神经元的细胞周围调制与脑功能
2017-05-30陈直张
陈直张
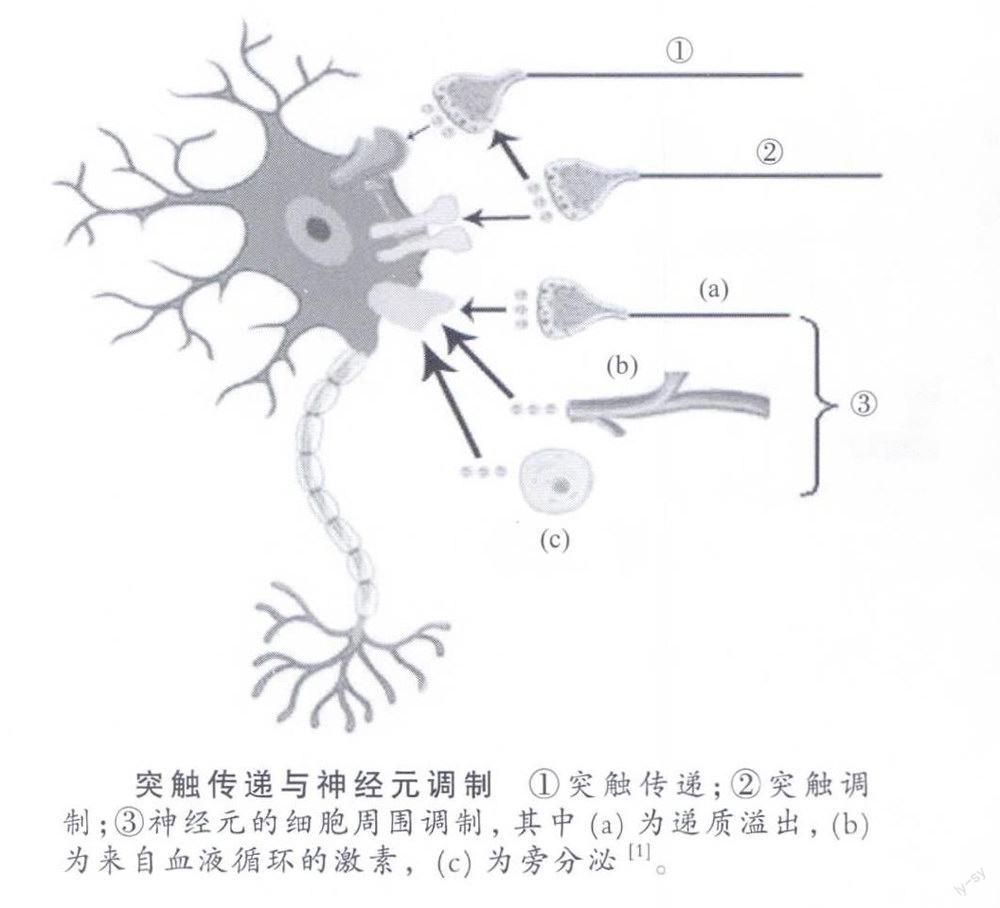

神经元间的信号传输,历来是神经科学一个基础而重要的研究领域。科学家在已经对神经传导和突触传递进行了长期研究,取得了丰富成果的历史基础上,又开始关注神经元的细胞周围调制问题。经科学家研究脑的功能,它可以分为两大类:一个是调节功能,另一个是神智(mind)功能。这两类功能的实现都依靠三种基本的神经活动,即神经传导、神经传递以及神经元的调制。本文着重要讲的是神经元的调制,这里所讲的调制实际是对神经元兴奋性的调制。在人们的日常经验里,神经系统的作用有快有慢。比如说,针刺了身体某处皮肤,人感到疼痛,脑和神经的反应很快;但是如果感到心烦、不舒服,这种作用就属于慢的成分。有一种感受,英文叫作feeling,笔者把它翻译为“内态感”。与感觉和知觉相比,内态感的产生有慢成分的因素在里面,比较难以单纯用神经传导或突触传递来解释。下面就来谈一谈传导和传递以外的神经元调制(modulation ofneurons)问题。
基本神经活动研究发展的不均衡
上面说到三种基本神经活动。简单说来,神经传导是指神经兴奋的信号以动作电位的形式沿着神经元轴突或者长树突传输;突触传递是指神经信号在两个神经元之间,或神经元跟其他可兴奋细胞之间,以电或化学的形式进行传送;神经元调制是指通过突触传递以外的途径,影响神经元兴奋性以及神经信号的传输。
事实上,神经传导和突触传递这两个方面历来是神经科学的中心话题与重点内容,相关的工作很早就开始了。在1791-1797年之间,伽伐尼(L.Galvani)做了一系列实验,刺激青蛙腿的坐骨神经会引起青蛙腿部肌肉收缩。就现在的眼光来看,该实验其实包括了两个成分,一个是神经兴奋的传导,一个是突触传递。以后,关于神经传导的研究占据了神经科学研究的主流,先是有动作电位的细胞外记录,然后有细胞内记录并提出了“离子学说”,后来的膜片钳研究又充实了离子学说。1998年成功解析了钾离子通道的三维结构,最近又有一些关于离子型受体的三维结构研究,如AMPA受体、乙酰胆碱受体等。
突触传递是神经科学研究的又一强劲发展的领域。突触传递的基础在于,每个神经元都是独立单元。最早由卡哈尔(s.Ramon-Cajal)提出,一个一个神经元是各自独立的,这奠定了神经元学说的基础,于是后人才得以引出一个神经元与另一个神经元之间的传递。在研究反射活动的过程中,1906年谢灵顿(c.s.Sherrington)出了一本专业影响巨大的书,叫作《神经系统的整合作用》,书中详细分析了脊髓反射的各种活动规律。更重要的是谢灵顿于1897年提出突触概念,而神经解剖学家在电子显微镜下看见突触是在1950年代。谢灵顿所分析的脊髓反射活动规律,实际上都涉及突触活动。1950年代是突触传递研究蓬勃发展的年代,一方面神經解剖学家研究突触的超微结构,另一方面恰恰是谢灵顿的学生、神经电生理学家埃克尔斯(J.c.Eccles)记录到突触传递的电反应,即兴奋性突触后电位(EPSP)和抑制性突触后电位(IPSP)。以后更深入的研究涉及突触小泡怎样外排,突触后致密(PSD)是一个怎样的结构,突触后的递质受体如何引发突触后反应等。突触传递成为热门的话题。
相对于以上两种神经活动而言,神经科学界对神经元调制这个问题注意得不够。实际上,神经元的调制包括三个方面,即神经传导的调制、突触传递的调制,以及对神经元个别区域或某一状态的调制。在这三方面,对非突触部位的调制,即对一般细胞膜极化状态的调制,也就是对神经元的细胞周围调制(ambientmodulation of neurons)。研究还刚刚开始,正是本文所要讲的重点内容。另外,这里提到对神经元某一状态的调制,什么是神经元的状态?1991-1993年,加拿大一家实验室的研究表明,用细胞内电极记录新纹状体棘神经元(spiny neuron)的细胞内电位可以发现,实际上神经细胞的膜电位是会有波动的,可以一会儿更加去极化,一会儿更加超极化,这样就有“下状态”(downstate,更超极化)和“上状态”(up state,更去极化)之分。由此看来,神经元的膜电位状态不是一成不变的,它可以一会儿趋于上状态,一会儿趋于下状态。这些状态应该也是可以被调制的,不过还没有看到这方面的报道。
神经传导和突触传递的调制
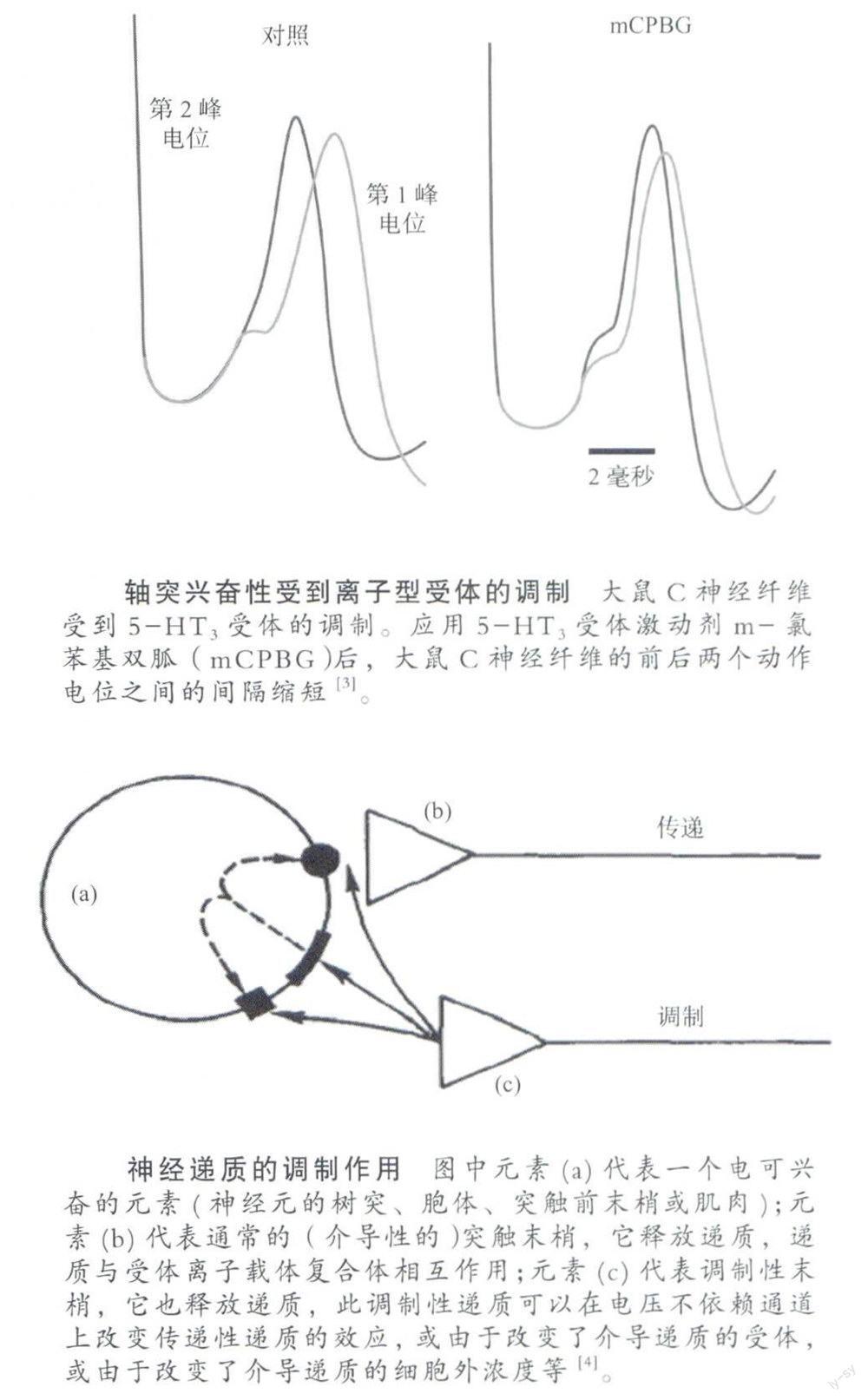
可能有人会认为,神经传导是比较稳定的,不容易发生变动。其实并不尽然,神经传导可以受神经递质调制。实验表明,与对照条件相比,使用γ氨基丁酸(GABA)以后轴突上传导的动作电位显著减少。又有实验表明,应用5-羟色胺(5-HT3)受体拮抗剂以后,轴突上前后两个动作电位之间的间隔缩短了,其原因是轴突本体上有受体,轴突始段也有受体,通过这些受体的作用,神经传导就受到了调制。
最早提出神经调制问题的是1979年库普弗曼(I.Kupfermann)的综述,他认为突触传递过程可以受到调制,而“神经调制”这个词就是与突触传递相对比而提出来的。库普弗曼正确地分析了突触传递与突触传递受调制这两种过程的区别。前者是神经递质的作用,它直接作用于突触后细胞的离子通道;后者通过间接作用,或影响突触后细胞离子通道的功能,或影响突触前的功能,从而影响传递。在这种学术思想基础上,衍生出了有关突触传递如何受到调制的大量研究,这里不作详细介绍。
神经元的细胞周围调制
本文着重要介绍的是神经元调制的第三种,即“神经元的细胞周围调制”,也可以说是“神经元状态的调制”。根据目前的认识,在以下三种情况下可发生神经元的细胞周围调制,或者说影响细胞膜的膜电位:一是较远处突触部位释放递质,递质的溢出(spillover)可以作用到受影响神经元的细胞周围膜上;二是来自血液循环的激素影响神经元的细胞周围膜;三是旁分泌,即邻近细胞分泌的活性物质作用于受影响神经元的细胞周围膜上。下面分别加以介绍。
由GABA溢出所产生的张力性抑制
小脑颗粒细胞上的GABA。受体是获得GABA张力性作用的一个好实验标本。当刺激突触前神经元时有递质(GABA)释放,GABA作用于突触后GABAA受体并把它激活,于是产生抑制性突触后电位(IPSP)。但是如果刺激的频率很高,释放的GABA很多,所释放GABA除激活直接的突触后区GABA。受体以外,还会激活邻近的、突触后膜区以外的受体,这种受体叫突触外受体(extrasyanptic receptor)。在激活突触外受体的条件下,IPSC相对于原来的记录曲线,幅度增大,时间拉长。所增加的这一部分就是张力性抑制。张力性抑制往往只发生在递质释放比较多的情况下。如果把激活时间再拉长一点,张力性抑制会更明显,递质溢出所产生的效应会更远一些,甚至与原来突触距离较远,这就称为细胞周围抑制。
实现张力性抑制的突触外GABA。受体的组构已经清楚,这种组构跟受体的亚单位组成有关。一般位相性受体的亚单位以α、β、γ为多,但实现张力性抑制的受体都含有8亚单位(不过有一种特殊的张力性受体不含δ亚单位)。因此,张力性GABA。受体所介导的电流比较慢,而且一下子也不是很强。现在可以看到,这种调制并不是由G蛋白偶联受体(GPCR)所介导,而是由受体离子通道介导。这里就涉及一个问题:一般认为突触传递通过递质门控离子通道,而神经调制通过GPCR,即使在2013年的科学文献中也是这样写的。其实在笔者看来,此认识并不全面。问题的关键是,在本来通常的突触定义中,只考虑了位相性的、快的作用,但是现在已经知道有的“张力性抑制”(tonicinhibition)和“张力性兴奋”(tonic excitation),其作用是神经调制性质的,却又是可以通过递质门控离子通道实现其作用的。
举一个具体例子,如果晚上睡觉不好,吃了半颗安定就睡好了。这个安定作用在哪里呢?是GABAA。受体,但不是通常所说的突触部位的GABAA受体,至少主要不是。起作用的主要是细胞周围GABAA受体。安定主要不是作用于位相性GABAA受体,而是作用于张力性GABAA受体。
以上讲的就是来自突触部位溢出的递质的神经调制作用。GABAA受体的张力性抑制与临床关系密切,例如在疾病方面,GABAA受体与睡眠异常、应激和精神疾病、癫痫、酒精对脑的影响、卒中及其他脑损伤后脑功能的恢复和保护有关;在药物方面,此受体涉及苯二氮卓类(benzodiazepines)、唑吡坦(zolpidem)、加巴喷丁(gabapentin)、噻加宾(tiagabine)、氨己烯酸(vigabatrin)、加波沙朵(gaboxad01)、加奈索酮(ganaxolone)、非那司提(finasteride)、神经甾体、L-655,508、SNAP-5114、NPPB以及其他药物的作用。苯二氮卓类包括前面讲到的安定;还有唑吡坦,也是一种安眠药。这些药物都是作用在GABA的张力性抑制受体上。说到其中详细机制如何,实际上还有待研究。
旁分泌所产生的神经甾体的作用
接下来介绍神经甾体(neural steroid)的旁分泌作用。神经甾体是神经细胞或神经胶质细胞自己产生的甾体。可作用于含δ亚单位的GABAA受体,选择性地增强GABA反应性。神经甾体对某些GABAA受体的激动作用有时比GABA还要强,它可以引起一个GABA电流。研究表明,神经甾体所引起的都是含8亚单位GABAA受体的电流。
来自血液循环的激素
下面讲激素的调制作用,这涉及笔者的本行。我们在1987、1988年的實验中发现,糖皮质激素之一的皮质醇(cortisol,F)能引起豚鼠腹腔神经节神经元的超极化反应。当皮质醇与血清白蛋白(BSA)结合而成为F.BSA之后,此反应依然存在。因为F.BSA是水溶性大分子,不能透过细胞膜,所以F.BSA应该作用于细胞膜的外侧面。另外,这个反应可以被糖皮质激素拮抗剂RU38486所拮抗,说明膜上存在有一定特异性的接受位点,也可以初步称之为糖皮质激素膜受体。这就是糖皮质激素非基因组机制,或快速反应。
回想当年,有关糖皮质激素膜受体的研究做起来很辛苦。100个细胞里只有几个或者十来个有反应,大多数没有反应。这可能是因为,每个细胞上面的受体数量都不太一样。我们的其他实验证明,糖皮质激素膜受体与传统糖皮质激素核受体在与配基(即甾体激素)结合方面有交叉反应,因此有可能利用糖皮质激素核受体的抗原来辨认和检测膜受体的存在。为了说明糖皮质激素膜受体的存在,我们用胶体金标记糖皮质激素核受体的抗原,来寻找神经元质膜上是否有糖皮质激素核受体样的抗原活性。实验结果表明,神经元质膜上出现一个一个的金颗粒点,而且金颗粒的点可以在非突触区出现,这篇文章发表在《生理学报》上。鉴于金颗粒点可以在非突触区出现,我们当时写了这么一句话:“说明它(指受体)可以接受来自血液循环的激素的作用。”当时我们就提出,对于血液循环里面来的这部分激素,不要仅仅考虑它对核受体的作用,还要考虑它的膜效应、它的快速作用。现在回过头来看,膜上的这一部分作用。实际上就是糖皮质激素对神经元的细胞周围调制作用。
最近还看到另外一项关于鸟类雌激素快速作用的研究。鸣禽用叫声去吸引另外一些鸟,比如雄的吸引雌的。从鸟的大概相当于高等动物大脑皮层的神经元位置记录单个神经元的放电,实验中播放同类鸟叫的录音,以此作为对照实验;另设实验组,同样播放鸟叫声,但在脑局部注射雌激素。将对照组与实验组进行对比时发现,给予雌激素后,反应增强了;反之,假如用拮抗剂,反应就减弱;假如把雌激素合成途径阻断,反应也会减弱。
值得重视的神经元细胞周围调制研究
本文想要强调的是,神经元的细胞周围调制的研究只是刚开始,当然这种研究很难做。下面讨论三个问题。
问题一:细胞周围调制的机制
既然神经元有细胞周围调制,那么这种调制是否可以不影响传导也不影响传递,本身就直接引起脑功能变化?也就是说,传导也没有变,传递也没有变,只因为神经元细胞周围的极化状态变了,人的神智就会改变,人的调节功能就会改变。如果情况是这样的,那么对于脑的认识就应当有一个极大的改变;如果不是这样,神经细胞的周围调制就是通过影响神经传导或突触传递,这样才影响了人的神智或调节功能。后一种情况下也要问。神经元细胞周围的极化状态改变又是如何影响神经传导或突触传递的呢?这也是一个大问题。要回答这个问题,可能需要做新的电生理实验设计。例如,需要做神经元局部微域(microdomain)的电记录,测量的范围大概介于整个神经元跟膜片钳之间的尺度,姑且可以叫作中等膜片钳(mesoclamp)。如果膜片钳所用的膜片是1~4微米2的話,中等膜片钳大概需要50~100微米2左右,要测量这片膜片的电位,要测量它对周围膜的电位有什么影响。这是笔者的一个设想,否则怎么考虑此细胞的周围调制是如何起作用的呢?这个实验将来能否做得出来还不知道,但是如果要考虑这个问题,上述设想应该提出。早在2005年,已经有了电压敏感染料(voltagesensitive dye),它当时被应用于比较宏观的研究,如猴子的大脑皮层,大概是几个平方毫米的面积。现在笔者的设想是比它要小两个数量级,然后来测量神经元细胞膜局部相当大范围的膜电位变化,再看它有没有影响传导,有没有影响传递。
问题二:细胞周围调制与突触区和突触外区受体的关系
突触区的受体跟突触外区及细胞周围区的受体,它们是配基门控离子通道,还是G蛋白偶联受体?它们的分布怎样,密度怎样,有何不同?它们的亚单位构成又怎样?这些受体的激活或抑制是不是影响了上状态和下状态?还要考虑的是,GABA的张力性抑制研究已经做了,其他神经递质的张力性抑制又如何?张力性兴奋的情况如何?这些研究都值得深入做下去。
问题三:细胞周围调制与脑整体活动的关系
与细胞周围调制有关系的脑整体活动,最好的例子是全身麻醉和觉醒转入睡眠这两个问题。这两种脑整体活动都和上行网状激活系统的作用减弱有关。那么,当人从清醒转入昏睡时,或是进入全身麻醉状态时,信号又是怎么被阻断的?哪些信号被阻断了?事情显然不仅仅是神经传导和突触传递的中断或者减弱。前面已经介绍过,至少突触外区GABAA受体的张力性激活是起作用的。以全身麻醉的机制而言,以前曾有学说认为,全身麻醉是由于细胞膜的脂质双层受影响甚至受损伤。现在知道,这种学说过时了。有人提出了新的看法,认为全身麻醉的产生至少与两个递质受体、一个离子通道有关,它们是GABAA受体、NMDA受体、2PK通道。现在的问题在于,哪个神经调制系统(神经传导通路)的哪个或哪些神经元上面的这些受体或离子通道发生了改变。特别要研究:这些元素的细胞周围调制是否起了作用,又起了什么样的作用。只有这样,像全身麻醉、觉醒一睡眠之类问题才能得到比较深入的解决。
总之,神经细胞的周围调制值得重视。
