干浮石,上置裁剪好的带孔排水板。排水板表面垫2层无纺布,防止淋洗过程中土壤随水淋失堵塞排水孔。沙土混合基质为过筛干土与粗沙按4∶1(w/w)混匀。
1.3 试验设计
试验分别于植株减数分裂期(MS,抽穗前14~15 d,剑叶抽出1/2)和破口期(RS,主茎抽穗前3 d),选择发育进程一致的植株进行淋洗处理,以不淋洗植株作为对照(CK)。淋洗前1 d,所有栽培盆浇水1 L确保土壤水饱和,第二天下午拔掉橡胶塞,开始第1次淋洗。12 h后收集周转箱内淋洗溶液,各盆加橡胶塞并浇水1 L,12 h后开始第2次淋洗,共淋洗3次。淋洗过程中花盆略微倾斜放置,便于淋洗液尽快排出。所有栽培盆置于遮雨棚下,淋洗均于夜间进行,减少日间高温引起的淋洗液蒸发和氨挥发的损耗。
1.4 测定项目与方法
1.4.1 植株上3叶氮浓度测定均于淋洗处理结束后,取主茎上3叶烘干,磨成细粉,用消煮法测定上3叶氮浓度(%,FOSS全自动定氮仪),各处理5次重复。
1.4.2 植株上3叶氮代谢酶活性测定分别于淋洗处理结束后,取主茎上3叶液氮速冻后,置于-80℃冰箱保存,用于硝酸还原酶(NR)、谷氨酰胺合成酶(GS)、谷氨酸合酶(GOGAT)、谷氨酸脱氢酶(GDH)、谷草转氨酶(GOT)和谷丙转氨酶(GPT)的测定。所取叶片组织鲜质量为0.100 g,使用全自动样品快速研磨仪(Tissuelyser-48,上海净信)液氮条件下磨粉,采用试剂盒(苏州科铭)提取组织样品的原液,经试剂盒待测液用酶标仪(Tecan Infinite M200 PRO)测定各指标。各指标均测4次。
1.5 数据统计与分析
数据统计分析采用Microsoft Excel 2016和SPSS 23.0软件结合进行;Duncan新复极差法进行差异显著性检验;Origin 8.5软件制图,图中误差线均为标准误差。
2 结果与分析
2.1 植株上3叶氮浓度
土壤中90%以上的氮素以有机形态存贮,只有经过矿化转变成无机态的铵态氮和硝态氮,才可被作物吸收利用[17]。笔者之前试验采用淋洗结构+沙土混合基质,分别于水稻减数分裂期和破口期进行淋洗处理,土壤硝态氮和铵态氮含量降幅分别达42.1%~59.3%和35.0%~43.9%,可实现预期的土壤减氮效果,短期内对土壤速效氮供应能力形成有效抑制(论文待审)。与此同时,参试品种上3叶氮浓度均同步大幅下降(图2),意味着淋洗处理对水稻造成短期氮亏缺。
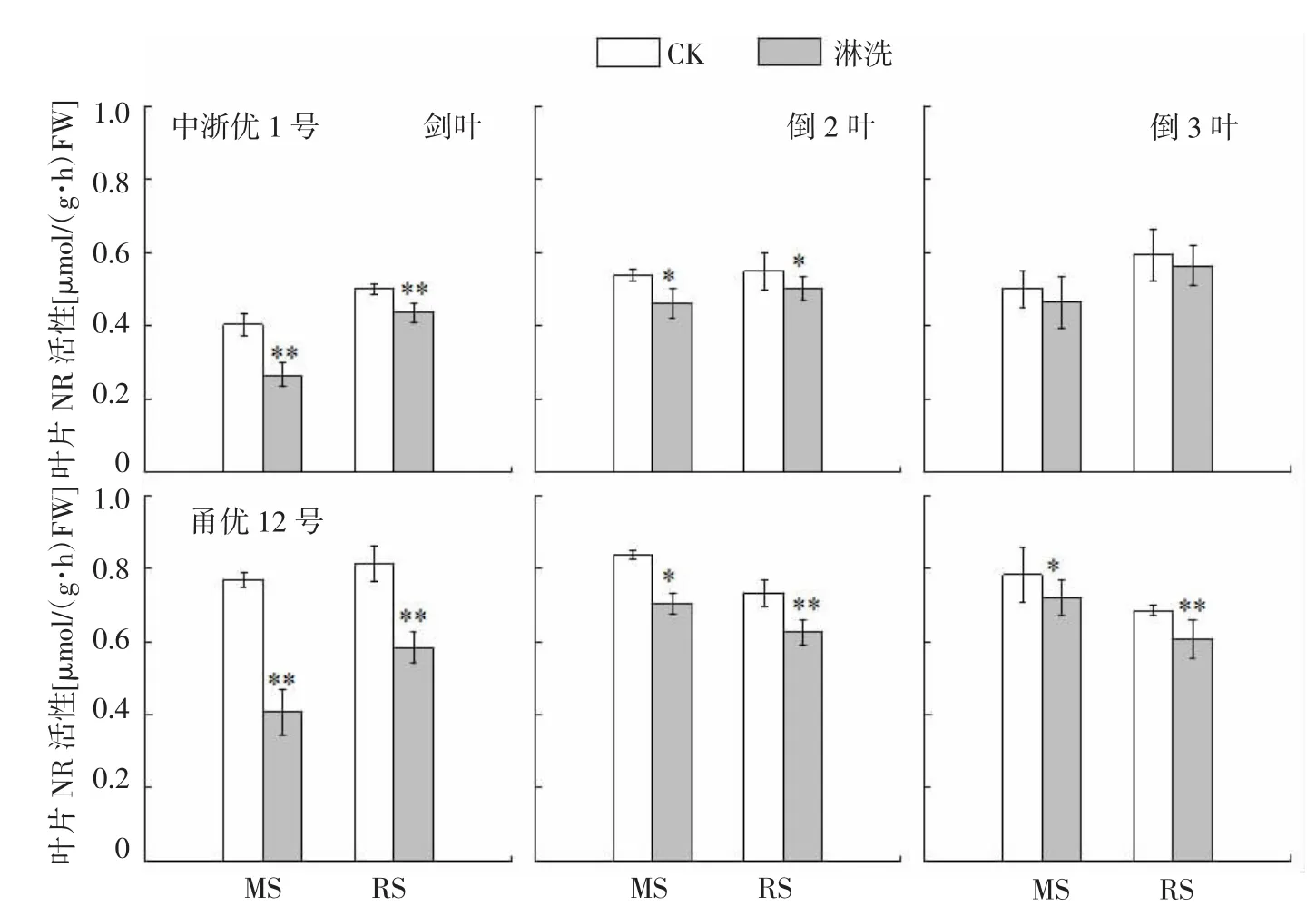
图3 水稻氮亏缺对上3叶NR活性的影响
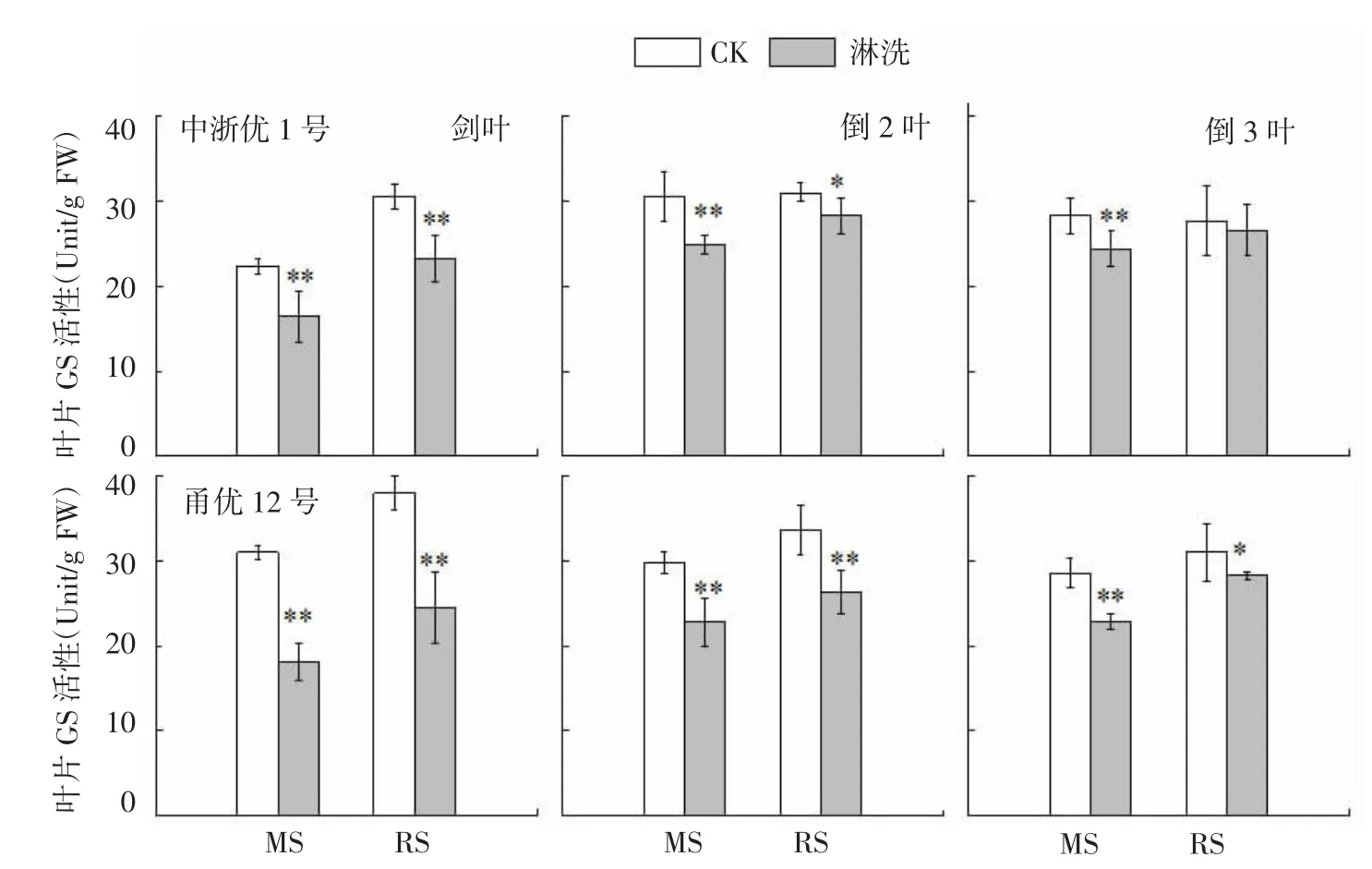
图4 水稻氮亏缺对上3叶GS活性的影响
与CK相比,减数分裂期(MS)淋洗处理导致中浙优1号叶片氮浓度显著下降,各叶位降低幅度为13.2%~19.5%,破口期(RS)淋洗处理下中浙优1号各叶位氮浓度仅下降5.2%~7.1%,与CK差异未达显著水平。淋洗处理导致甬优12叶片氮浓度显著下降,降幅分别达18.2%~23.3%(MS)和11.3%~15.8%(RS)。总体上,各叶位氮浓度降幅排序为剑叶>倒2叶>倒3叶,品种间则表现为甬优12>中浙优1号。与破口期淋洗处理相比,水稻减数分裂期淋洗处理剑叶仅抽出一半,倒2叶和倒3叶氮积累时间亦比破口期少11~14 d,植株上3叶氮浓度对淋洗处理的响应远大于破口期。
2.2 硝酸还原酶(NR)
NR是催化硝态氮转化为铵态氮过程中的限速酶,不同时期土壤速效氮供应不足则导致水稻叶片NR活性大幅下降。由图3可知,与CK相比,中浙优1号上3叶NR活性在减数分裂期(MS)和破口期(RS)亏氮处理中分别下降7.3%~33.9%和5.1%~13.0%,特别是剑叶和倒2叶NR活性与CK呈显著或极显著差异。花前两时期氮亏缺均导致甬优12叶片NR活性显著下降,降幅分别达8.1%~46.9%(MS)和11.2%~28.5%(RS)。2个品种各叶位NR活性降幅排序为剑叶>倒2叶>倒3叶,品种间NR活性水平及对氮亏缺敏感性均表现为甬优12>中浙优1号。
2.3 谷氨酰胺合成酶(GS)
GS将植物吸收的无机态铵转为有机态的谷氨酰胺,是处于氮代谢中心的多功能酶,其活性对花前土壤氮亏缺的响应与NR变化相似(图4)。与CK相比,中浙优1号上3叶GS活性在减数分裂期(MS)和破口期(RS)亏氮处理中分别下降13.9%~26.3%和3.9%~23.8%,且均与CK呈显著差异(除破口期处理倒3叶)。花前两时期氮亏缺均导致甬优12叶片GS活性显著下降,降幅分别达19.9%~41.6%(MS)和8.6%~35.5%(RS)。与NR类似,两品种各叶位GS活性降幅排序为剑叶>倒2叶>倒3叶,品种间GS对氮亏缺敏感性表现为甬优12>中浙优1号。
2.4 谷氨酸合酶(GOGAT)
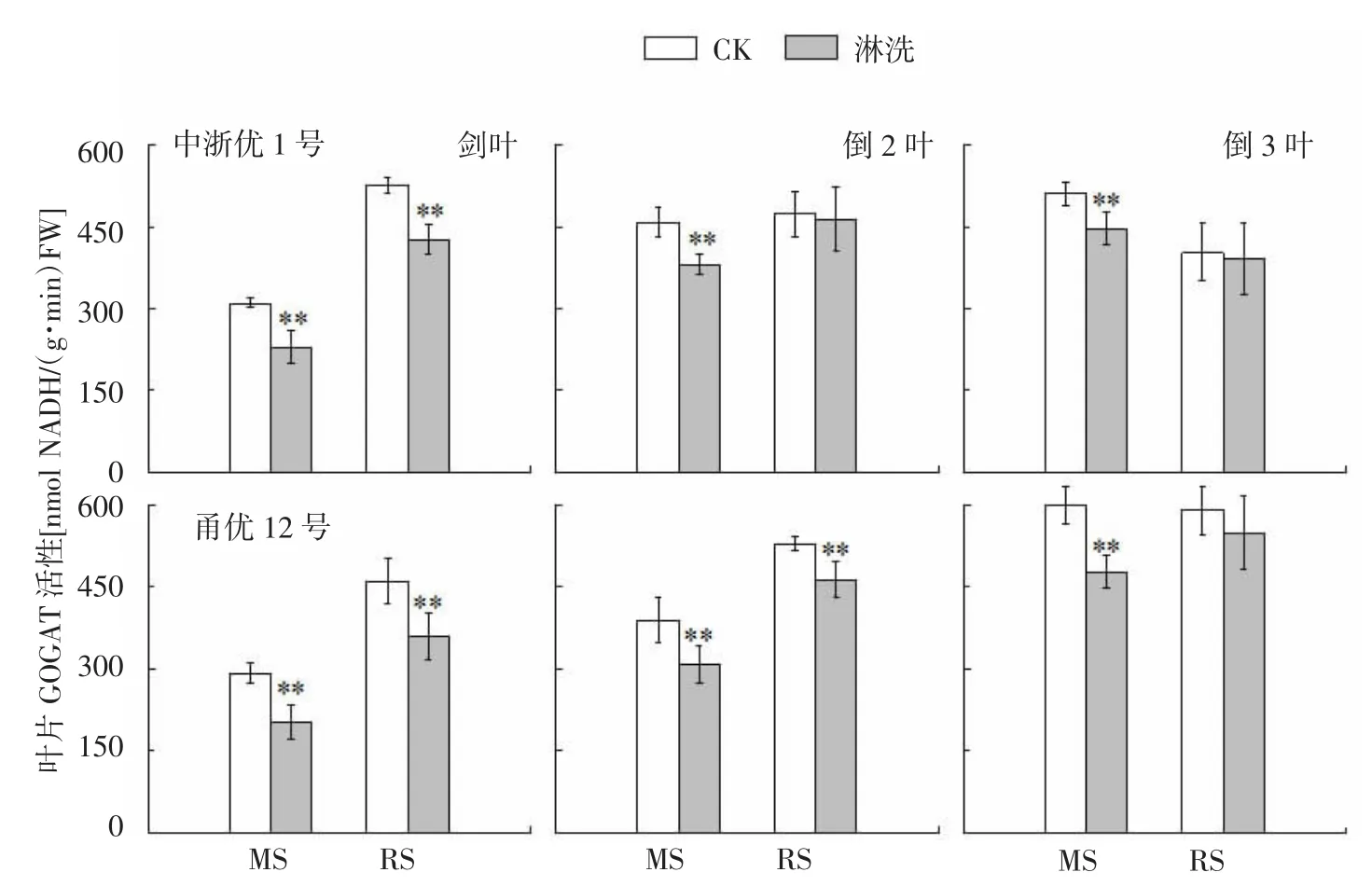
图5 水稻氮亏缺对上3叶GOGAT活性的影响
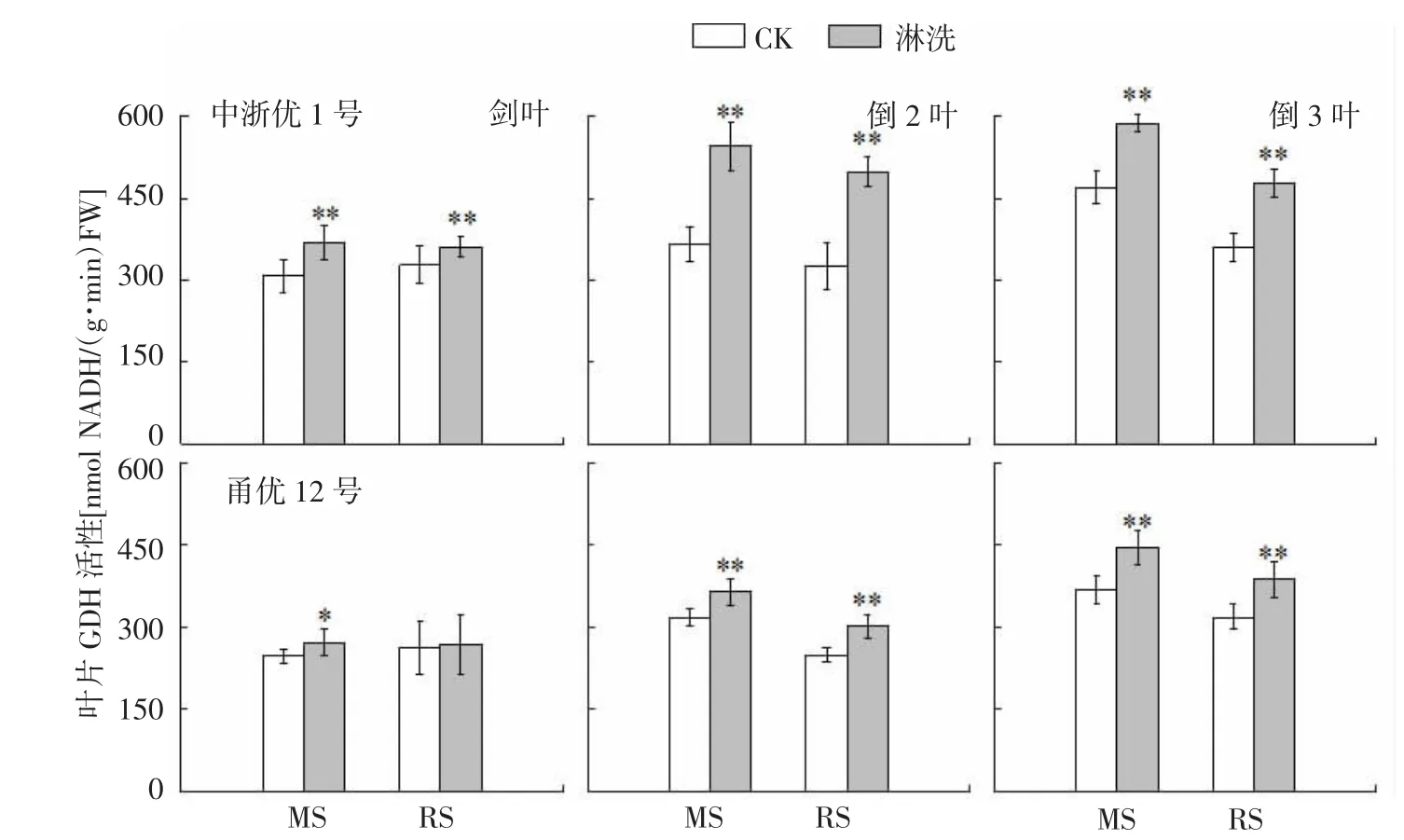
图6 水稻氮亏缺对上3叶GDH活性的影响
由图5可知,减数分裂期氮亏缺条件下,中浙优1号上3叶GOGAT活性较CK下降12.5%~26.1%,差异达显著水平;破口期亏氮导致剑叶GOGAT活性下降18.9%,对倒2叶和倒3叶则几无影响。花前两时期氮亏缺导致甬优12叶片GOGAT活性分别下降20.3%~30.6%(MS)和6.9%~22.0%(RS),除破口期倒3叶外均与CK差异显著。MS处理正值剑叶抽出阶段,参试的2个品种各叶位GOGAT活性高低排序均为剑叶<倒2叶<倒3叶,RS时期主功能叶全部展平,中浙优1号各叶位GOGAT活性高低排序为剑叶>倒2叶>倒3叶,而甬优12上3叶仍表现为剑叶<倒2叶<倒3叶。参试的2个品种各叶位对氮亏缺敏感性均表现为剑叶>倒2叶>倒3叶。
2.5 谷氨酸脱氢酶(GDH)
GDH在铵同化中起补充GS/GOGAT循环的作用,当作物遇到环境胁迫和作物发育后期时发挥着重要作用[18]。由图6可知,中浙优1号叶片GDH活性在减数分裂期和破口期亏氮处理中较CK分别增长19.7%~49.7%(MS) 和 10.0%~53.0%(RS),上3叶增幅排序为倒2叶>倒3叶>剑叶,甬优12同比增长9.9%~21.0%(MS)和2.7%~21.7%(RS),上3叶增幅排序为倒3叶>倒2叶>剑叶。
2.6 谷草转氨酶(GOT)
GOT、GPT是作物体内2种主要的转氨酶,涉及合成限制性必需氨基酸,并与光合作用、呼吸代谢间联系紧密,是反映叶片氨转运和贮藏、氮同化及氮代谢与碳代谢的协调方面的主要指标之一。由图7可见,花前不同时期亏氮处理导致参试2个品种叶片GOT活性显著下降,中浙优1号上3叶降幅达16.0%~29.8%(MS)和15.9%~22.5%(RS),甬优12同比降低了13.7%~19.1%(MS)和10.0%~18.0%(RS),各叶位对氮亏缺敏感性均表现为倒2叶>倒3叶>剑叶。
2.7 谷丙转氨酶(GPT)
花前不同时期亏氮处理中,参试2个品种叶片GPT活性均呈下降趋势,但与GOT存在明显差异。减数分裂期亏氮处理导致参试2个品种上3叶GPT活性分别下降7.9%~20.1%(中浙优1号)和8.8%~19.1%(甬优12),各叶位降幅排序为倒2叶>倒3叶>剑叶;破口期则同比降低7.1%~31.3%(中浙优1号)和2.9%~24.3%(甬优12),各叶位降幅排序为剑叶>倒2叶>倒3叶。
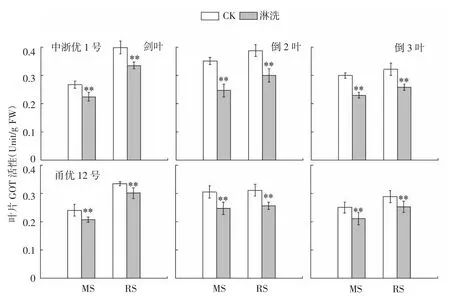
图7 水稻氮亏缺对上3叶GOT活性的影响
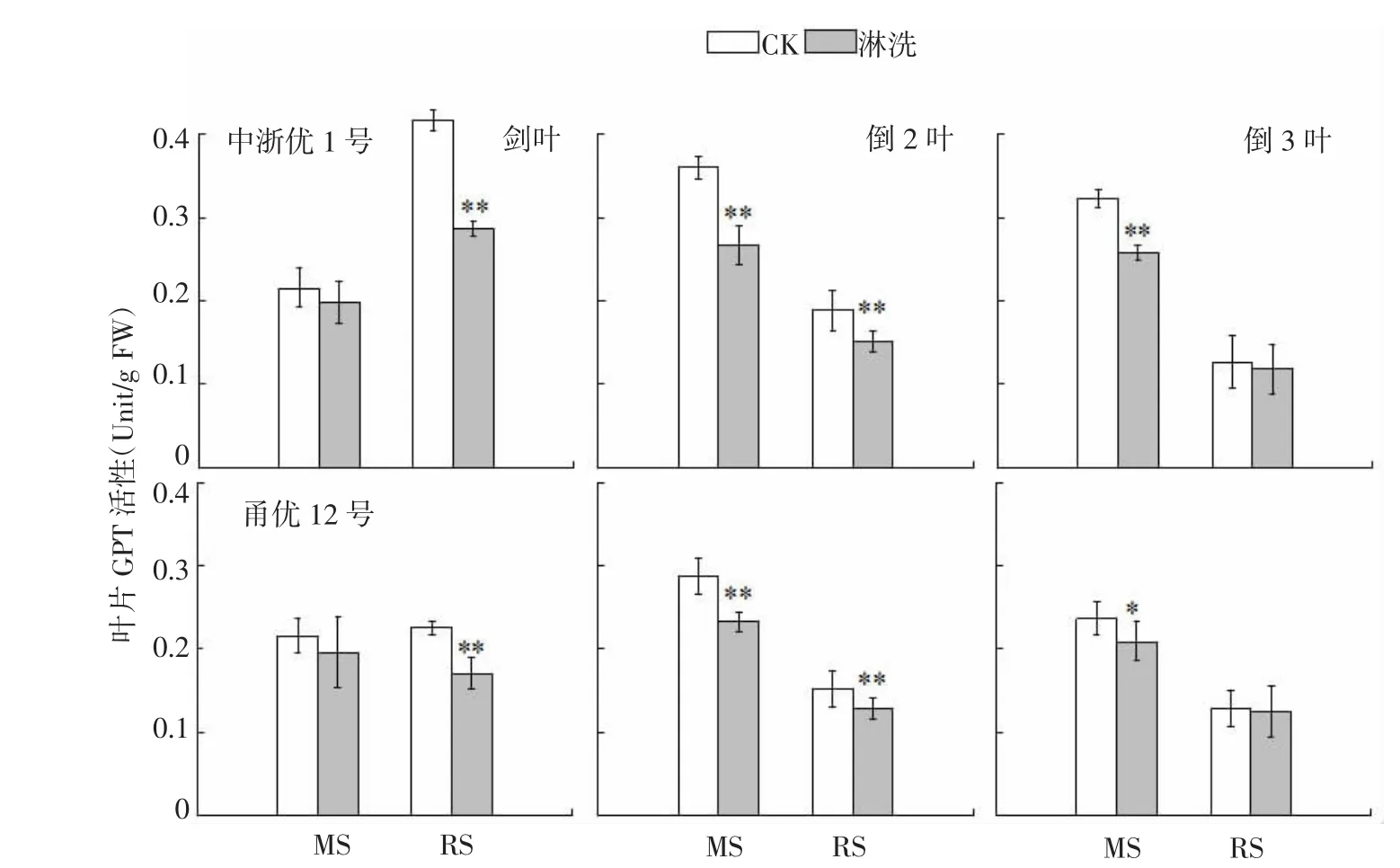
图8 水稻氮亏缺对上3叶GPT活性的影响
3 讨论与结论
本研究发现,水稻花前短期氮亏缺后,不同叶位叶片NR、GS和GOGAT酶活性均显著下降,表明植株短期内对无机氮的吸收同化受到限制,导致叶片氮浓度降低,且各叶位对土壤短期氮亏缺敏感度表现为剑叶>倒2叶>倒3叶,这与前人研究结果相反[19]。其原因在于长期氮饥饿引起下位叶含氮物质持续分解并向上转运[10],而短期亏氮则仅引起氮同化受限,且新叶受限最为严重,因此减数分裂期氮亏缺叶片氮浓度和氮同化酶活性下降幅度远大于破口期氮亏缺。氨通过GS/GOGAT循环后,进一步通过GOT和GPT催化转入到其他氨基酸中,两者均能在营养生长期消耗谷氨酸提供氮源,在衰老期产生谷氨酸供应植物[20],其酶活性的显著下降表明氨转化途径受阻,植株生长所需氨基酸来源不足,而GDH的显著增高意味着叶片储存蛋白快速降解[21],以补偿氮亏缺引起的氨基酸供应不足。
此外,不同类型水稻叶片的氮同化酶活性对花前氮亏缺的响应存在明显差异。破口期亏氮处理对籼型杂交稻中浙优1号上3叶氮浓度、NR、GS和GOGAT活性的影响远小于减数分裂期处理,表明中浙优1号植株氮代谢酶对亏氮敏感性由减数分裂期至破口期逐步下降,对土壤速效氮的需求同步降低。与之相比,两时期减氮处理均导致籼粳杂交稻甬优12上3叶氮浓度和氮同化酶活性显著下降,这意味着此类型水稻在开花前后对亏氮敏感性和土壤速效氮需求较籼型杂交稻更高。同时,花前植株GDH酶活性和氮亏缺引起的增幅均表现为中浙优1号远大于甬优12,说明中浙优1号主要依赖快速降解存储蛋白质来应对短期氮亏缺,而甬优12则更多通过氮吸收、同化和氨转化来进行氨基酸循环。
[1] Peng S B,Buresh R J,Huang J L,et al.Strategies for overcoming low agronomic nitrogen use efficiency in irrigated rice systems in China [J].Field Crop Res,2006,96(1):37-47.
[2] Guo J H,Liu X J,Zhang Y,et al.Significant Acidification in Major Chinese Croplands[J].Science,2010,327(5968):1 008-1 010.
[3]张福锁,王激清,张卫峰,等.中国主要粮食作物肥料利用率现状与提高途径[J].土壤学报,2008,45(5):915-924.
[4]曹仁林,贾晓葵.我国集约化农业中氮污染问题及防治对策[J].土壤肥料,2001(3):3-6.
[5] Vitousek P M,Naylor R,Crews T,et al.Nutrient imbalances in agricultural development[J].Science,2009,324(5934):1 519- 1 520.
[6] Gilbert N.The disappearing nutrient[J].Nature,2009,461(7265): 716-718.
[7]俞巧钢,叶静,杨梢娜,等.不同施氮量对单季稻养分吸收及氨挥发损失的影响[J].中国水稻科学,2012,26(4):487-494.
[8]王秀芹,张洪程,黄银忠,等.施氮量对不同类型水稻品种吸氮特性及氮肥利用率的影响 [J].上海交通大学学报:农业科学版,2003,21(4):325-330.
[9]陈玉泉,王延颐.水稻群体叶色的光谱诊断及其在高产栽培中的应用基础[J].江苏农业学报,1995(2):6-10.
[10]赵全志,陈静蕊,刘辉,等.水稻氮素同化关键酶活性与叶色变化的关系[J].中国农业科学,2008,41(9):2 607-2 616.
[11]李刚华,王惠芝,王绍华,等.穗肥对水稻穗分化期碳氮代谢及颖花数的影响[J].南京农业大学学报,2010,33(1):1-5.
[12]孙永健,孙园园,李旭毅,等.水氮互作下水稻氮代谢关键酶活性与氮素利用的关系[J].作物学报,2009.35(11):2 055-2 063.
[13]付景,王志琴,袁莉民,等.施氮量对超级稻产量和一些生理性状的影响[J].中国水稻科学,2014,28(4):391-400.
[14]曾建敏,崔克辉,黄见良,等.水稻生理生化特性对氮肥的反应及与氮利用效率的关系[J].作物学报,2007,33(7):1 168-1 176.
[15]罗凤,卢永恩,杨猛,等.氮胁迫对水稻营养生长期氮代谢及相关基因表达量的影响[J].华中农业大学学报,2012,31(1):16-22.
[16]易媛,董召娣,张明伟,等.减氮对半冬性中筋小麦产量、NUE及氮代谢关键酶活性的影响[J].核农学报,2015,29(2):365-374.
[17]张玉玲,陈温福,虞娜,等.北方地区滨海盐渍土型稻田土壤供氮能力的研究[J].土壤通报,2011,42(5):1 143-1 147.
[18]Kwinta J,Bielawski W.Glutamate dehydrogenase in higher plants[J]. Acta Physiologiae Plantarum,1998,20(4):453-463.
[19]Wang S,Zhu Y,Jiang H,et al.Positional differences in nitrogen and sugar concentrations of upper leaves relate to plant N status in rice under different N rates[J].Field Crop Res,2006,96(2-3):224-234.
[20]路兴花,吴良欢,庞林江.节水栽培水稻某些氮代谢生理特性研究[J].植物营养与肥料学报,2009,15(4):737-743.
[21]汪和廷,董慧,齐龙昌,等.种植方式及施氮量对水稻灌浆初期氮代谢关键酶活性和产量性状的影响 [J].中国生态农业学报,2015,23(9):1 210-1 214.
Effects of Pre-anthesis Nitrogen Deficiency on Nitrogen Metabolism Enzyme Activities of Rice Leaves
WU Hui,XIANG Jing,CHEN Huizhe,ZHANG Yuping,ZHANG Yikai,WANG Yaliang,JI Guangmei,SHI Hongzhi,ZHU Defeng*
(State Key Laboratory of Rice Biology/China National Rice Research Institute,Hangzhou 310006,China;1st author:wuhuiscience@163.com;*Corresponding author:cnrice@qq.com)
Effects of pre-anthesis nitrogen deficiency at different growth stages on nitrogen metabolism enzyme activities in main functional leaves were studied,using indica-japanica hybirid rice variety Yongyou 12,and indica hybrid rice variety Zhongzheyou 1 as materials.The results showed that,affected by nitrogen deficiency at meiosis stage and rupturing stage,nitrogen concentration,NR, GS,GOGAT,GOT and GPT enzyme activities of top-three leaves were dramatically decreased,but GDH activity was increased significantly.In general,the sensitivity of enzyme activities of leaves at different leaf positions to nitrogen deficiency were enhanced from bottom to top.Furthermore,the effects of nitrogen deficiency at rupturing stage on nitrogen assimilation enzyme activities of indica hybrid rice Zhongzheyou1 was much less than that of meiosis stage.However,indica-japanica hybirid rice Yongyou 12 was still sensitive to nitrogen deficiency during rupturing stage.This indicated that the nitrogen deficiency sensitivity of nitrogen metabolism enzymes in Zhongzheyou 1 plants decreased gradually from meiosis stage to rupturing stage,and the demand for soil available nitrogen was decreased synchronously,while Yongyou 12 had higher demand for soil nitrogen supply.
rice;nitrogen deficiency;pre-anthesis;nitrogen metabolism;meiosis stage;rupturing stage
S511.04
A
1006-8082(2017)04-0014-06
2017-06-21
粮丰工程项目(2016YFD0300502);国家自然科学基金青年科学基金项目(31501272);2015年浙江省“三农六方”项目
