高通量测序在酱香白酒微生态多样性研究中的应用
2017-06-01郭敏黄永光邱树毅胡建峰胡峰钟方达
郭敏,黄永光*,邱树毅,胡建峰,胡峰,钟方达
(1.贵州大学酿酒与食品工程学院,贵州贵阳550025;2.贵州大学贵州省发酵工程与生物制药重点实验室,贵州贵阳550025;3.贵州茅台酒厂(集团)习酒有限责任公司,贵州习水564622)
高通量测序在酱香白酒微生态多样性研究中的应用
郭敏1,2,黄永光1,2*,邱树毅1,2,胡建峰3,胡峰3,钟方达3
(1.贵州大学酿酒与食品工程学院,贵州贵阳550025;2.贵州大学贵州省发酵工程与生物制药重点实验室,贵州贵阳550025;3.贵州茅台酒厂(集团)习酒有限责任公司,贵州习水564622)
通过提取酱香型白酒生产发酵过程3轮次、7轮次酒醅微生物基因组DNA,采用高通量测序技术对酒醅中的微生态多样性进行测序分析。结果表明,酱香型白酒发酵过程3轮次酒醅(L3)中乳杆菌属(Lactobacillus)和芽孢杆菌属(Bacillus)是原核微生物中绝对优势菌,而7轮次酒醅(L7)中盐单胞菌属(Halomonas)是原核微生物中最优势菌属;同时在L3样品中嗜热真菌属(Thermomyces)和嗜热子囊菌属(Thermoascus)是发酵过程真核微生物中的绝对优势菌,而在L7样品中隐球菌属(Cryptococcus)则是真核微生物中的最优势菌属。对比两轮次酒醅中微生物分析结果,二者的微生态多样性存在较大的差异。分析认为可能正是因为两轮次酒醅中微生物在发酵过程中处于动态变化的差异性,导致两个轮次生产出的酒在产量和质量风味上自然存在较大的差异。
酱香型白酒;酒醅;高通量测序方法;微生物多样性
GUO Min1,2,HUANG Yongguang1,2*,QIU Shuyi1,2,HU Jianfeng3,HU Feng3,ZHONG Fangda3
(1.School of Liquor and Food Engineering,Guizhou University,Guiyang 550025,China;2.Key Laboratory of Fermentation Engineering and Biological Pharmacy of Guizhou Province,Guizhou University,Guiyang 550025,China; 3.Guizhou Moutai Brewery(Group)Xijiu Co.,Ltd.,Xishui 564622,China)
酱香型白酒是中国最具有代表性、独特性的蒸馏酒,以其独特的酿造工艺形成独特的风格体系;以酱香突出、酒体醇厚、空杯留香等风格而闻名[1]。酱香型白酒生产过程发酵酒醅在堆积、窖池发酵过程富集、培养了大量微生物,微生物在酒醅中生长繁殖并产生大量代谢物和风味化合物。在酱香型白酒生产中,第3、4、5轮次生产酒酱香风格典型突出,酒体较为醇厚、丰满,产量和质量效益优势明显;而第7轮次酒的生产属于酱香型白酒酿造工艺的最后一轮,发酵底物的发酵特性处于最差状态,微生物的群落结构也明显与3、4、5轮次存在明显差异,所生产基酒的风格、质量、出酒率也都相差较大。发酵酒醅中的微生物结构直接决定着酒的产质量与酒体风味,酿酒微生物主要包括细菌、真菌等,真菌包括霉菌、酵母菌等。细菌在发酵过程主要功能作用是分泌风味型酶、代谢产生风味物质、调节发酵过程微生态结构等。霉菌主要分泌糖化酶、蛋白酶等,将淀粉转化为可发酵性的糖、分解蛋白质,提高淀粉、蛋白质的利用率。酵母菌主要包括了酒精酵母、产酯酵母等,酒精酵母主要起代谢产酒精的作用,产酯酵母具有产酯能力,能够增加酒体的酯含量,赋予酒舒适的香气。因此,研究酱香型白酒发酵过程的微生态多样性及其结构对诠释酱香型白酒独特风味的形成具有重要的意义。
高通量测序技术(high-throughput sequencing)可以满足对多个样品中的微生物进行深入分析,其主要特点是通量高、速度快、准确度高、实时检测、数据信息量大等,能在很短时间内获取大量的数据[2-3]。目前,国内应用高通量测序技术分析研究浓香型、清香型白酒发酵酒醅中细菌或真菌微生态多样性已有报道[4-7],且高通量测序技术也被用于研究不同发酵食品中的微生物生态,包括醋[8]和牛奶[9],而在酱香型白酒酿造领域还很少涉入。
本研究以酱香型白酒酿造的第3轮次和第7轮次的酿造酒醅为样本,探索利用高通量测序分析酒醅中的细菌16SrDNA的V4变异区序列和真菌内的转录间隔区(internal transcribed spacer,ITS)序列,研究酱香型白酒酿造过程酒醅中细菌和真菌的生态结构。旨在从微生态多样性的角度解析导致两个轮次产酒在产量、质量和风格上存在较大差异的原因,建立高通量测序技术应用于酱香型白酒制酒微生态多样性及其结构的研究分析方法,更加准确、完整的解析酱香型白酒酿造过程酒醅中微生物的群落结构及其主导功能,为今后全面研究酱香型白酒酿造微生态多样性及其相关研究提供可行性的方法和奠定基础。
1 材料与方法
1.1 材料与试剂
样品取自茅台集团XJ公司酱香型酒制酒车间2016年第3轮次、第7轮次生产酒醅,取后置于-20℃冰柜保存,备用。
DNA提取试剂盒:美国MP Biomedicals公司;引物由深圳市恒创基因科技有限公司合成。
1.2 仪器与设备
SpeedCycler PCR仪,德国Analytikjena公司;GS junior高通量测序仪,美国Roche公司。
1.3 方法
1.3.1 样品处理方法
称取10 g样品,用15 mL灭菌后的0.1 mol/L磷酸盐缓冲液(phosphate buffer solution,PBS)悬浮,加入适量玻璃珠,漩涡振荡5 min,300 r/min离心5 min,取上清,沉淀用PBS缓冲液重复洗涤3次,离心后收集上清,将所有收集上清混匀。全部上清于9 000 r/min离心3 min,弃去上清,收集细胞沉淀。再用5 mL PBS洗3次,每次于9 000 r/min离心3 min,收集沉淀。
1.3.2 微生物DNA提取
采用试剂盒方法提取DNA,提取过程参照试剂盒说明书进行。
1.3.3 微生物扩增和焦磷酸测序
使用16S rDNA通用引物515F(5'-GTGCCAGCMGCC GCGG-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3')对细菌DNA的V4高变区进行扩增;使用真菌引物ITS1F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和ITS2R(5'-GC TGCGTTCTTCATCGATGC-3')对真菌ITS1F-ITS2R区域进行扩增。
1.3.4 PCR反应体系
TransStartFastpfuDNA聚合酶20μL反应体系:Forward引物(5 μmol/L)0.8 μL;Reverse引物(5 μmol/L)0.8 μL;5×FastPfu Buffer 4 μL;2.5 mmol/L脱氧核糖核苷三磷酸(deoxy-ribonucleoside triphosphate,dNTPs)2 μL;FastPfu Polymerase0.4μL;TemplateDNA10ng;补ddH2O至20μL。
聚合酶链反应(polymerase chain reaction,PCR)扩增程序:细菌为95℃预变性5 min;95℃变性30 s;55℃复性30 s;72℃延伸45 s;共27个循环。最后72℃延伸10 min。真菌为95℃预变性5 min;95℃变性30 s;55℃复性30 s;72℃延伸45 s;共29个循环。最后72℃延伸10 min。
1.3.5 高通量测序与数据分析
采用高通量测序仪进行测序。应用QIIME软件总体分析测序数据后再利用Greengene数据库进行物种注释,进一步确定序列对应微生物的分类学地位。
2 结果与分析
2.1 样品微生物测序指标分析
对酒醅中微生物的DNA提取并PCR产物纯化及定量后上机测序分析,其Shannon、Chao1指数等分析指标见表1。

表1Shannon等指标分析结果Table 1 Analysis results of Shannon and other indexes
由表1可知,在所测定2个样本的细菌中共获得69651条16SrDNA基因序列(其中L3为32 846,L7为36 805),真菌中共获得34 497条ITS基因序列(其中L3为634,L7为12 459)。为了获得样品中微生物多样性信息,根据97%相似性将序列聚类成为操作分类单元(operational taxonomic unit,OUT),OUT数一定程度上说明了样品多样性的程度。细菌的总OUT数为657(其中L3为634,L7为23),真菌的总OUT数为127(其中L3为54,L7为73),说明样品中微生物构成中,原核微生物的复杂程度高于真核微生物。Alpha值可用来估计样品中细菌群落物种的丰度和多样性,包括Chao1、Shannon等分析指数,在相似度97%以上的分类水平上,利用应用软件计算样品的丰度和多样性指数。Chao1指数用来衡量群落丰度的指数,随着值的增大,微生物群落的丰度也相应较高。本研究中,样品的Shannon曲线值分别是0.12和4.27,Chao 1指数值分别是23和669.68,细菌的测序深度为30 000,因此只需对30 000条序列进行测定分析就可以覆盖酒醅中所有细菌。本次研究样品中细菌多样性分析了36 805和32 846条序列;在真菌中,样品的Shannon曲线值分别为3.9和1.47,Chao 1指数值分别为73和64,真菌的测序深度为10000,因此也只需对10000条序列进行测定分析就可以覆盖酒醅中所有真菌,而本次研究真菌多样性分析了12 459和22 038条序列,说明样品的物种多样性和丰富度都较高。
由表1还可知,2个样品中细菌的Chao1指数大小顺序为L3>L7,说明第3轮次堆积酒醅中细菌的群落丰度比第7轮次高;比较2个样品真菌的Chao1指数,结果表明Chao1指数大小为L7>L3,说明第7轮次堆积酒醅中真菌的群落丰度比第3轮次堆积酒醅高,这与生产实际符合。Shannon指数的大小与Chao 1指数的变化一致。测序结果表明酱香型酒醅样品中的生物物种多样性和丰度都比较高,所覆盖的信息量足以真实性的表征酒醅中微生物群落结构的多样性。
2.2 细菌多样性分析
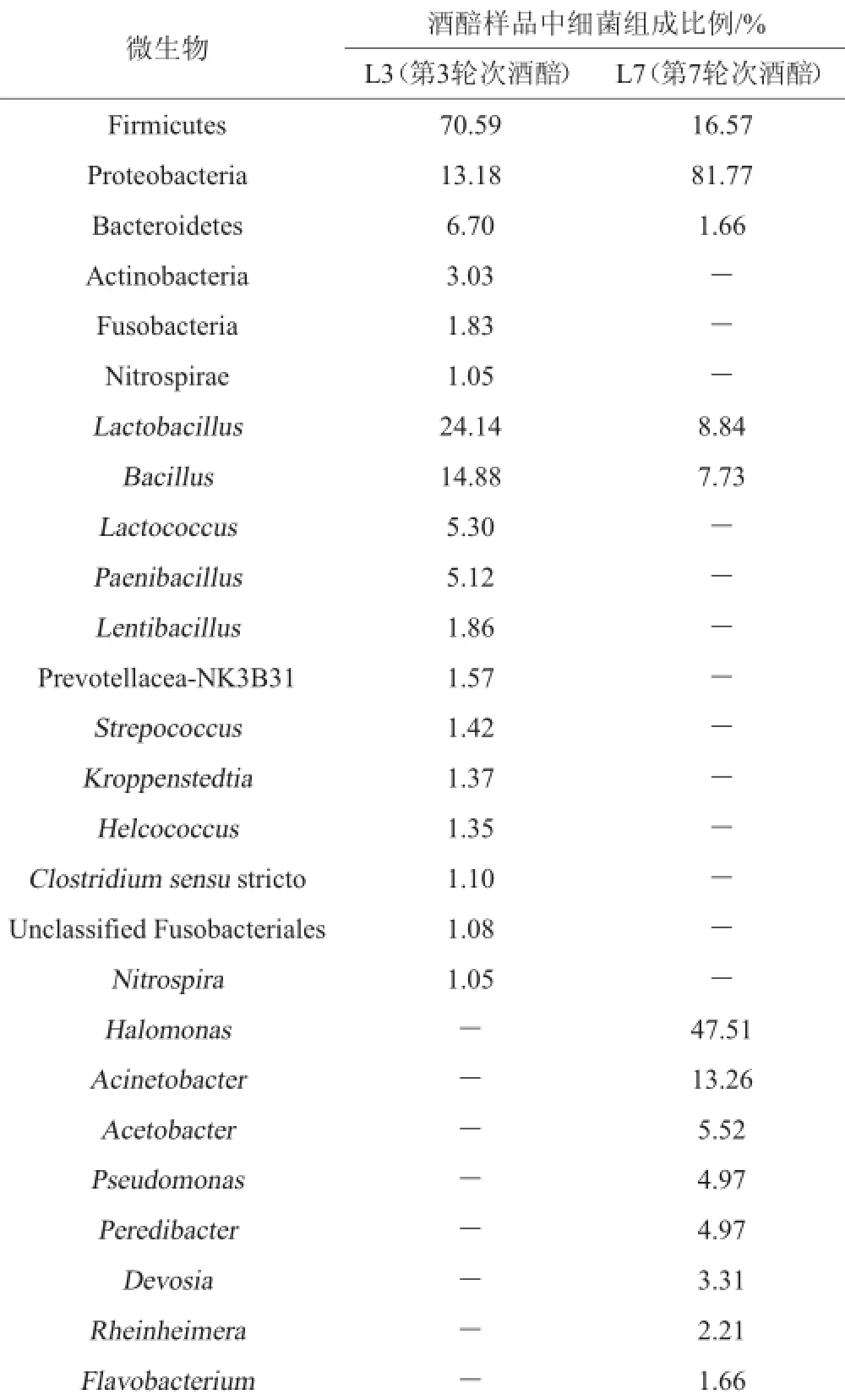
表2 样品中细菌序列的百分含量Table 2 Percentage contents of bacterial sequences in samples
样品中细菌菌群测序数据解析结果见表2。由表2可知,在L3样品中厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、梭杆菌门(Fusobacteria)、硝化螺旋菌门(Nitrospirae)为优势菌门。在属分类水平上,优势菌属分别为乳杆菌属(Lactobacillus)24.14%、芽孢杆菌属(Bacillus)14.88%、乳球菌属(Lactococcus)5.30%、类芽孢杆菌属(Paenibacillus)5.12%、Lentibacillus属1.86%、普雷沃氏菌科-NK3B31属(Prevotellaceae-NK3B31)1.57%、链球菌属(Strepococcus)1.42%、科罗彭斯特菌属(Kroppenstedtia)1.37%、创伤球菌属(Helcococcus)1.35%、狭义的梭菌属(Clostridium sensu strict)1.10%、未分类梭杆菌科(UnclassifiedFusobacteriales)1.08%、硝化螺旋菌属(Nitrospira)1.05%。在L3样品中特有的优势细菌属有:乳球菌属(Lactococcus)、类芽孢杆菌属(Paenibacillus)、Lentibacillus、普雷沃氏菌科-NK3B31属(Prevotellacea-NK3B31)、链球菌属(Strepococcus)、科罗彭斯特菌属(Kroppenstedtia)、创伤球菌属(Helcococcus)、狭义的梭菌属(Clostridium sensu stricto)、未分类梭杆菌科(Unclassified fusobacteriales)、硝化螺旋菌属(Nitrospira)。
由表2还可知,在L7样品中,以厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)为主要优势门。在属的分类水平上,盐单胞菌属(Halomonas)47.51%是最主要的优势属,此外,占总序列的13.26%、8.84%、7.73%、5.52%、4.97%、4.97%、3.31%、2.21%、1.66%的序列分别属于不动杆菌属(Acinetobacter)、乳杆菌属(Lactobacillus)、芽孢杆菌属(Bacillus)、醋酸杆菌属(Acetobacter)、假单胞菌属(Pseudomonas)、吞菌弧菌属(Peredibacter)、戴沃斯菌属(Devosia)、南极适冷菌属(Rheinheimera)、黄杆菌属(Flavobacterium),还包括金黄杆菌属(Chryseobacterium)、土壤杆菌属(Agrobacterium)、纤维弧菌属(Cellvibrio)。在L7样品中特有的优势细菌属有盐单胞菌属(Halomonas)、不动杆菌属(Acinetobacter)、醋酸杆菌属(Acetobacter)、假单胞菌属(Pseudomonas)、吞菌弧菌属(Peredibacter)、戴沃斯菌属(Devosia)、伦黑墨氏菌属(Rheinheimera)、黄杆菌属(Flavobacterium)。其中,盐单胞菌属(Halomonas)为最优势菌属(占总序列的47.51%),该菌属在酿酒领域研究中的报道较少,唐婧[10]利用宏基因组学方法对茅台酒酒曲中的细菌进行多样性分析过程检测到盐单胞菌,该菌为耐盐细菌,部分盐单胞菌可应用于处理高盐染料废水[11]。不动杆菌属(Acinetobacter)为条件致病菌,其耐药性较强,对氨苄青霉素、氯霉素等抗菌药物具有耐药作用[12],该类菌主要为石油烃的降解者,对石油烃具有乳化和降解作用,可以降低石油烃生物毒性[13]。假单胞菌属(Pseudomonas)于条件致病菌[14],该菌类大多可对蛋白质和脂肪进行分解;假单胞菌属与某些高级醇类具有较好的相关性,说明该属的细菌能生成高级醇。假单胞菌属在酿酒领域报道较少,侯小歌等[15]以河南宋河白酒发酵30 d的糟醅为实验材料,通过富集培养及稀释涂布法分离厌氧及兼性厌氧菌,共分离到5株己酸菌、6株丁酸菌、2株甲烷菌、6株乳酸菌。丁酸菌、己酸菌和乳酸菌革兰氏染色均为阳性,甲烷菌为革兰氏阴性菌。丁酸菌和己酸菌分别归梭菌属(Clostridium)和芽孢杆菌属(Bacillus),甲烷菌为假单胞菌属(Pseudomonas),乳酸菌为芽孢乳杆菌属(Sporolactobacillus)。乳杆菌属(Lactobacillus)和芽孢杆菌属(Bacillus)是酱香型白酒发酵过程的优势细菌属,并且乳杆菌属(Lactobacillus)和芽孢杆菌属(Bacillus)在L3样品中为绝对优势细菌,比例分别为24.14%和14.88%,而在L7样品中含量相对较少,分别为8.84%和7.73%,这与第3轮次酒和第7轮次酒的风味差异成正相关,这一群落结构也决定了第3轮次酒和第7轮次酒的本质差异。乳酸杆菌属广泛存在于酒醅中,该类细菌为酒类发酵过程中的生酸微生物,可以通过同型和异型乳酸两种发酵方式产生大量乳酸,并能产生抗菌素等物质,对酿酒过程中致病菌的生长繁殖具有抑制作用,可以促进酵母、霉菌等有益微生物的生长,对酿酒过程的微生态环境具有保护、稳定调控作用[16];芽孢杆菌是酱香型白酒生产过程中的主要产酱香功能菌,具有较强的分泌蛋白酶、淀粉酶和纤维素酶等酶类的能力,可分解大分子物质形成双乙酰、含氮化合物等芳香物质,对酱香型白酒风味物质的形成具有重要贡献作用。多项研究证明芽孢杆菌是酱香风味成分形成的基础和关键,并将其应用于制作强化大曲和堆积过程具有增强白酒酱香风味与提高酱香型白酒质量等作用[17-18]。当前对芽孢杆菌耐高温、产酶、产香的功能性研究较多,不同的芽孢杆菌代谢分泌的酶活力不同且功能特性也不一样[19]。
2.3 真菌多样性分析
酱香型白酒酿造过程真菌类微生物主要有酵母和霉菌。酵母具有产酒和产酯香功能;霉菌主要分泌糖化酶、蛋白酶等,降解淀粉、蛋白质等物质,对产酒具有积极的促进作用。同时,在生长代谢过程还产生一些呈香呈味物质,对酱香酒的风味具有一定的贡献。样品中真菌序列的百分含量分析结果见表3。
由表3可知,在L3样品中子囊菌门(Ascomycota)为优势菌门,占99.84%。在属分类水平上,优势菌属分别为嗜热霉属(Thermomyces)40.73%、嗜热子囊菌属(Thermoascus)25.98%、未分类的散囊菌目(Unclassfied-eurotiales)3.16%、Rasamsonia属4.16%、曲霉属(Aspergillus)2.59%、丝衣霉属(Byssochlamys)2.35%、假丝酵母属(Candida)0.55%、红曲霉属(Monascus)0.17%、毛孢子菌属(Trichosporon)0.07%、小囊菌属(Microascus)0.06%、Trichomonascus属0.03%、青霉属(Penicillium)0.02%、链格孢霉属(Alternaria)0.02%。在L3样品中特有的优势真菌属包括嗜热霉属(Thermomyces)、嗜热子囊菌属(Thermoascus)、丝衣霉属(Byssochlamys)、红曲霉属(Monascus)等。其中,嗜热霉属(Thermomyces)和嗜热子囊菌属(Thermoascus)是酱香型白酒发酵过程绝对优势的真菌微生物且都属于嗜热真菌类,它们是白酒发酵环境中重要酶类的来源,大多能产生具有高活力和热稳定性的纤维素酶、蛋白酶等,对降解高分子多糖和蛋白质具有促进作用,有利于酿酒原料被微生物利用,起到促进微生物繁殖生长以及产酒生香的作用。SHI J H等[20]研究发现,嗜热霉菌属为汾酒曲曲心的优势真菌,且该属真菌在多种不良环境中都具有较好的热稳定性和较高的活力,表明其对酱香型白酒制酒环境的适应性较好。嗜热子囊菌能产生热碱稳定性较好的过氧化氢酶,该酶广泛运用于食品消毒、临床分析、医学诊断以及纺织、造纸、制浆等工业[21]。

表3 样品中真菌序列的百分比含量Table 3 Percentage contents of fungal sequences in samples
在L7样品中主要有两个门类微生物—子囊菌门和担子菌门,分别占49.5%和47.1%。在属分类水平上,隐球酵母属(Cryptococcus)占总序列的48.29%。此外,占总序列13.42%、6.32%、4.17%、2.59%、2.35%、1.8%、1.49%、1.46%的序列分别属于曲霉菌属(Aspergillus)、球果伞属(Stro-bilurus)、假丝酵母属(Candida)、木拉克酵母属(Mrakia)、Kazachstania属、梗孢酵母属(Sterigmatomyces)、锁霉菌属(Itersonilia)、红酵母菌属(Rhodotorula)。还包括链孢霉属(Neurospora)、嗜热子囊菌属(Thermoascus)、嗜热霉属(Thermomyces)、许旺酵母属(Schwanniomyces)、丝衣霉属(Byssochlamys)等。在L7样品中特有的优势真菌属有隐球酵母属(Cryptococcus)、木拉克酵母属(Mrakia)、Kazachstania属、梗孢酵母属(Sterigmatomyces)、锁霉菌属(Itersonilia)、红酵母菌属(Rhodotorula)。其中,隐球酵母属(Cryptococcus)为最优势菌属,关于隐球酵母属在酱香型酒醅中的报道较少,大多报道表明隐球酵母属具有较强的产酯能力[22],并且作为非酿酒酵母对醇类的形成影响也较大,并在酒精发酵中发挥重要作用,具体作用尚未明确,正在对隐球酵母属在酱香型白酒中的作用机制做进一步的研究。
曲霉属(Aspergillus)和假丝酵母属(Candida)是酱香型白酒发酵过程中的优势真菌属,曲霉是酿酒过程中的一大类微生物,是酿造霉菌中种类最多的菌。由于酱香型白酒酿造环境偏酸性,曲霉分泌以酸性水解酶类为主,如酸性淀粉酶、酸性蛋白酶等,这些酶不仅耐乙醇,还具有耐酸特性,使曲霉能在酸性酿造环境条件下正常生长代谢,分解原料中的淀粉与蛋白质,为发酵提供持续性的动力[23]。假丝酵母是白酒酿造发酵过程中的主要产酒酵母,对酱香酒风味形成和感官品质具有重要影响。目前,在多种发酵类食品中均有报道,例如酒曲[24]、奶酪[25]等,该属真菌可以产生大量的酯类化合物,例如乙酸乙酯、乙酸异戊酯等酯类物质[26]等,可为曲坯增加浓郁的酯香,此外假丝酵母还具有较强的产2-丁酮的能力[27]。刘芸雅[28]对黄酒中的假丝酵母属真菌与发酵中有机酸的相关性进行研究,结果表明在白酒发酵过程中该菌类对形成有机酸具有重要作用,并对酒体风格的形成具有重要影响。
3 结论
本研究使用高通量测序方法对酒醅样品中微生物进行了分析研究。结果表明,在酱香型白酒发酵过程中,在第3轮次样品中乳杆菌属(Lactobacillus)和芽孢杆菌属(Bacillus)占有绝对优势,而在第7轮次样品中盐单胞菌属(Halomonas)为最优势菌属;同时在第3轮次样品中,嗜热霉属(Thermomyces)和嗜热子囊菌属(Thermoascus)是发酵过程真菌微生物中的绝对优势菌,而在第7轮次样品中隐球酵母属(Cryptococcus)则为最优势菌属。对比第7轮次和第3轮次酒醅中微生物的分析结果,二者在微生态多样性上存在较大的差别。分析认为可能正是因为第3轮次和第7轮次的酒醅中的微生物在白酒发酵过程中处于动态变化的差异性,导致两个轮次生产出的白酒在产量和风味质量上存在较大的差异。
高通量测序技术与酱香型白酒酿造过程微生物研究的传统方法相比较,由于传统微生物技术的局限性,只能对酒醅中少量的功能微生物进行研究,远不能满足对复杂的微生物群落结构及其多样性研究的需求[20,29];而高通量测序技术从基因组的水平上解析微生物群落结构,对于研究酱香型白酒生产过程中的微生物多样性结构分析更加全面,检测获取的生物信息量更大,更能说明微生物结构的真实性,显示出高通量技术在酱香型白酒生产中对复杂微生物生态结构研究中的明显优势[30-32]。因此该研究为探究白酒酿造微生物群落结构特征提供了一种可借鉴的方法,可为进一步研究酱香型酒醅中功能微生物提供参考。参考文献:
[1]沈怡方.白酒生产技术全书[M].北京:中国轻工业出版社,1998:135-143.
[2]LI R,ZHU H,RUAN J,et al.De novo assembly of human genomes with massivelyparallel short read sequencing[J]. Genom Res,2010,20(2):65-72.
[3]YANG F,ZENG X,NING K,et al.Saliva microbiomes distinguish caries-active from healthy human populations[J]. ISME J,2012,6(1): 1-10.
[4]SUN W N,XIAO H Z,PENG Q,et al.Analysis of bacterial diversity of Chinese Luzhou-flavor liquor brewed in different seasons by Illumina Miseq sequencing[J].Ann Microbiol,2016,66(3):1293-1301.
[5]LIU M K,ZHAO K,TANG Y M,et al.Analysis of clostridium cluster i community diversity in pit mud used in manufacture of Chinese Luzhouflavor liquor[J].Food Sci Biotechnol,2015,24(3):995-1000.
[6]LI X R,MA E B,YAN L Z,et al.Bacterial and fungal diversity in the starter production process of fen liquor,a traditional Chinese liquor[J].J Microbiol,2013,51(4):430-438.
[7]LIANG H P,LI W F,LUO Q C,et al.Analysis of the bacterial community in aged and aging pit mud of Chinese Luzhou-flavour liquor by combined PCR-DGGE and quantitative PCR assay[J].J Sci Food Agr,2014, 95(13):2729-2735.
[8]LI P,LI S,CHENG L L,et al.Analyzing the relation between the microbial diversity of Daqu and the turbidity spoilage of traditional Chinese vinegar[J].Appl Microbiol Biotechnol,2014,98(13):6073-6084.
[9]LIU W J,ZHENG Y,KWOK L Y,et al.High-throughput sequencing for the detection of the bacterial and fungal diversity in Mongolian naturally fermented cow's milk in Russia[J].BMC Microbiol,2015,15(1):45.
[10]唐婧,苏迪,徐小蓉,等.基于宏基因组学的茅台酒酒曲细菌的多样性分析[J].贵州农业科学,2014(11):180-183.
[11]田存萍.盐单胞菌株在高盐染料废水处理中的应用研究[D].大连:大连理工大学,2006.
[12]不动杆菌属、莫拉菌属、金黄杆菌属常规鉴[C].海口:海南省卫生厅医院内感染监测中心,2007:6.
[13]刘玉华,王慧,胡晓珂.不动杆菌属(Acinetobacter)细菌降解石油烃的研究进展[J].微生物学通报,2016,43(7):1579-1589.
[14]梁伟,李娟.非发酵革兰阴性杆菌临床耐药性分析[J].当代医学,2011,17(29):127-128.
[15]侯小歌,王俊英,李学思,等.宋河白酒发酵糟醅中厌氧及兼性厌氧菌的分离与鉴定[J].周口师范学院学报,2013,30(2):65-69.
[16]LV X C,WENG X,ZHANG W,et al.Microbial diversity of traditional fermentation starters for Hong Qu glutinous rice wine as determined by PCR-mediated DGGE[J].Food Control,2012,28(2):426-434.
[17]杨国华,邱树毅,黄永光.酱香白酒生产中产香微生物研究[J].中国酿造,2011,30(4):24-27.
[18]马荣山,刘婷,郭威.麸曲酱香酒醅中酵母菌的分离、筛选及应用[J].中国酿造,2008,27(1):17-18.
[19]李习.高产α-淀粉酶地衣芽孢杆菌的筛选及重组质粒的构建[D].武汉:湖北工业大学,2014.
[20]SHI J H,XIAO Y P,LI X R,et al.Analyses of microbial consortia in the starter of Fen liquor[J].Lett Appl Microbiol,2009,48(4):478-485.
[21]方芳,李寅,堵国成,等.一株嗜热子囊菌产生的碱性耐热过氧化氢酶及其应用潜力[J].生物工程学报,2004,20(3):423-428.
[22]相光明,赵祥颖,刘建军.高产油脂隐球酵母菌发酵条件研究[J].粮食与油脂,2009(3):17-20.
[23]孙剑秋,刘雯雯,臧威,等.酱香型白酒酒醅中霉菌群落组成与功能酶活性[J].中国食品学报,2013,13(8):239-247.
[24]王海燕.PCR-DGGE技术对清香型汾酒微生物群落结构演变规律的研究[D].无锡:江南大学,2014.
[25]GOLIC N,CADEZ N,TERZIC-VIDOJEVIC A,et al.Evaluation of lactic acid bacteria and yeast diversity in traditional white pickled and fresh soft cheeses from the mountain regions of Serbia and lowland regions of Croatia[J]. Int J Food Microbiol,2013,166(2):294-300.
[26]王晓丹,庞博,陈孟强,等.酱香白酒酒醅中产香酵母分离与鉴定[J].食品安全质量检测学报,2014,5(6):42-46.
[27]GUESSAN N,FLORENT K,DRI N,et al.Saccharomyces cerevisiae andCandida tropicalisas starter cultures for the alcoholic fermentation of tchapalo,a traditional sorghum beer[J].World J Microbiol Biotechnol,2010,26(4):693-699.
[28]刘芸雅.绍兴黄酒发酵中微生物群落结构及其对风味物质影响研究[D].无锡:江南大学,2015.
[29]黄永光,徐岩.酱香白酒酿造环境曲霉的分离及Aspergillums hennebergii酶分泌胁迫条件研究[J].食品与生物技术学报,2015,32(8):15-22.
[30]HUANG Y G,WU Q,XU Y.Isolation and identification of a blackAspergillumsstrain and the effect of its novel protease on the aroma of Moutai-flavoured liquor[J].J I Brewing,2014,120(3):268-276.
[31]邓杰.基于高通量测序的浓香型白酒窖泥微生物群落结构研究[D].自贡:四川理工学院,2015.
[32]雷振河.采用高通量测序技术分析清香型白酒酿造微生物[J].食品与发酵工业,2015,41(9):164-167.
TS261.1
0254-5071(2017)05-0146-06
10.11882/j.issn.0254-5071.2017.05.031
2017-02-09
贵州省工业攻关项目(黔科合GZ字[2011]3015);贵州省科学技术厅重大专项(黔科合重大专项字[2012]601-5,黔科合重大专项字[2015]6012);校立项目(研理工2017020)
郭敏(1992-),女,硕士研究生,研究方向为酱香型白酒微生物。
*通讯作者:黄永光(1976-),男,研究员,博士,研究方向为酱香型白酒。
