野生大豆育成品种与其亲本间的SSR聚类分析
2017-07-15吕祝章王文哲梁青邱丽娟
吕祝章+王文哲+梁青+邱丽娟
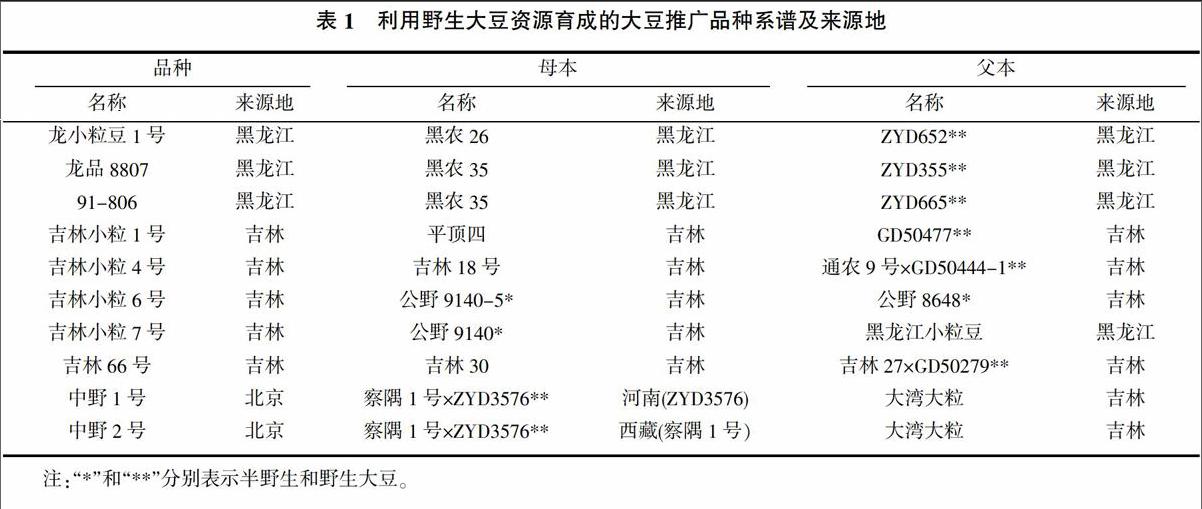
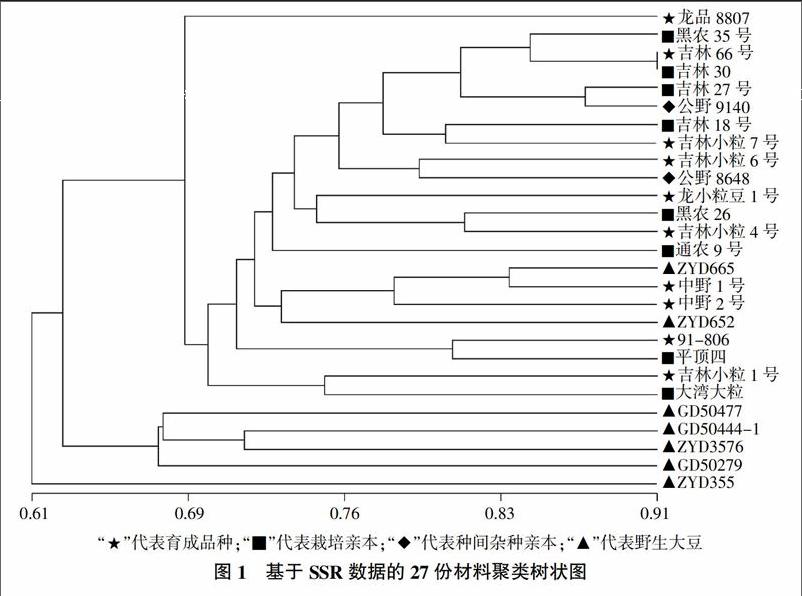
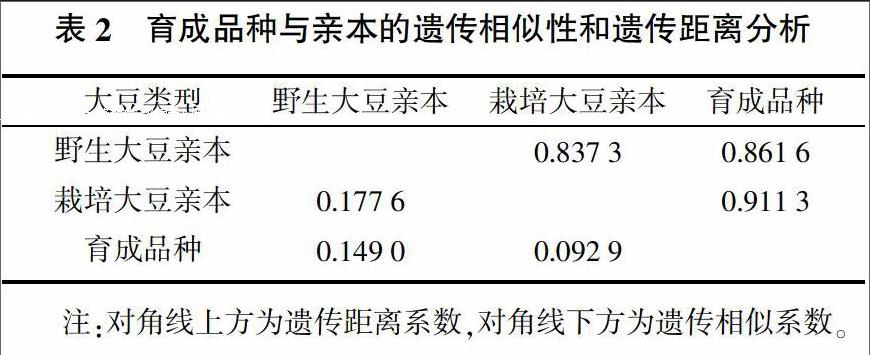
摘要:采用分布于大豆(Glycine max)全基因组的90对SSR引物,对来自全国在生产上大面积推广应用的具有野生大豆(Glycine soja)血缘的10个大豆育成品种及其17个亲本材料进行聚类和遗传距离分析。结果表明,当遗传相似系數为0.67时,所有供试材料被分成三类,即育成的大豆品种与其栽培大豆亲本聚为一类,野生大豆亲本在三类中均有分布;野生大豆亲本与栽培大豆亲本间的遗传距离0.177 6>野生大豆亲本与育成大豆品种间的遗传距离0.149 0>栽培大豆亲本与育成大豆品种间的遗传距离0.092 9。表明野生大豆与栽培大豆间有较大的遗传差异性,应用遗传基础不同及遗传差异性较大的野生大豆与栽培大豆之间进行杂交选择,进而拓宽大豆的遗传基础、丰富大豆遗传多样性是有效和可行的。
关键词:野生大豆(Glycine soja);育成品种;聚类分析;遗传距离;SSR
中图分类号:S565.1 文献标识码:A 文章编号:0439-8114(2017)12-2215-04
DOI:10.14088/j.cnki.issn0439-8114.2017.12.005
SSR Cluster Analysis between Cultivars and Their Parents in Wild Soybean
L?譈 Zhu-zhang1,WANG Wen-zhe1,LIANG Qing1,QIU Li-juan2
(1. Rizhao Polytechnic College, Rizhao 276826, Shandong, China; 2. The Key Open Laboratory of Crop Germplasm and Biotechnology of the Ministry of Agriculture/the Major Project of Crop Gene Resources and Genetic Improvement of China/Crops Research Institute, Chinese Academy of Agricultural Sciences, Beijing 100081, China)
Abstract: 90 pairs of SSR primer distributed in soybean genome were used for clustering and genetic distance analysis of 10 improved soybean varieties with wild soybean blood and their 17 parental materials. Results showed that when the genetic similarity coefficient was 0.67,all tested materials were divided into three groups, namely improved soybean varieties and cultivated soybean parents together as a class,wild soybean parents were distributed in all three categories. The genetic distance of 0.177 6 between wild soybean parents and cultivated soybean parents>the genetic distance of 0.149 0 between wild soybean parents and improved soybean varieties > the genetic distance of 0.092 9 between cultivated soybean parents and improved soybean varieties.It indicated that there were large genetic differences between wild and cultivated soybean, and it was feasible and effective to expand soybean genetic basis and enrich soybean genetic diversity by hybridization and selection between wild soybean and cultivated soybean with different genetic basis and big genetic difference.
Key words: wild soybean(Glycine soja); improved variety; cluster analysis; genetic distance; SSR
种质资源是基因的载体,是遗传改良和育种的基础,育种工作成就的大小,在很大程度上取决于对种质资源的了解、掌握和利用程度。纵观世界近代和现代农业三次跨越式发展,作物种质资源的发掘与利用都起到了巨大作用[1-6]。中国是大豆(Glycine max)的发源地,有占世界90%以上的丰富的野生大豆(Glycine soja)资源。野生大豆被认为是提高栽培大豆蛋白质含量、改善品质、增强抗性的重要基因来源[7]。近三十年来,中国大豆育种工作者也重视和利用野生大豆资源开展大豆育种工作,创新培育出获国家发明奖的铁丰18号和吉林小粒1号,获省级科技进步奖的杂交豆1号、龙品8807和龙小粒豆1号,先后有18个育成品种被国家和省级审定,取得了骄人的成绩。
近年来分子标记技术在作物亲缘关系及其遗传相似性研究方面发挥了重要作用[8],已有利用分子标记和系谱资料综合分析作物品种间亲缘关系及其遗传相似性的研究报道[9-12],但应用SSR对利用野生大豆育成品种进行系谱分析和聚类分析的研究报道较少。
本研究选择了10个在生产上大面积推广应用的具有野生大豆血缘的大豆育成品种及其17个亲本作为供试材料,对分布于大豆全基因组的90个SSR座位进行分析,旨在分子水平上明确大豆育成品种与其亲本间的遗传与演化关系,探索利用野生大豆资源拓宽中国大豆遗传基础的趋势,为进一步有效地利用野生大豆资源开展大豆育种工作提供参考信息。
1 材料与方法
1.1 试验材料
本研究选用了10个用野生大豆做亲本育成的在生产上大面积推广应用的大豆品种及其9份野生大豆亲本和8份栽培大豆亲本为供试材料,亲本材料中有察隅1号和黑龙江小粒豆未被搜集到。各品种的杂交组合方式及亲本来源地见表1。
1.2 试验方法
1.2.1 SSR分析 每份材料用种子磨成豆粉,用SDS法[13]提取DNA。PCR反应体系为20μL,其中含有40 ng基因组DNA模板、1×PCR 缓冲液、1.25 mmol/L MgCl2、0.2 mmol/L dNTP、0.2 μmol/L SSR引物和1 U Tag DNA 聚合酶。選用90对SSR引物。反应在PE-9600型号的PCR扩增仪上进行。反应程序为95 ℃预变性4 min;95 ℃变性30 s,47 ℃退火30 s,72 ℃延伸30 s,运行35个循环;72 ℃延伸10 min后置于4 ℃保存。扩增产物用6%聚丙烯酰胺、8 mol/L脲素制成的测序胶分离,银染技术检测。
1.2.2 统计分析 每个SSR位点,以1和0记录等位变异的有和无,缺失记为2,获得矩阵,用NTSYS 2.10进行聚类分析,在遗传相似系数矩阵的基础上,用UPGMA法构建27份材料的分子系统树。
2 结果与分析
2.1 育成品种与亲本材料的聚类分析
利用UPGMA法对获得的SSR分子数据进行聚类,当遗传相似系数为0.67时,将所有供试材料分成了三大类(图1)。一类是囊括供试的10份大豆育成品种和8份栽培大豆亲本,另外还包括了4份分别原产于黑龙江省双城县的野生大豆ZYD652、ZYD665和原产于吉林省的半野生大豆公野8648、公野9140;在这一类中,大豆育成品种龙品8807的位置显得比较独特,可能是由于该育成品种较多地遗传利用了野生大豆亲本ZYD355的10个特异等位变异,且其自身在又3个位点上产生出了3个新的等位变异,再加上原产于黑龙江省佳木斯市的ZYD355在聚类图中单独聚为了一类,表现出了其独树一帜的一面,因而导致了育成品种龙品8807在聚类图(图1)中所表现出的与众不同;在这一类中,吉林66号和它的栽培大豆亲本吉林30在0.91处仍被聚在一起,这可能是由于吉林66号仅利用了其野生大豆亲本GD50279在Satt012位点上的1个特异等位变异,而较多地选择利用了栽培大豆亲本吉林30的特异等位变异。另两类均为野生大豆亲本材料,其中一类仅包括ZYD355一份野生大豆材料,另外一类包括4份分别原产于吉林省的GD50279、GD50444-1、GD50477和原产于河南省济阳县的ZYD3576野生大豆亲本材料。在聚类图(图1)中,供试的9份野生大豆亲本在三类中均有分布,并未与其具有亲缘关系的育成品种聚在一起,表现出较大的遗传差异性,这可能不仅与其自身不同的进化、生态及基因类型有关,同时也与育种目标的定向选择因素影响有关,表明野生大豆的遗传背景比较复杂,遗传多样性也更为丰富。
2.2 育成的大豆品种与其亲本间的遗传距离和遗传相似性分析
对育成的大豆品种与其亲本间的遗传距离的分析表明:野生大豆亲本与栽培大豆亲本间的遗传距离(0.177 6)最大,栽培大豆亲本与育成品种间的遗传距离(0.092 9)最近,而野生大豆亲本与育成品种间的遗传距离(0.149 0)介于两者之间。育成的大豆品种与其亲本间的遗传相似性分析表明:野生大豆亲本与栽培大豆亲本间的遗传相似性(0.837 3)最差,栽培大豆亲本与育成品种间的遗传相似性(0.911 3)最为接近,而野生大豆亲本与育成品种间的遗传相似性(0.861 6)介于两者之间,其遗传相似性分析与遗传距离分析结果一致(表2)。由此可见,野生大豆与栽培大豆的遗传距离较大,遗传相似性较差,遗传关系远缘,存在着较大的遗传差异性;育成品种与栽培大豆亲本遗传距离较近,并有较好的遗传相似性,这可能与在杂交育种的选择过程中,有意淘汰掉了栽培性状表现较劣的分离后代,从中选择保留了产量、品质等与栽培大豆相近的符合育种目标要求的后代等因素有关,从而也导致了育成品种较多地利用了其栽培大豆亲本的特异等位变异,而较少地利用了其野生大豆亲本的特异等位变异,这可能也是造成育成品种与其野生大豆亲本间的遗传相似性及遗传距离介于上述两种情况之间的主要原因之一。综上所述,应用地理远缘或种间远缘的遗传基础不同及遗传差异性较大的野生大豆与栽培大豆之间进行杂交育种选择,进而拓宽大豆的遗传基础、丰富大豆遗传多样性是有效和可行的。
3 讨论
当前大豆育种工作存在的主要问题是亲本遗传基础狭窄[14-16],杂交后代主要经济性状变幅较小,难以产生新的突破性品种,因而造成大豆育成品种的遗传背景及遗传多样性有变窄的趋势[17-20]。综合以上的分析结果可以看出,扩大栽培大豆遗传变异的潜在来源是野生大豆。过去虽然对野生大豆优异种质的筛选与评价开展了大量有益的研究,从野生大豆中筛选和创新出了一大批优异种质,但从围绕野生大豆资源如何加以开发与利用方面还显得明显不足,一是筛选与鉴定的种质不够全面,某些重要性状的筛选与鉴定只集中在部分野生大豆种质上;二是对野生大豆的抗病虫、抗逆境、耐低营养因子、异黄酮含量等的性状鉴定还显不足;三是对野生大豆含有的优异隐蔽基因的发掘工作还没有开展。
育种实践表明,目前利用主要农艺性状优良、配合力高的当地栽培品种间的杂交进行品种选育,仍可提高大豆的增产潜力,但这种现象在一定程度上减弱了人们对野生大豆资源加以利用的兴趣,从而限制了大豆遗传基础的拓宽和遗传多样性的丰富。这给人们一点重要启示,即在大豆遗传育种选择中,既要逐渐引用和渗入遗传基础和遗传差异性大的新种质进行杂交,以拓宽栽培大豆品种的遗传基础,提高大豆新品种的增产潜力,同时也要利用当地优良品种进行品种间杂交,这样育成适应性强的高产、优质、多抗品种的机率会更大些。
参考文献:
[1] MARTIN G B. Gene discovery for crop improvement[J].Current Opinion in Biotech,1998,9:220-226.
[2] KOORNNEEF M,STAM P.Changing paradigms in plant breeding[J].Plant Physio,2001,125:156-159.
[3] SKOVMAND B,REYNOLDS M P,DELACY I H. Mining wheat germplasm collections for yield enhancing traits[J]. Euphytica,2001,119:25-32.
[4] MORGANTE M,SALAMINI F. From plant genomics to breeding practice[J].Current Opinion in Biotech,2003,14:214-219.
[5] 贾继增,张启发.为第二次”绿色革命”发掘基因资源[J].中国基础科学,2001,1(7):4-8.
[6] 贾继增,黎 裕.植物基因组学与种质资源新基因发掘[J].中国农业科学,2004,37(11):1585-1592.
[7] 吴 禹,陈爱国,王 岩.辽宁省部分地区野生大豆资源遗传多样性分析[J].辽宁农业科学,2015(4):1-7.
[8] NARVEL J M,FEHR W R,CHU W S,et al. Simple sequence repeat diversity among soybean plant introductions and elite genotypes[J]. Crop Sci,2000,40:1452-1458.
[9] 熊冬金,王吴彬,赵团结,等.中国大豆育成品种10个重要家族的遗传相似性和特异性[J].作物学报,2014,40(6):951-964.
[10] 熊冬金,赵团结,盖钧镒.中国大豆育成品种亲本分析[J].中国农业科学,2008,41(9):2589-2598.
[11] 张 军,赵团结,盖钧镒.我国黄淮和南方主要大豆育成品种家族产量和品质优异等位变异在系谱中遗传的研究[J].作物学报,2009,35(2):191-202.
[12] 关荣霞,郭娟娟,常汝镇,等.国外种质对中国大豆育成品种遗传贡献的分子证据[J].作物学报,2007,33(9):1393-1398.
[13] 关荣霞,常汝镇,邱丽娟.用于SSR分析的大豆DNA的快速提取[J].大豆科学,2003,21(1):73-74.
[14] 廖 林,刘玉芝,卢亦军,等.吉林省大豆新品种(系)血缘组成分析[J].吉林农业科学,1997(2):1-6.
[15] 邱家驯,赵团结,盖钧镒.中国大豆育成品种中苏沪地区种质的遗传贡献[J].南京农业大学学报,1997,20(4):1-8.
[16] 张 磊,戴瓯和,朱国富,等.皖豆系列大豆品种系谱分析[J].安徽农业科学,2000,28(2):139-140,142.
[17] 盖钧镒,崔章林.中国大豆育成品种的亲本分析[J].南京农业大学学报,1994,17(3):19-23.
[18] 盖钧镒,赵团结,崔章林,等.中国1923-1995年育成的651个大豆品种的遗传基础[J].中国油料作物學报,1998,20(1):17-23.
[19] GIZLICEL Z. Genetic base for North American public soybean cultivars released between 1947-1988[J].Crop Sci,1994,34:1143-1147.
[20] ZHOU X L,CARTER T E,CUI Z L,et al. Genetic base of Japanese soybean cultivars released during 1950 to 1988[J].Crop Sci,2000,40:1794-1802.
