甜菜褐斑病抗性基因的克隆
2017-07-24刘大丽马龙彪纪岩张革艳刘丽萍鲁振强黑龙江省高等学校生物化学与分子生物学重点实验室黑龙江大学生命科学学院哈尔滨50080黑龙江省高等学校甜菜遗传育种重点实验室黑龙江大学农作物研究院哈尔滨50080
刘大丽,马龙彪,纪岩,张革艳,刘丽萍,鲁振强*(.黑龙江省高等学校生物化学与分子生物学重点实验室/黑龙江大学生命科学学院,哈尔滨50080;.黑龙江省高等学校甜菜遗传育种重点实验室/黑龙江大学农作物研究院,哈尔滨50080)
甜菜褐斑病抗性基因的克隆
刘大丽2,马龙彪2,纪岩1,张革艳1,刘丽萍1,鲁振强1*
(1.黑龙江省高等学校生物化学与分子生物学重点实验室/黑龙江大学生命科学学院,哈尔滨150080;2.黑龙江省高等学校甜菜遗传育种重点实验室/黑龙江大学农作物研究院,哈尔滨150080)
为了获得抗病的甜菜遗传材料,通过RT-PCR技术,分别对两类真菌抗性基因AtChitinase1(GenBank No.NM_100117)和AtGlucanase2(GenBank No.NM_115586)的功能域进行了克隆;将所得的片段连接到载体pMD 18-T并鉴定。结果表明,AtChitinate1基因的核苷酸长度为819 bp,编码272个氨基酸,AtGlucanase2基因的核苷酸长度为1020 bp,编码339个氨基酸,确认获得两个抗病基因功能片段。
甜菜;褐斑病;几丁质酶;葡聚糖酶;基因克隆
甜菜(Beta vulgaris L.)是我国的主要糖料作物,在我国东北、西北和华北等地大面积种植。甜菜褐斑病是Cercospora beticola Sacc.引起的真菌类病害,病症初期表现为叶片局部褐色斑块,继发则导致叶片全部坏死,干扰了甜菜正常的生长发育,破坏了植物正常的生理生化反应,对甜菜的产量品质影响极大。使用杀菌剂抗病,在生产中不仅需要多种杀菌剂联合轮换使用,而且在病害重发区域需要多次喷施杀菌剂处理,大量长期的使用势必污染环境[1-2]。从遗传上入手,通过导入真菌病害抗性基因,培育抗褐斑病甜菜品种,具有重要的意义与价值。几丁质酶(Chitinase)是植物内的一类水解酶,而几丁质正是真菌病原菌细胞壁的主要成分,通过破坏真菌细胞结构,从而抑制了病原菌的生长。当病原菌侵染植物细胞,植物几丁质酶受到诱导进而表达,这时病菌菌丝体的细胞壁被几丁质酶水解破坏而变薄,引发球状突起,进而质膜破裂,菌丝的生长和孢子萌发受到抑制,从而达到抑制病原菌生长的效果。葡聚糖酶(Glucanase)是一类植物抗病蛋白,一方面它的催化产物可以诱发植物抗病反应,寡糖是葡聚糖酶的酶解产物,寡糖是一种诱发植物抗病防御反应的信号分子,可以诱导植物建立复杂的抗病防御系统,间接增强植物的抗病能力。另一方面,葡聚糖酶可以直接降解真菌细胞,使得病菌细胞内含物外溢,抑制病原菌菌丝生长和孢子萌发。Chitinase基因和Glucanase基因是目前发现的两种广谱性抗真菌病害基因,并且这两个基因具有协同增效的作用[3]。
基于这个背景与需求,本研究以提高甜菜对褐斑病的遗传抗性为目标,采用RT-PCR技术对抗性基因AtChitinase1和AtGlucanase2的功能域分别进行了克隆,为后期对甜菜褐斑病的遗传研究及获得抗病甜菜品种奠定基础。
1 材料和方法
1.1 实验材料与主要试剂
选用Ecotype Colombia型Arabidopsis thaliana为靶基因克隆的植物材料。BIOZOL(BioFlux)、pMD18-T载体、胶回收试剂盒(TaKaRa);大肠杆菌DH5α(本实验室);RNA电泳所用试剂均为进口分析纯。
1.2 试验方法
1.2.1 总RNA的提取与检测称取1 g植物幼嫩叶片组织,加入5mL BIOZOL试剂中,室温15min;离心;将上层清液中加入1/5体积的氯仿,冰浴15min;再次离心,将上层清液中加入等体积异丙醇,于-20℃放置30min;离心后,将沉淀于1mL 70%乙醇中洗涤两次,晾干后,溶于DEPC水中,储存于-70℃备用。利用紫外分光光度计,分别在OD260和OD280条件下测定RNA的含量和纯度,并将样品的浓度稀释成1μg/μL备用。取2μg总RNA,加入15.8μL变性液,电泳缓冲液为1×MOPs,经1.0%琼脂糖电泳,进一步确定RNA质量。
1.2.2 引物设计根据AtChitinase1(GenBank No.NM_100117)基因和AtGlucanase2(GenBank No. NM_115586)基因的序列信息,利用Primer premier 5.0分别设计了两个基因的克隆引物,预计扩增AtChitinase1基因的CDS功能域长度为819 bp,扩增AtGlucanase2基因的CDS功能域长度为1020 bp。扩增引物由上海生工合成。
1.2.3 RT-PCR实验在12μL的反应体系中加入3μg RNA、10μM Oligo dT以及RNase free H2O于65℃反应5min,并立即置于冰上;向第一步变性反应液中加入5×RTBuffer,20mM dNTPs,10 U RNase Inhibiter以及20U的Rever Tra Ace(ToYoBo),总体积为20μL。反应条件为42℃60min;99℃5min;4℃5min。所得的反转录产物保存于-20℃备用。PCR扩增:50μL的反应体系中,分别加入2μL cDNA模板,5μL 10×Taq Buffer、4μL 10mM dNTPs、1μL 20μM上游引物和下游引物,0.5μL rTaq(TaKaRa)。PCR扩增条件为:94℃预变性5min;94℃(30s),52℃(45s),72℃(3min),30个循环;最后72℃延伸10min。扩增产物于1.0%琼脂糖凝胶中进行电泳,并利用GeneSnap 6.08(f)进行检测。
1.2.4 酶切验证及序列测定将扩增产物进行电泳,回收;将pMD18-T载体和目的基因回收片段按照摩尔比1∶5进行连接,转化到DH5α感受态细胞中,通过Amp抗性及蓝白斑筛选出待测阳性克隆。待测阳性克隆经过提质粒,酶切鉴定,最终进行测序验证。

图1 AtChitinase1基因和AtGlucanase2基因CDS功能域的RT-PCR扩增A:总RNA;B、C:RT-PCR扩增;1:AtChitinase1基因CDS;2:AtGlucanase2基因CDS;M(Maker):1 kb DNA Ladder(NEB)
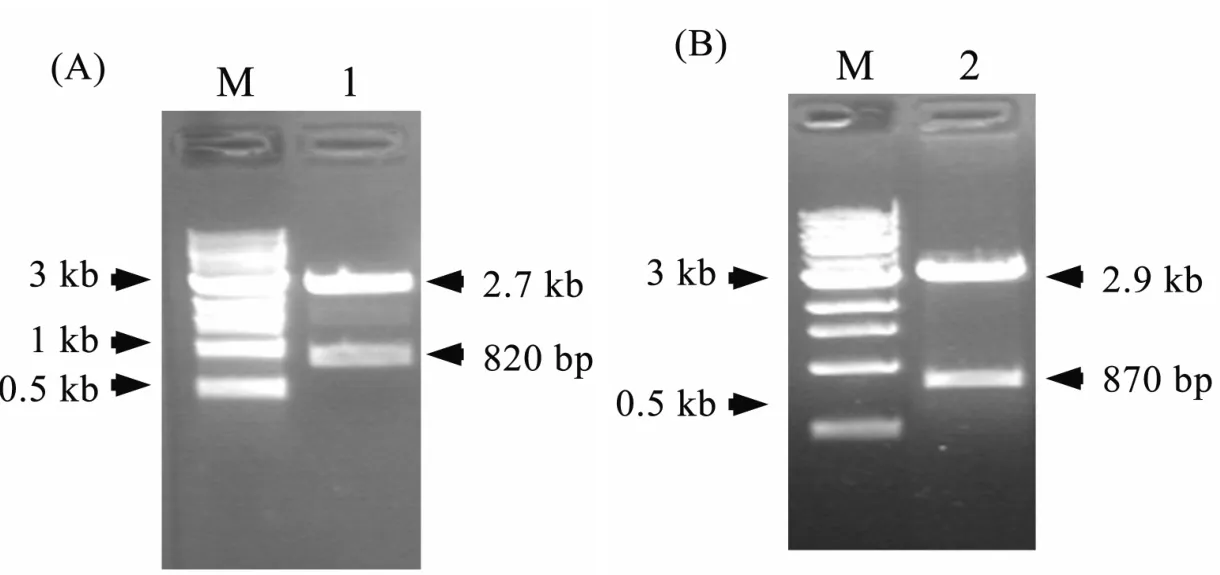
图2 pMD 18-T-AtChitinase1和pMD 18-T-AtGlucanase2载体的构建A.pMD 18-T-AtChitinase1载体质粒酶切鉴定;B.pMD 18-T-AtGlucanase2载体质粒酶切鉴定;1:SalI和XbaI;2:HincII;M(Maker):1 kb DNA Ladder(NEB)
2 结果与分析
2.1 AtChitinase1和AtGlucanase2基因的克隆
实验以Ecotype Colombia型Arabidopsis thaliana为靶基因克隆的植物材料,利用BIOZOL试剂提取了总RNA(图1A)。电泳后28S rRNA和18S rRNA在紫外灯下清晰可见,并且,28S rRNA的亮度是18S rRNA的2倍左右,这说明RNA基本上没有被降解,且无弥散。同时,加样孔附近没有杂带,说明产物中没有DNA存在;在18S rRNA下方存在一些较为模糊的条带,这可能是由tRNA、5.8SRNA和5S rRNA组成的迁移较快的条带。这些结果都说明了实验获得了较高纯度和得率的总RNA。
以总RNA为模板,利用AMV获得了cDNAs,再通过RT-PCR扩增,显示在820 bp的位置获得了片段(图1B),这与AtChitinase1(GenBank No. NM_100117)基因序列功能域的大小—819 bp相一致;在1020 bp的位置获得的片段(图1C),与AtGlucanase2(GenBank No.NM_115586)基因序列的功能域1020 bp相符。
2.2 重组质粒pMD 18-T-AtChitinase1和pMD 18-T-AtGlucanase2的构建
将PCR获得的AtChitinase1和AtGlucanase2基因片段分别进行回收,与pMD 18-T连接,并转化到DH5α感受态细胞中,涂布于含有Amp、IPTG和X-gal的LB固体培养基上。挑取白斑阳性克隆,并对应提取质粒DNA。通过对AtChitinase1和AtGlucanase2基因CDS序列限制性内切酶图谱的分析,得出AtChitinase1基因内部无常规酶切位点,而在AtGlucanase2基因CDS区域850 bp位置有一个HincII限制性内切酶酶切位点。因此,选取pMD 18-T载体上的SalI和XbaI,对pMD 18-T-AtChitinase1重组质粒进行酶切鉴定;对于pMD 18-T-AtGlucanase2重组质粒来说,选取同时存在于载体上游及基因内部HincII来进行鉴定(图2)。
如图2A所示,经过SalI和XbaI酶切重组质粒,获得了两条位于2.7 kb和820 bp左右位置的条带,这与T载体(2690 bp)和AtChitinase1全长(819 bp)长度基本一致。
由图2B可见,经过SacI酶切重组质粒,分别于2.9 kb和870 bp左右的位置获得了两条酶切条带。由于AtGlucanase2基因内部的HincII酶切位点位于其起始密码子端的850 bp的位置,因此,870 bp的片段长度意味着在重组质粒中,AtGlucanase2基因逆向插入到pMD 18-T载体上。
3 讨论
迄今为止,虽然美国、欧洲等抗真菌、细菌、病毒等病甜菜品种已研制成功,但其申请了专利保护,输入并控制了包括中国在内的许多国家甜菜抗病商用品种的价格,已形成了全球的垄断趋势[4]。本研究对甜菜抗褐斑病基因—几丁质酶AtChitinase1和葡聚糖酶AtGlucanase2基因的克隆,不仅可以打破国外的垄断,培育自主产权的抗病甜菜品种,而且对于提高甜菜产量、增加农民收入,促进经济发展具有重要的科学意义。
[1]罗成飞,孔凡江.谈甜菜抗褐斑病育种[J].中国甜菜糖业,2000(4):28-30
[2]乔志文,李彦丽,柏章才.3种杀菌剂防治甜菜褐斑病间隔期及效果[J].中国农学通报2015,31(29):72-76
[3]Mebeaselassie Andargie,Jianxiong Li.Arabidopsis thaliana:A Model Host Plant to Study Plant–Pathogen Interaction Using Rice False Smut Isolates of Ustilaginoidea virens[J].Frontiers in plant Sci,2016,7:1-10
[4]James C.Global status of commercialized biotech/GM Crops:2008[R].NY:ISAAA,2008.
M olecular Cloning of Resistant Genes Related to Sugarbeet Cercospora beticola
LIU Da-li2,MA Long-biao2,JIYan1,ZHANG Ge-yan1,LIU Li-ping1,LU Zhen-qiang1*
(1.Key Laboratory of Biochemistry and Molecular Biology/College of Life Sciences,Heilongjiang University,Harbin 150080,China; 2.Key Laboratory of SugarbeetGenetics and Breeding/Academy of Crop Sciences,Heilongjiang University,Harbin 150080,China)
To improve the resistance of sugarbeet to Cercospora beticola Sacc.,target genes AtChitinase1 (GenBank No.NM_100117)and AtGlucanase2(GenBank No.NM_115586)were selected,and the CDS of two geneswere cloned respectively by RT-PCR.Two CDSwere constructed into pMD 18-T Vector respectively,and identified with enzymes digested and sequenced.AtChitinase1 and AtGlucanase2 were 819 bp and 1020 bp, deduced to encode 272 and 339 amino acids respectively,and identified as predicted.
sugar beet(Beta vulgaris);cercospora beticola Sacc.;chitinase;glucanase;gene cloning
S566.3;Q 78
A
1007-2624(2017)04-0005-02
10.13570/j.cnki.scc.2017.04.002
2017-03-30
黑龙江省教育厅资助项目(12541608)。
刘大丽(1981-),女,博士,主要从事植物分子生物学研究。Tel:0451-86609494;E-mail:daliliu2007@sina.com
鲁振强,博士,硕士生导师,主要从事植物研究。E-mail:zhenqianglu@163.com
