尼罗罗非鱼P2X4R基因克隆及原核表达分析
2017-05-30余艳玲张永德潘传燕冯鹏霏罗洪林
余艳玲 张永德 潘传燕 冯鹏霏 罗洪林
摘要:【目的】克隆尼罗罗非鱼P2X4R基因,并构建原核表达载体进行诱导表达,为深入研究P2X4R在鱼类中的生物学功能打下基础。【方法】利用PCR克隆尼罗罗非鱼P2X4R基因的3个片段(Gl、G2和G3),拼接获得目的基因后连接pCold Ⅱ载体构建pCold Ⅱ-P2X4R重组质粒,再转化大肠杆菌BL21(DE3)感受态细胞,以IPTG进行诱导表达。分别采用SDS-PAGE和HWestem blotting检测分析重组蛋白P2X4R的表达情况,并运用生物信息学在线分析软件对其理化性质、糖基化位点、跨膜区域、亚细胞定位及信号肽等进行预测分析。【结果】克隆获得的尼罗罗非P2X4R基因大小为1108 bp,与pCold Ⅱ载体重组后转化BL21(DE3)感受态细胞获得的原核表达载体经IPTG诱导可表达获得目的蛋白,在IPTG 1.0 mmol/L、37℃诱导4 h的条件下重组蛋白表达量高于在IPTG 0.1 mmol/L、16℃过夜诱导的表达量。重组蛋白P2X4R的分子量约43.0 kD,其氨基酸数量为354个,理论等电点(pI)为6.78,不稳定指数为37.62,属于稳定蛋白,脂肪族指数为74.35;重组蛋白P2X4R具有3个N-糖基化位点和1个O-糖基位点;该蛋白未见跨膜区,其蛋白几乎100%位于细胞膜内,不含信号肽。【结论】诱导表达获得的尼罗罗非鱼P2X4R蛋白具有3个N-糖基化位点和1个O-糖基化位点,推测其存在糖基化现象,可制备相应抗体用于揭示罗非鱼巨噬细胞的抗原呈递作用机制。
关键词:罗非鱼;P2X4R基因;克隆;原核表达;生物信息学分析
中图分类号:S965.125 文献标志码:A 文章编号:2095-1191(2017)12-2259-07
0引言
【研究意义】嘌呤能P2X受体(P2XR)是由细胞外ATP激活的配体结合非选择性阳离子通道家族成员(Khakh and North,2006),目前已发现有7个亚型,分别为P2X1R~P2X7R(Egan et al.,2004;Weinholdet al.,2010),各亚基结构特征均包含1个大的细胞外环、2个跨膜结构域和2个(氨基和羧基末端)细胞质结构域(North,2002)。经ATP激活后,P2XR在正常和病理条件下可引起不同的细胞反应(Ralevic and Bumstock,1998),且不同P2XR亚型在免疫细胞中均有表达,参与先天免疫调节(Vitiello et al.,2012),其中P2X4R是中枢神经系统中表达量最高的P2XRSIE型。P2X4R基因主要在呼吸道细胞(Nagaoka et al.,2009;Miklavc et a1.,2013)、小胶质细胞(Trang and Salter,2012)、心肌细胞(Yang et al.,2014)、生殖细胞(Gorodeski,2015)、中性白细胞、嗜酸性粒细胞、肥大细胞、T细胞和B淋巴细胞(Di Virgilio and Vu-erich,2015)中表达,因此,明确P2X4R表达分带情况对研究机体免疫及其调控机理具有重要意义。【前人研究进展】已有研究表明,P2X4R在小胶质细胞中可能参与神经性病理疼痛、炎性疼痛(Guo et al.,2005;Tsuda et al.,2005)或癌性疼痛(Gilchrist et al.,2005)调节。在大鼠C6胶质瘤模型中,P2X4R基因在与肿瘤相关的巨噬细胞中上调表达(Guo et al.,2004),]tP2X4R与P2X7R互动激活NLRP3炎性小体(Zechet al.,2016)。P2X4R在控制动物先天性免疫应答过程中也发挥重要作用(de Rivero Vaccari et al.,2012;Kawano et al.,2012b;Chen et al.,2013),具体表现为调节巨噬细胞自噬能力和激活T细胞功能(Burnstock and Boeynaems,2014),但在嗜酸性粒细胞等其他免疫细胞中的功能尚未明确。成功表达P2X4R蛋白是进一步验证其作用机理的基础工作,李琳琳等(2014)成功构建了稳定表达大鼠P2X4R的HEK293细胞系;Zech等(2016)研究发现,P2RX4缺乏可缓解支气管肺泡嗜酸性粒细胞增多、支气管周炎、Th2细胞因子产生和支气管高反应性,即P2RX4拮抗剂是治疗过敏性哮喘的新选择;Asatryan等(2017)研究发现,P2X4R可在较低的ATP浓度(<0.1mmol/L)下被激活,激活后有助于脑源性神经营养因子的释放,且其在触觉异常性疼痛和神经性疼痛中的作用已被证实。【本研究切入点】与哺乳动物相比,P2X4R在低等脊椎动物中的研究报道极少,尤其在鱼类免疫学中的作用机理亟待阐明。【拟解决的关键问题】通过克隆尼罗罗非鱼P2X4R基因,并构建其原核表达载体进行诱导表达,以期为深入研究P2X4R在鱼类中的生物学功能打下基础。
1材料与方法
1.1试验材料
供试尼罗罗非鱼取自国家级广西南宁罗非鱼良种场,体重326.3g/尾;大肠杆菌DH5a、BL21(DE3)表达菌株、pCold Ⅱ克隆载体和限制性内切酶购自宝生物工程(大连)有限公司;2xFast Pfu Mix、NovoRecPCR一步定向克隆試剂盒及DL2000 DNAMarker购自Novoprotein公司;AxyPrep质粒DNA小提试剂盒购自康宁生命科学(吴江)有限公司;IPTG购自美国sigma公司;其他试剂均为国产分析纯。
1.2 pCodII-P2X4R质粒载体构建
根据尼罗罗非鱼P2X4R基因序列(GenBank登录号XM 003448554),利用Primer Premier 5.0对目的基因分3个片段(G1、G2和G3)进行引物设计(表1),分别为P2X4R-F/P2X4R-R1、P2X4R-F1/P2X4R-R2和P2X4R-F2/P2X4R-R,对应目的片段大小为100、948和109 bp。引物由生工生物工程(上海)股份有限公司合成。
PCR反应体系50.0μL:2×Fast Pfu Mix 25.0μL,上、下游引物(10 μmol/L)各2.0μL,DNA模板1.0μL,ddH20补足至50.0μL。扩增程序:94℃预变性5 min;94℃20 s,55℃20 s,72℃延伸(片段G1和G3为20 s,片段G2为1 min),进行30个循环;最后72℃延伸10 min。然后将扩增获得的3段PCR产物以摩尔比1:1:1等量混合作为模板,再以P2X4R-F和P2X4R-R为引物进行PCR扩增,将3段基因序列进行拼接,PCR反应体系及扩增程序同上。
利用限制性内切酶Nde Ⅰ与Hind Ⅲ对pCold Ⅱ载体进行双酶切线性化处理,然后将拼接获得的目的基因片段与线性化载体以摩尔比5:1混合,采用NovoRecPCR一步定向克隆试剂盒进行连接。PCR反应体系20.0μL:10xReaction Buffer 2.0μL,P2X4R250 ng,pCold Ⅱ(Nde Ⅰ/Hind Ⅲ)300 ng,Seamless Cloning Enzyme 1.0μL。混匀,37℃下放置20 min后立即转化DH5aα感受态细胞,抽提测序结果正确的阳性克隆即为pCold Ⅱ-P2X4R重组质粒。
1.3 PCR鉴定
挑取转化培养基上的白色单克隆,置于500.0μL含氨苄青霉素(Amp)的LB液体培养基中,37℃下摇床(180 r/min)培养3~4 h后,用于菌液PCR鉴定。PCR反应体系20.0μL:2xFast Taq Master Mix 10.0μL,上游引物P2X4R-F(10μmol/L)和下游引物P2X4R-R(10μmol/L)各1.0μL,DNA模板1.0μL,ddH20 7.0μL。扩增程序:94℃预变性5 min;94℃20 s,55℃20 s,72℃1 min,进行28个循环;最后72℃延伸10 min。扩增产物用1%琼脂糖凝胶电泳进行检测,阳性克隆经扩大培养后送至生工生物工程(上海)股份有限公司测序。
1.4 P2X4R原核表达
1.4.1表达菌株转化采用AxvPrep质粒DNA小提试剂盒从转化的DH5a感受态细胞中提取pColdII-P2X4R重组质粒,取10.0μL抽提产物加入到50.0μL的BL21(DE3)感受态细胞中,冰浴30 min后42℃水浴热激45 s,立即取出置于冰上冷却2 min,加入500.0μL不含抗生素的LB液体培养基,37℃下振荡培养1 h。将上述培养液均匀涂布于含Amp的LB固体培养基上,37℃下倒置培养16 h,挑取长势较好的单菌落接种于LB液体培养基中,37℃下摇床(180r/min)培养3 h。对菌液进行PCR鉴定,并用1%琼脂糖凝胶电泳检测PCR产物,阳性克隆送至生工生物工程(上海)股份有限公司测序。
1.4.2目的蛋白诱导表达 挑取阳性单菌落接种于1.O mL的LB液体培养基(Amp 50 μg/mL)中,37℃下摇床(180 r/min)培养过夜,次日按1%(v/v)比例进行扩大培养,37℃下摇床(180 r/min)培养至OD600=0.8,加入IPTG进行诱导。诱导条件:①IPTG1.0 mmol/L,37℃诱导4 h;②IPTG 0.1 mmol/L,16℃过夜诱导。按上述条件诱导表达后收集菌体,并悬浮于PBS缓冲液中,4℃下超声波破碎,直至溶液透明。12000 r/min离心20 min,分别收集上清液和沉淀,然后进行SDS-PAGE分析及Western blotting检测。
1.5罗非鱼P2X4R蛋白生物信息学分析
诱导表达获得的尼罗罗非鱼P2X4R蛋白序列采用ProtPara(http:∥web.expasy.org/protparam)进行基本理化性质分析,采用NetNGlyc 1.0及NetOGlyc 4.0(http:∥www,expasy.org/tools)对其糖基化位点进行预测,以ProtScale(http:∥web.expasy.org/protparam/)预测其亲/疏水性及稳定性,通过TMHMM(http:∥www.cbs.dtu.dk/services/SignalP)進行蛋白跨膜区和信号肽预测,以TargetP(http:∥www.cbs.dtu.dk/ser-vice/TargetP)进行亚细胞定位,并通过SWISS-MOD-EL(https:∥swissmodel.expasy.org)进行P2X4R蛋白同源性建模。
2结果与分析
2.1尼罗罗非鱼P2X4R基因克隆
尼罗罗非鱼P2X4R基因分3个片段(G1、G2和G3)进行扩增,电泳结果如图1所示。其中,G1、G2和G3片段大小分别为100、948和109 bp,且均为单一的特异性条带,与预期结果一致。将3个基因片段进行PCR拼接扩增,结果得到大小为1108 bp的基因片段(图2),与预期结果一致,说明目的基因拼接效果良好。
2.2 pCold Ⅱ-P2X4R重组质粒的PCR鉴定结果
挑取转化培养基上的6个白色单克隆,经PCR扩增和电泳检测,结果显示,目的基因片段大小1108bp的单一特异性条带(图3),与P2X4R基因的PCR拼接扩增结果一致,说明P2X4R基因已成功连接至pCold Ⅱ载体上。pColdⅡ-P2X4R重组质粒中P2X4R基因的位置如图4所示。
2.3重组P2RX4蛋白的诱导表达情况
采用两种不同诱导条件对重组P2X4R蛋白进行诱导表达,经SDS-PAGE分析和Westem blotting检测发现,两种诱导条件下重组P2X4R蛋白均有表达(图5和图6),重组P2X4R蛋白分子量约43.0 kD,其表达形式均为包涵体,未检测到可溶性蛋白。此外,在IPTG 1.0 mmol/L、37℃诱导4 h的条件下重组蛋白表达量高于在IPTG 0.1 mmol/L、16℃过夜诱导的表达量。
2.4尼罗罗非鱼P2X4R蛋白的生物信息学分析结果
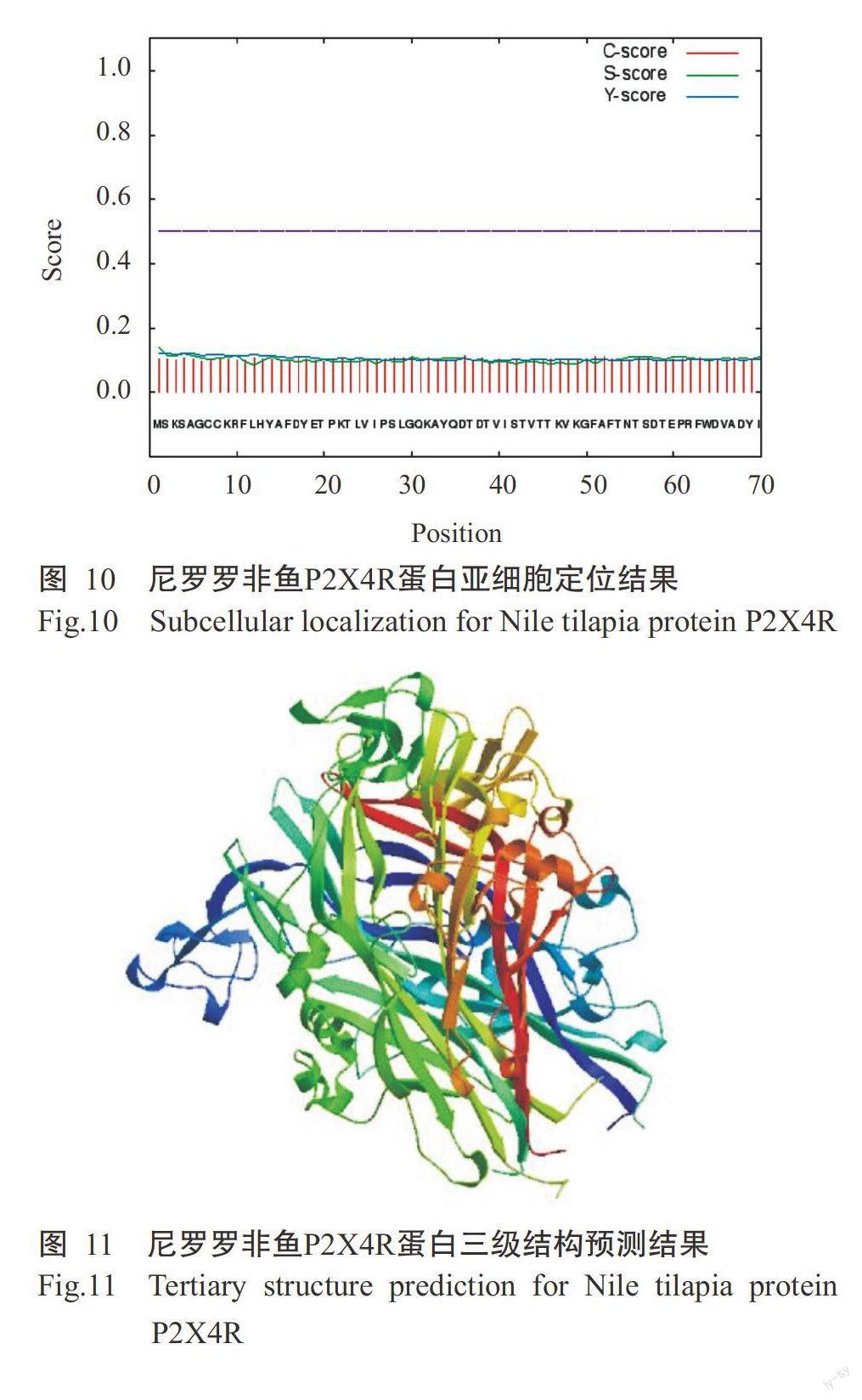
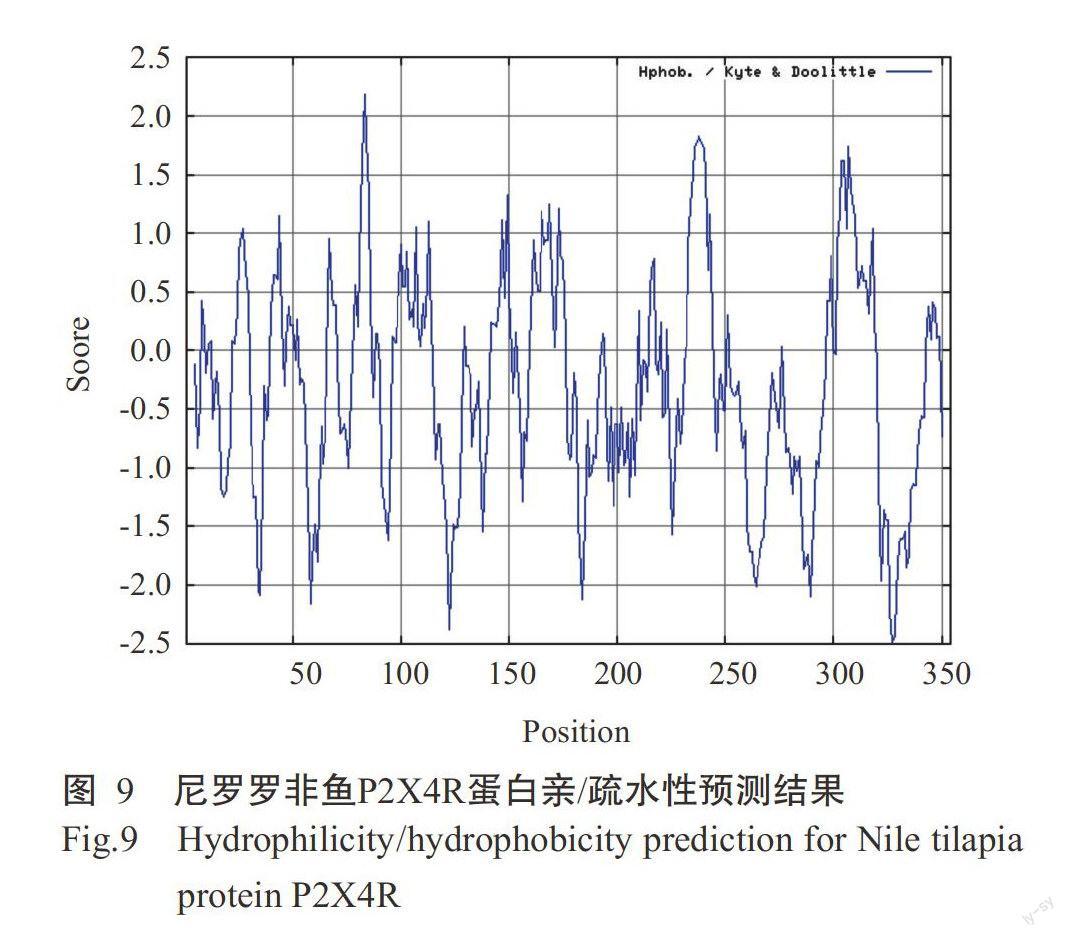
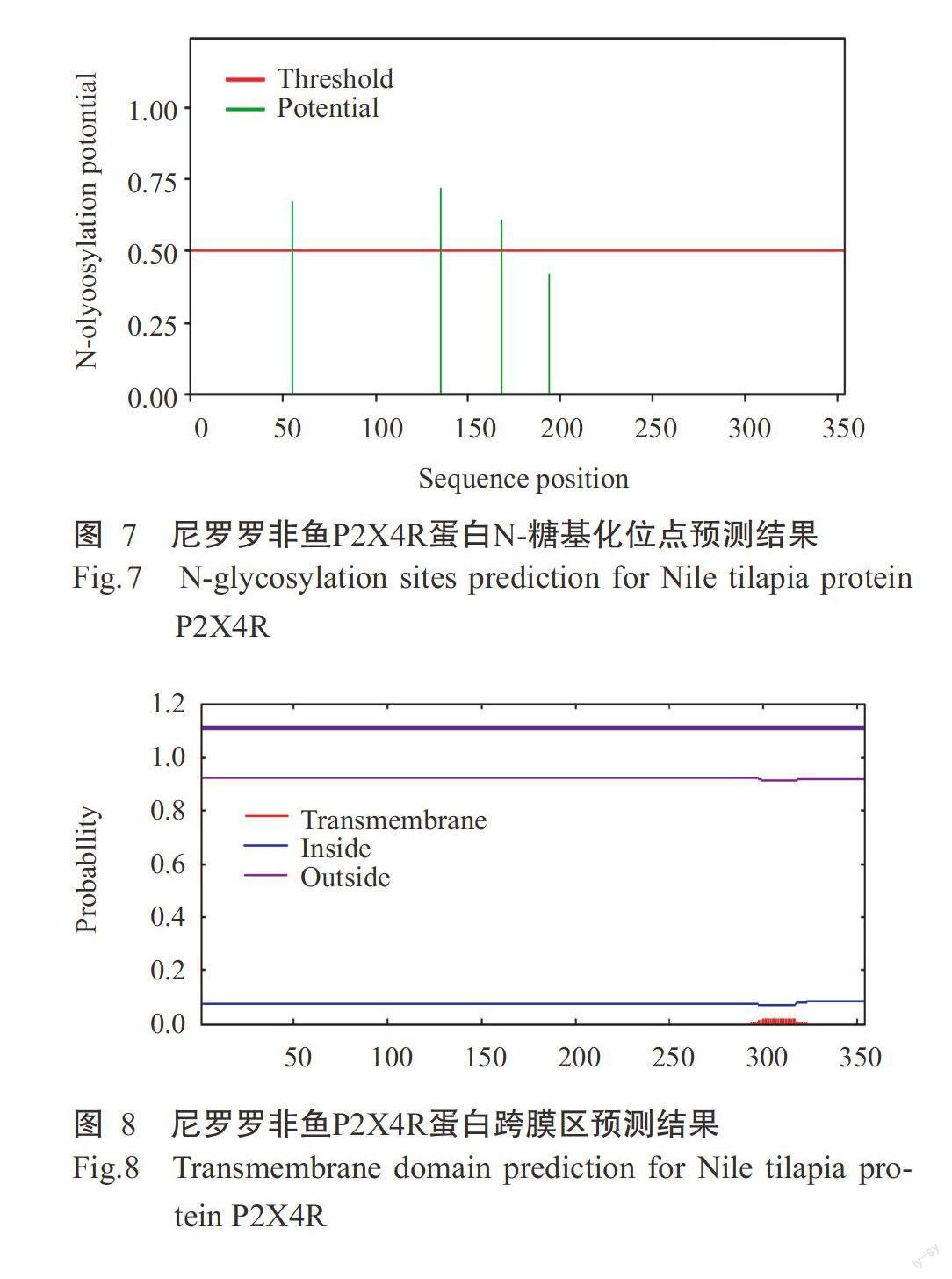
采用ProtPara对诱导表达获得的尼罗罗非鱼P2X4R蛋白进行基本理化性质分析,结果发现该蛋白氨基酸数量为354个,理论等电点(pI)为6.78。尼罗羅非鱼P2X4R蛋白在哺乳动物网织红细胞(体外)的半衰期为30 h,在酵母细胞内的半衰期大于20 h,在大肠杆菌内的半衰期大于10 h;不稳定指数(Ⅱ)为37.62,属于稳定蛋白,脂肪族指数为74.35。糖基化位点预测结果表明,尼罗罗非鱼P2X4R蛋白有3个N-糖基化位点,阈值为0.50(图7),1个O-糖基位点。采用TMHMM对尼罗罗非鱼P2X4R蛋白跨膜区进行预测分析,未发现该蛋白存在跨膜区(图8),其蛋白几乎1 00%位于细胞膜内,不含信号肽。亲/疏水性预测结果(图9)显示,尼罗罗非鱼P2X4R蛋白的平均亲和性(GRAVY)为-0.307,说明该蛋白为亲水性蛋白。亚细胞定位结果发现,尼罗罗非鱼P2X4R蛋白的分泌途径为“-”型(图10),即定位在其他细胞器,无预测剪切位点序列,与TMHMM预测结果一致。利用SWISS-MODEL对尼罗罗非鱼P2X4R蛋白进行三级结构预测,结果如图11所示。
3讨论
P2X4R是不同免疫细胞表达的主要嘌呤能P2X受体亚型之一,在哺乳动物中发挥重要的先天性免疫作用(Coutinho-Silva et al.,2005;de Rivero Vaccari et al.,2012)。与ATP结合后,P2X4R被快速活化,进而导致细胞内钙离子浓度升高、丝裂原活化蛋白激酶(MAPKs)激活和促炎细胞因子产生(Seil et al.,2010;Tsuchiya et al.,2014)。P2X4R还参与NLRP3炎症小体激活(de Rivero Vaccari et al.,2012;Chenet al.,2013)和P2X7R依赖性巨噬细胞凋亡(Kawano et al.,2012a),在微生物感染过程中发挥重要作用(Hung et al.,2013)。2009年,Kawate等首次构建了斑马鱼P2X4R的晶体结构;Li等(2015)通过克隆日本比目鱼(Paralichthys olivaceus)P2X4基因的全长cDNA序列,发现P2X4R是一种膜糖蛋白,可与ATP释放通道Pannexin-1相互作用,主要在日本比目鱼肝胰腺中表达。本研究通过原核载体成功诱导表达获得的尼罗罗非鱼P2X4R蛋白主要以包涵体形式存在,其可溶性蛋白含量较少。原核表达蛋白常以包涵体形式存在,可通过蛋白复性加以解决,但蛋白复性处理程序较繁琐,因此在诱导表达目的蛋白时最好降低甚至消除包涵体的产生,实现可溶性蛋白完全表达。导致原核蛋白表达形成包涵体的原因主要有两种:一种是由于周围环境的影响或缺乏所需要的酶和辅助因子,另一种是在超表达时由于折叠过快,未能及时进行正确的折叠所导致(Clark,2001)。本研究结果表明,通过降低诱导温度和诱导剂浓度可降低P2X4R蛋白包涵体含量,但并未促使可溶性蛋白大量表达,后期仍需进行包涵体复性处理以获得可溶性P2X4R蛋白。
通过对比NCBI数据库中罗非鱼P2X4R蛋白序列与本研究诱导表达获得的尼罗罗非鱼P2X4R蛋白,发现二者问的最大差异在于氨基酸数量和跨膜结构,其跨膜区分析结果表明,参考罗非鱼P2X4R蛋白有5个片段结构,其中2个片段分布在细胞膜上,2个片段为跨膜区,1个片段分布在细胞内;而本研究诱导表达获得的尼罗罗非鱼P2X4R蛋白未见跨膜区,蛋白序列几乎100%位于细胞膜内,推测该蛋白为完整罗非鱼P2X4R蛋白的一部分,且是分布在细胞内的蛋白片段。理论上,诱导表达的尼罗罗非鱼P2X4R蛋白条带应略低于(Western blotting极性处理组)目的条带,但该结果仅在IPTG 0.1 mmol/L、16℃过夜诱导的条件下出现,且出现一条分子量大于目的蛋白的条带,即双条带现象。蛋白糖基化修饰是细胞内质网糖链添加到蛋白质上形成寡糖链的过程,研究表明细胞中有一半以上的蛋白质发生糖基化。糖基化作为一种主要的翻译后修饰方式,对蛋白功能的正常发挥起重要作用,主要调控蛋白质在组织和细胞中的定位、功能、活性、寿命和多样性。Gong等(2002)研究认为,具有糖基化位点的蛋白可通过糖基化位点基因诱变以去除糖基化位点的表达。本研究发现,两种IPTG诱导条件下表达获得的尼罗罗非鱼P2X4R蛋白具有3个N-糖基化位点和1个O-糖基化位点,推测其存在糖基化现象,但尚需通过糖基化染色进一步验证。
4结论
诱导表达获得的尼罗罗非鱼P2X4R蛋白具有3个N-糖基化位点和1个O-糖基化位点,推测其存在糖基化现象,可制备相应抗体用于揭示罗非鱼巨噬细胞的抗原呈递作用机制。
