菲律宾蛤仔选育家系间杂交的Kung育种值及配合力分析
2017-07-24孙谦,段强,桑士田等
菲律宾蛤仔选育家系间杂交的Kung育种值及配合力分析
孙谦,段强,桑士田,霍忠明,闫喜武
(大连海洋大学水产与生命学院,辽宁省贝类良种繁育工程技术研究中心实验室,辽宁大连116023)
为评估菲律宾蛤仔Ruditapes philippinarum家系间杂交的遗传效应,以生长和抗逆性状优良的壳面具有斑块花纹的蛤仔家系子一代(C31、C32)及壳面背景颜色为橙色的蛤仔家系子三代(Or)作为基础群体,采用单对单双列杂交方法构建选育家系,对建立的蛤仔家系进行Kung育种值、配合力和杂种优势分析。结果表明:15日龄时,C32(♀)×Or(♂)家系Kung育种值最大,30~90日龄时,C31(♀)×C32(♂)家系Kung育种值最大;6日龄时,各杂交家系一般配合力有极显著性差异(P<0.01),特殊配合力无显著性差异(P>0.05);9~90日龄时,除60日龄杂交家系特殊配合力无显著性差异外(P>0.05),各测量日龄杂交家系的一般配合力、特殊配合力和反交效应均有显著或极显著性差异(P<0.05或P<0.01);30日龄后,C31(♀)×C32(♂)杂交家系子代杂种优势最为明显。研究表明,以斑块花纹C31(♀)与斑块花纹C32(♂)为亲本的杂交家系表现出较好的Kung育种值、配合力和杂种优势,是较好的杂交亲本来源。
菲律宾蛤仔;杂交;Kung育种值;配合力
相比于玉米、水稻、小麦等农作物育种工作的开展,中国贝类遗传育种研究则起步较晚。目前,经过贝类育种专家的多年努力,中国已相继培育出“大连1号”、“西盘鲍”等杂交鲍新品种,“中科红”、“海大金贝”、“蓬莱红”、“海益丰12”等扇贝新品种,“海大1号”、“海大2号”、“华南1号”、“金蛎1号”等牡蛎新品种,“斑马蛤”、“白斑马蛤”等蛤仔新品种,“科浙1号”、“万里红”等文蛤新品种,“乐清湾1号”等泥蚶新品种,以及多个珠母贝新品种。由此可见,近年来中国的贝类遗传育种工作进展良好,在贝类产业发展中起到了积极地推动作用。贝类育种方法也由最初的单一起步摸索阶段,逐渐发展为综合多种育种方法的多性状复合育种阶段。
在贝类遗传育种中,性状纯化和杂种优势利用是研究者关注的焦点,其中,家系建立是遗传基础研究和新品种选育的重要手段之一。在此过程中,通过建立不同地理群体或同一群体不同优势性状家系,再通过遗传参数、育种值的估算,评估出具有目标优势性状家系,而后,在此基础上设计不同家系间的杂交组合,构建闭锁选育群,通过逐代家系间杂交选育,培育出多个优势性状聚合的新品种[1]。目前,采用Kung育种值和配合力评估具有目标优势性状的杂交组合的预测方法是育种实践中的主要方法之一[2-5],其中,Kung育种值和配合力的测定是利用杂种优势的必要前提[6-8]。
目前,在贝类遗传育种中,杂交育种的研究仅限于同种不同野生群体间或不同种间远缘杂交[9-15]。但野生群体杂合度高、遗传背景复杂,由此导致杂种优势利用效率低,仅为纯系间杂交效率的二分之一[16]。远缘杂交虽能获得较大的杂种优势,但由于亲本间的精卵识别能力差、受精率低,其应用前景也有一定的局限性。家系间杂交是利用基因型相对纯合的家系,相互间杂交,一方面可以提高杂种优势的利用效率,另一方面可以避免配子不亲和由此导致的受精率低的问题[16]。因此,家系间杂交培育新品种(系)的育种方法具有广阔的应用前景。
菲律宾蛤仔Ruditapes philippinarum是中国主要的养殖贝类品种,其产量占中国滩涂贝类总产量的73%。世界每年300多万t的养殖产量主要来自中国大陆[17]。本研究中,选用生长和抗逆性状优良的3个菲律宾蛤仔全同胞家系,采用家系间双列杂交方法,通过估算不同杂交家系的Kung育种值、配合力及杂种优势,评定出最优杂交组合,旨在为菲律宾蛤仔的遗传改良研究提供理论依据。

表1 蛤仔3个全同胞家系的双列杂交设计Tab.1 Diallel crosses among three full-sib families of Manila clam Ruditapes philippinarum
1 材料与方法
1.1 材料
试验在大连海洋大学辽宁省贝类良种繁育工程技术研究中心进行,以已建立的生长和抗逆性状优良的壳面具有斑块花纹的蛤仔家系子一代(C31、C32)及壳面背景颜色为橙色的蛤仔家系子三代(Or)作为基础群体。在每个家系中选取贝壳无损伤、壳型规整、壳色美观、活力强的蛤仔作为亲本。
1.2 方法
1.2.1 试验设计 蛤仔亲本经阴干、流水刺激后,在水温为23℃、盐度为28、pH为7.8的条件下,产卵排精,采用3×3双列杂交的方法构建杂交家系(表1)。
1.2.2 饲养管理 在盛满新鲜海水的100 L塑料桶中培育各试验组幼虫,培育密度为4~5个/mL,每天投喂饵料为湛江等鞭金藻Isochrysis zhangjiangensis和小球藻Chlorella vulgaris,饵料量为2000~5000 cells/(mL·d),分4次投喂。每天半量换水1次,每2天全量换水1次,随着幼虫的生长,筛绢网由300目逐渐更换为100目。为了使培育密度不影响研究结果,在幼虫培育阶段,每2天对培育密度调整1次,使各个试验组密度基本保持一致。各个试验组个体严格隔离,分桶培育。
当幼虫出足时,逐渐将幼虫培育密度调整为1~2只/mL,幼虫附着变态后,稚贝饵料投喂量为10 000~20 000 cells/(mL·d),分4次投喂,根据具体摄食情况可适当增减饵料量,以保持培育海水中饵料充足。稚贝培育至60日龄以后,将稚贝装入60目的网袋(40 cm×60 cm)中,每袋200~300粒,挂养在室外生态池中,浮筏吊养。池塘养殖期间,水温为21~28℃,盐度为24~28,pH为7.64~8.62。
1.2.3 指标的测定 使用显微镜测量3、6、9、15、30日龄蛤仔的壳长,使用游标卡尺测定60、90日龄蛤仔的壳长,每个试验组测量30只个体。
1.3 数据处理
双列杂交分析按Griffing[18]的方法Ⅰ计算。第一步,检测亲本、子代和反交组合间是否存在基因型差异。第二步,当第一步检测差异显著时进一步进行配合力方差分析。配合力分析模型为

其中:Yij为第i×j基因型效应;gi为第i个亲本的一般配合力效应(GCA);gj为第j个亲本的一般配合力效应(GCA);sij为特殊配合力效应(SCA);rij为反交效应;为随机误差效应。
分别计算所有家系的Kung育种值、一般配合力、特殊配合力和杂种优势。
Kung育种值的计算公式[18]为

一般配合力计算公式为
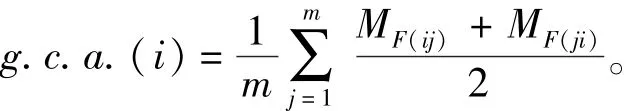
其中:m为假设有m个群体;MF(ij)为第i种群与第j种群杂交子一代群体壳长的平均值;MF(ji)为第j种群与第i种群杂交子一代群体壳长的平均值。
特殊配合力计算公式为
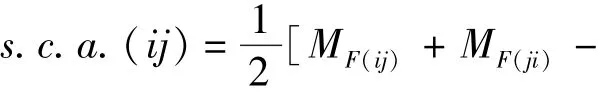

平均母本效应计算公式为

其中:g.c.a.Di和g.c.a.Si分别为第i种群母本和父本的一般配合力。
杂种优势计算公式为

其中:MF1和MF1′为正反交两家系壳长的观测平均值;MP1和MP2分别为亲本纯繁家系壳长的观测平均值。
配合力方差分析和平均值均使用Excel 2000和SAS 8.0软件进行统计计算,所需程序均参照林德光[19]设计步骤。
2 结果与分析
2.1 各家系蛤仔壳长性状Kung育种值的比较
由表2可知:15日龄时,C32(♀)×Or(♂)家系的Kung育种值最高(454.88 μm),C32(♀)× C31(♂)家系的Kung育种值次之(369.48 μm), C31(♀)×C32(♂)家系的Kung育种值最低(247.66 μm);30日龄时,C31(♀)×C32(♂)家系的Kung育种值最高(401.15 μm),C32(♀)× Or(♂)家系的Kung育种值次之(378.88 μm), C31(♀)×Or(♂)家系的Kung育种值最低(317.96μm);60、90日龄时,C31(♀)× C32(♂)家系的Kung育种值均最高(3600、4910 μm),而C31(♀)×C31(♂)家系的Kung育种值均为最低(1350、3820 μm)。
2.2 各杂交家系生长性状的方差分析
由表3可知,3日龄时,各家系生长均无显著性差异(P>0.05),从6日龄起,各家系壳长均有极显著差异(P<0.01)。因此,需从6日龄开始做进一步地配合力分析。
2.3 各杂交家系一般配合力、特殊配合力和反交效应的方差分析
由表4可知:6日龄时,一般配合力有极显著性差异(P<0.01),而特殊配合力和反交效应均无显著性差异(P>0.05);9、30、60、90日龄时,各试验组的一般配合力、特殊配合力和反交效应均有显著或极显著性差异(除60日龄试验组的特殊配合力外),而反交效应从30日龄开始出现极显著性差异(P<0.01),表明正交及反交的交配方式对杂家家系的表现也有显著影响。

表2 蛤仔各家系壳长性状的Kung育种值Tab.2 Kung breeding value of shell length in families of Manila clam Ruditapes philippinarum

表3 各杂交家系家系生长的方差分析Tab.3 Analysis of variance for growth of shell-length in families of Manila clam Ruditapes philippinarum
2.4 各家系间一般配合力、特殊配合力、杂种优势和母本效应的估计
由表5可知:在亲本一般配合力方面,15日龄时,依次为C31>C32>Or,30、60、90日龄时,依次均为C32>C31>Or;而在平均母本效应方面, 15日龄时,依次为C32>C31>Or,30、60、60日龄时,依次均为C31>C32>Or,其中,90日龄时C31和C32的平均母本效应大致相等,C31略大,而在30和60日龄时,亲本C32和Or的平均母本效应均表现为负值。
由表6可知:在特殊配合力方面,15日龄时,依次为C32(♀)×Or(♂)>C31(♀)×C32(♂)> C31(♀)×Or(♂),30、60、90日龄时,依次均为C31(♀)×C32(♂)>C32(♀)×Or(♂)>C31(♀)× Or(♂);而在平均杂种优势方面,15日龄时, C32(♀)×Or(♂)及C31(♀)×Or(♂)家系的子代表现出明显的杂种优势,其中,C32(♀)×Or(♂)家系的子代优势最明显,到30日龄时,C31(♀)× C32(♂)家系的子代群体平均杂种优势逐渐明显,并在此后各时期均为3种组合中优势最明显的组合。
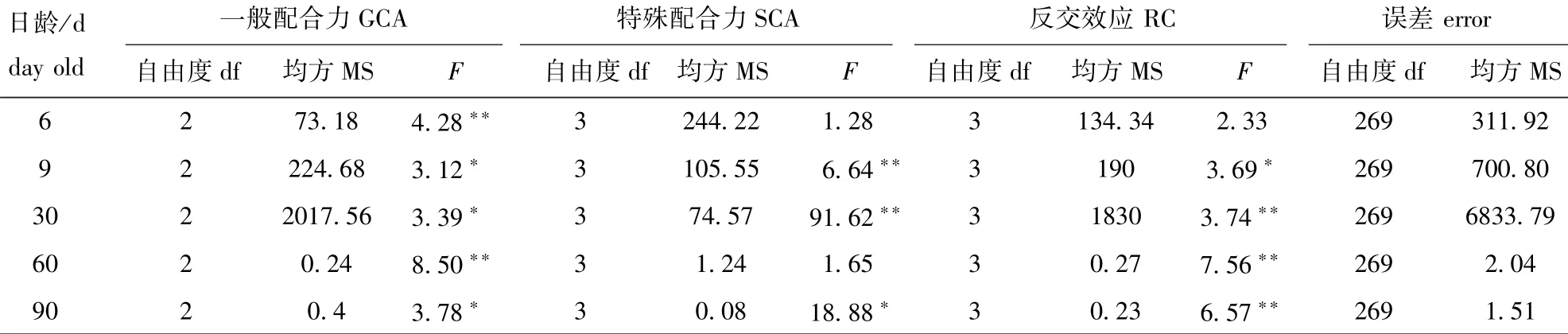
表4 一般配合力、特殊配合力和反交效应的方差分析Tab.4 Analysis of variance of GCA,SCA and reversed cross in families of Manila clam Ruditapes philippinarum
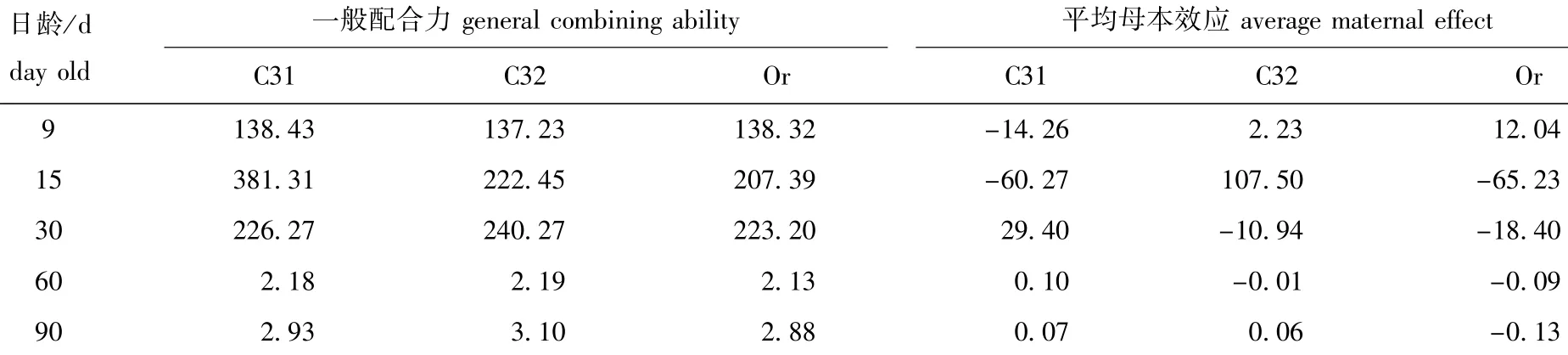
表5 蛤仔各家系一般配合力和母本效应Tab.5 General combining ability and average maternal effect in mixed families of Manila clam Ruditapes philippinarum

表6 蛤仔各家系特殊配合力和平均杂种优势Tab.6 Specfic combining ability and average heterosis in families of Manila clam Ruditapes philippinarum
3 讨论
3.1 菲律宾蛤仔选育家系间的Kung育种值分析
动物育种的目的是获得稳定遗传的优良性状,水产动物选择育种所重视的许多经济性状均为数量性状,如体质量、体长、出肉率等。数量性状的表型值是基因型值和环境共同影响的结果,而基因型存在3种效应,分别为等位基因间的显性效应、基因加性效应和非等位基因间的上位效应[20]。育种值是决定数量性状的基因加性效应值,是育种工作的重要指标,其中,Kung育种值是育种值中的一种,其数值大小表明了该性状能够顺利遗传给下一代的能力,育种值越大,说明育种效果越好,是家系选择中某一性状选育的重要参考值[21-22]。本试验中,C31(♀)×C32(♂)家系从30日龄开始,便是各家系中Kung育种值最高的,这表明C31(♀)× C32(♂)家系表现出良好的加性效应。
近年来,彭张明等[22]利用Kung育种值评定了马氏珠母贝红色闭壳肌F1代家系生长、存活等经济性状表现。谷龙春等[23]估计了3个群体合浦珠母贝杂交家系的Kung育种值。李多等[24]通过Kung育种值评定了壳长、壳高具有生长优势的缢蛏家系。刘志刚等[25]报道了海湾扇贝南部亚种的Kung育种值,指出利用高值亲本可提高优秀自交家系的选育。这些研究说明,Kung育种值在贝类育种中应用效果显著。
3.2 菲律宾蛤仔选育家系间的配合力分析
杂交是培育优良品种的重要手段之一,亲本的选择对杂交成功与否具有重要影响。但不是任意两个种群杂交均能获得好的杂种优势,因此,考察亲本性状遗传能力就显得尤为关键。一个种群的一般配合力主要是由基因的加性效应所致,而两个种群杂交的特殊配合力是基于杂交的显性、超显性和上位效应所致,正常情况下,利用一般配合力高的种群杂交可望获得好的杂交效果,而两种群的特殊配合力则是选取性状优良杂交组合的依据之一。因此,配合力的测定是利用杂种优势的必要前提。
本试验中通过对两杂交家系的生长性状方差分析表明,6日龄后,蛤仔幼虫及稚贝一般配合力、特殊配合力和反交效应有显著性差异,这说明蛤仔生长是由加性效应和非加性效应共同影响的。一般配合力方面,从30日龄起,C31(♀)×C32(♂)一直表现出较高的效应值,同时也表现出明显优于Or的母本效应,这表明这两个群体存在较多的加性效应基因可以遗传给子代,以促进子代生长,且雌性个体作为亲本对蛤仔后期生长发育有较大的促进作用,而幼虫期C32(♀)为雌性亲本的各群体有良好的生长存活表现,也是因为C32(♀)×Or(♂)母本效应的结果。在特殊配合力方面,C31(♀)× C32(♂)、C32(♀)×Or(♂)家系的总体表现优于C31(♀)×Or(♂)家系,而这也体现在良好的杂种优势上,尤其C31(♀)×C32(♂)群体,从9日龄后,蛤仔幼虫的壳长生长显著高于其他各群体。综合对各群体生长发育的比较,C31(♀)×C32(♂)、C32(♀)×Or(♂)家系有较高的特殊配合力,适宜分别作为杂交亲本组合,而C31(♀)×C32(♂)家系由于有较高的一般配合力并表现出较好的母本效应,C31(♀)更加适合被选作杂交组合中的雌性亲本。本试验中,C31(♀)×C32(♂)家系取得了最显著的生长优势。在农业生产上,用配合力研究使玉米[26-29]、水稻[30-32]、家蚕[33-34]等的育种研究取得了较大成就。目前,在水产贝类中,关于杂种优势及配合力的报道已见于菲律宾蛤仔[35]、文蛤[36]、鲍等品种[37],这些研究表明,杂种优势和配合力的估算可以辅助贝类群体间或家系间杂交选种工作的开展,并为贝类育种提供借鉴。
本研究中,综合蛤仔选育家系的Kung育种值、配合力、母本效应和杂种优势的分析,得到C31(♀)×C32(♂)家系可作为蛤仔速生品系选育重要的育种材料,同时也为蛤仔选择育种研究提供了有益参考。
[1] Crow J F.Dominance and overdominance[M]//Gowen J W.Heterosis.Ames,IA,USA:Iowa State College Press,1952:282-294.
[2] 梁秋菊.猪亲本及其杂种DNA甲基化差异与杂种优势的关系[D].武汉:华中农业大学,2006:8-27.
[3] Tave D,Smitherman R O,Jayaprakas V.Estimates of additive genetic effects,maternal effects,specific combining ability,maternal heterosis,and egg cytoplasm effects for cold tolerance in Oreochromis niloticus(L.)[J].Aquaculture Research,1989,20(2):159-166.
[4] Wang Xiaoxue,Ross K E,Saillant E,et al.Quantitative genetics and heritability of growth-related traits in hybrid striped bass(Morone chrysops♀×Morone saxatilis♂)[J].Aquaculture,2006, 261(2):535-545.
[5] Romanus K G,Hussein S,Mashela W P.Combining ability analysis and association of yield and yield components among selected cowpea lines[J].Euphytica,2008,162(2):205-210.
[6] 张淑霞,崔健,宋云云,等.配合力在作物育种上的应用[J].现代农业科技,2007(11):94-95.
[7] Hedgecock D,Davis J P.Heterosis for yield and crossbreeding of the Pacific oyster Crassostrea gigas[J].Aquaculture,2007,272:17-29.
[8] 张国范,刘述锡,刘晓,等.海湾扇贝自交家系的建立和自交效应[J].中国水产科学,2003,10(6):441-445.
[9] 周茂德,高允田,吴融.太平洋牡蛎与近江牡蛎、褶牡蛎人工杂交的初步研究[J].水产学报,1982,6(3):235-241.
[10] Manzi J J,Hadley N H,Dillon Jr R T.Hard clam,Mercenaria mercenaria,broodstocks:growth of selected hatchery stocks and their reciprocal crosses[J].Aquaculture,1991,94(1):17-26.
[11] 齐秋贞,林笔水,吴天明,等.菲律宾蛤仔室内催产研究——阴干、氨海水和性诱导法[J].水产学报,1981,5(3):235-243.
[12] Cruz P,Ibarra M.Larval growth and survival of two catarina scallop(Argopecten circularis,Sowerby,1835)populations and their reciprocal crosses[J].Journal of Experimental Marine Biology and Ecology,1997,212(1):95-110.
[13] 刘小林,常亚青,相建海,等.栉孔扇贝中国种群与日本种群杂交一代的中期生长发育[J].水产学报,2003,27(3):193-199.
[14] 刘小林,常亚青,相建海,等.栉孔扇贝不同种群杂交效果的初步研究:Ⅰ.中国种群与俄罗斯种群的杂交[J].海洋学报, 2003,25(1):93-99.
[15] Lamkey K R,Edwards J W.The quantitative genetics of heterosis [C]//Coors J G,Pandey S.Proceedings of the International Symposium on the Genetics and Exploitation of Heterosis in Crops. Mexico City,Mexico:CIMMYT,1999:31-48.
[16] 张国范,刘晓,阙华勇,等.贝类杂交及杂种优势理论和技术研究进展[J].海洋科学,2004,28(7):54-60.
[17] 范超,温子川,霍忠明,等.盐度胁迫对不同发育时期菲律宾蛤仔生长和存活的影响[J].大连海洋大学学报,2016,31 (5):497-504.
[18] Griffing B.Concept of general and specific combining ability in relation to diallel crossing system[J].Australian Journal of Biological Sciences,1956,9:463-493.
[19] 林德光.双列杂交育种法配合力分析的SAS实施[J].热带海洋学报,1999,20(2):44-52.
[20] 盛志廉,陈瑶生.数量遗传学[M].北京:科学出版社,1999.
[21] 刘辉,张兴志,鹿瑶,等.菲律宾蛤仔橙色品系壳形态性状对质量性状的通径及多元回归分析[J].大连海洋大学学报, 2015,30(5):514-518.
[22] 彭张明,陶后全,刘锦上,等.马氏珠母贝红色闭壳肌F1代的家系选育及家系评定[J].广东海洋大学学报,2015,35(4):37-45.
[23] 谷龙春,李金碧,喻达辉,等.合浦珠母贝双列杂交家系的建立与遗传分析[J].水产学报,2010,34(1):26-31.
[24] 李多,杜文俊,牛东红,等.缢蛏家系早期生长表现比较[J].中国海洋大学学报:自然科学版,2015,45(10):36-41.
[25] 刘志刚,章启中,朱晓闻,等.海湾扇贝南部亚种自交家系选育及其Kung育种值评价[J].中国水产科学,2013,20(2): 308-315.
[26] 秦艳杰,刘晓,张海滨,等.海湾扇贝正反交两个家系形态学指标比较分析[J].海洋科学,2007,31(3):22-27.
[27] 丰光,李妍妍,卢秉生,等.对玉米育种中配合力和性状选择问题的探讨[J].中国种业,2010(6):40-41.
[28] 李高科,胡建广,刘建华,等.优质、抗逆甜玉米群体配合力及遗传潜势分析[J].中国农学通报,2008,24(9):145-149.
[29] 蒙成,李体琛.玉米自交系“南99”的选育及配合力分析[J].中国农学通报,2010,26(7):112-114.
[30] 王昌贵,赵鹏珂,冯小磊,等.3个旱稻品种杂交配合力及其应用分析[J].中国农业大学学报,2010,15(6):1-5.
[31] 陈顺辉.水稻两系法亚种间杂种优势和配合力研究[J].北京农业大学学报,1993,19(S):15-20.
[32] 付新民,王岩,高冠军,等.利用水稻重组自交系进行配合力遗传分析[J].华中农业大学学报,2010,29(4):397-402.
[33] 杨仁奎.家蚕配合力效应及杂种优势研究[J].蚕学通讯, 2009,29(2):1-5.
[34] 秦利,冯祖绳,高玉章.双列杂交在柞蚕配合力测定上的应用[J].辽宁农业科学,1991(2):30-33.
[35] Huo Zhongming,Yan Xiwu,Zhao Liqiang,et al.Larval and juvenile growth performance of Manila clam hybrids of two full-sib families[J].Journal of Ocean University of China,2015,14(3):564-568.
[36] 吴杨平,陈爱华,姚国兴,等.3个不同地理群体红壳色文蛤杂交的配合力分析[J].海洋渔业,2014,36(4):314-319.
[37] Deng Yuewen,Liu Xiao,Zhang Guofan,et al.Heterosis and combining ability:a diallel cross of three geographically isolated populations of Pacific abalone Haliotis discus hannai Ino[J].Chinese Journal of Oceanology and Limnology,2010,28(6):1195-1199.
Analysis of Kung breeding value and combining ability of hybridization of selective families of Manila clam Ruditapes philippinarum
SUN Qian,DUAN Qiang,SANG Shi-tian,HUO Zhong-ming,YAN Xi-wu
(College of Fisheries and Life Science,Engineering Research Center of Shellfish Culture and Breeding in Liaoning Province,Dalian Ocean University, Dalian 116023,China)
Complete diallel crosses were carried out among two plaque pattern families F1(C31,C32)of Manila clam Ruditapes philippinarum selected for good growth and high resistance,and an orange family F3(Or),and Kung breeding value and combining ability of mixed families were analyzed in order to evaluate the genetic effects on the hybridization of Manila clam families.The results showed that the individuals in family C32(♀)×Or(♂) had the maximal Kung breeding value at day 15,and that the individuals in family C31(♀)×C32(♂)had the maximal Kung breeding value from day 30 to 90 day.There were very significant differences in general combining ability(GCA)(P<0.01),without significant differences in specific combining ability(SCA)(P>0.05)at day 6 among the hybrid families,indicating that additive effect influences the growth.There were significant differences in GCA,SCA and reversed cross effect(P<0.05)since day 6,especially in parent C31(♀)×C32(♂),showing that the C31(♀)and C32(♂)families are the selected parents for hybridization to breed good growth strain of Manila clam.
Ruditapes philippinarum;hybrid;Kung breeding value;combining ability
Q321.4
A
10.16535/j.cnki.dlhyxb.2017.04.001
2095-1388(2017)04-0381-06
2017-05-14
国家现代农业产业技术体系建设专项(CARS-48)
孙谦(1983—),男,硕士。E-mail:sunqian@dlou.edu.cn
霍忠明(1983—),男,博士,讲师。E-mail:huozm@dlou.edu.cn
