橡胶树不同倍性的气孔性状差异研究
2017-05-30张源源郭涵袁红章王祥军黄肖李媛李维国
张源源 郭涵 袁红章 王祥军 黄肖 李媛 李维国
摘 要 以通过流式细胞倍性检测鉴定出的橡胶树二倍体、三倍体、四倍体植株为研究对象,对不同倍性植株的气孔性状进行观察和统计。结果表明:不同倍性间的橡胶树叶片的气孔长度、气孔宽度、气孔密度均存在极显著差异。随着橡胶树倍性的提高,气孔的长度和宽度均增大,而气孔密度减小。二倍体、三倍体和四倍体的气孔长度范围分别为10.65~15.29、14.04~18.89 、15.15~19.37 μm,气孔宽度范围分别为7.13~10.65、8.58~11.83、10.38~13.83 μm,气孔密度范围分别为520.62~867.69、402.53~764.80、381.79~555.32个/mm2。以气孔长度15.15 μm、气孔宽度10.38 μm和气孔密度555.32个/mm2为临界值,可以鉴定二、四倍体混合群体中的四倍体。三倍体和二倍体、四倍体在气孔长度值、宽度值和密度值在分布区间上均存在较大的重复,但以气孔长度14.04 μm,气孔宽度8.58 μm和气孔密度764.80个/mm2为临界值,可以筛除二、三、四倍体混合群体中的部分二倍体,提高多倍体鉴定的效率。
关键词 橡胶树;多倍体;气孔性状;倍性鉴定
中图分类号 S722.3+4 文献标识码 A
Abstract The differences of stomatal characters among diploid, triploid and tetraploid were investigated in rubber tree[Hevea brasiliensis Muell. Arg.]. Results showed that there were significant differences in the stomatal length, stomatal width and stomatal density among different ploidy. With the increase of ploidy, the length and width of stomata increased while the stomatal density decreased. The stomatal length ranges of diploids, triploids and tetraploids was 10.65-15.29 μm, 14.04-18.89 μm and 15.15-19.37 μm, respectively. The stomatal width ranges of diploids, triploids and tetraploids was 7.13-10.65 μm, 8.58-11.83 μm and 10.38-13.83 μm, respectively. The stomatal density of diploids, triploids and tetraploids was 520.62-867.69 个/mm2、402.53-764.80 个/mm2 and 381.79-555.32 个/mm2, respectively. The demarcation of stomatal length of 15.15 μm, stomatal width of 10.38 μm and stomatal density of 555.32 個/mm2 could be used to identify tetraploid from mixed population of diploid and tetraploid. The demarcation of stomatal length of 14.04 μm, stomatal width of 8.58 μm and stomatal density of 764.80 个/mm2 could be used to exclude diploid from mixed population of diploid, triploid and tetraploid partly.
Key words rubber tree; polyploid; stomatal character; ploidy identification
doi 10.3969/j.issn.1000-2561.2017.03.001
多倍体植物具有细胞体积大、个体高大、叶片宽厚、分枝数少、花果少、种子大、生长速度快、代谢速率高、特殊代谢物质产量高、抗逆性和适应性强等特点[1-4],在以获得生物产量为目标的生产应用中具有二倍体无可比拟的优势。橡胶树育种的主要目标是获得特殊代谢物质胶乳,通过多倍体育种途径,可以在常规杂交育种的基础上,选育出更高产高抗的橡胶树新品种。在云南大面积推广的云研77-2和77-4被证明为三倍体[5],表明橡胶树多倍体育种作为常规杂交育种的有效补充,具有重要的研究价值。
气孔性状作为多倍体鉴定的指标之一,在杨树[6]、苹果[7]、梨[7]、葡萄[8-9]、月季[10]等多种植物上得到了研究和应用。在橡胶树中,目前已经通过人工诱导[11-21]、自然选择和分离[22-25]等途径获得了大量橡胶树多倍体植株,其中通过形态鉴定、细胞学鉴定、流式细胞倍性检测等方法筛选天然多倍体是最主要的途径[26]。橡胶树不同倍性的气孔性状的相关研究只有周钟毓等[27]对比了人工诱导的四倍体与二倍体对照的差异,关于多倍性水平的气孔性状差异的研究至今未见报道。
本研究以通过流式细胞倍性检测鉴定出的不同倍性的橡胶树为研究对象,分析橡胶树不同倍性的气孔性状差异,探讨通过气孔性状快速鉴定橡胶树倍性的可能性。
1 材料与方法
1.1 材料
试验材料中的二倍体和三倍体植株来源于橡胶树GT1自然授粉子代,四倍体植株来源于人工诱导获得的橡胶树。因资料缺失,四倍体的原始二倍体和亲本未知。所有植株的倍性均经流式细胞倍性检测确认。
1.2 方法
1.2.1 倍性分析 流式细胞倍性检测参考康向阳等[28]的方法,用流式细胞仪(CyFlow Ploidy Analyzer/Cy)进行倍性分析。
1.2.2 气孔长度和宽度测量,气孔密度的观察与计算
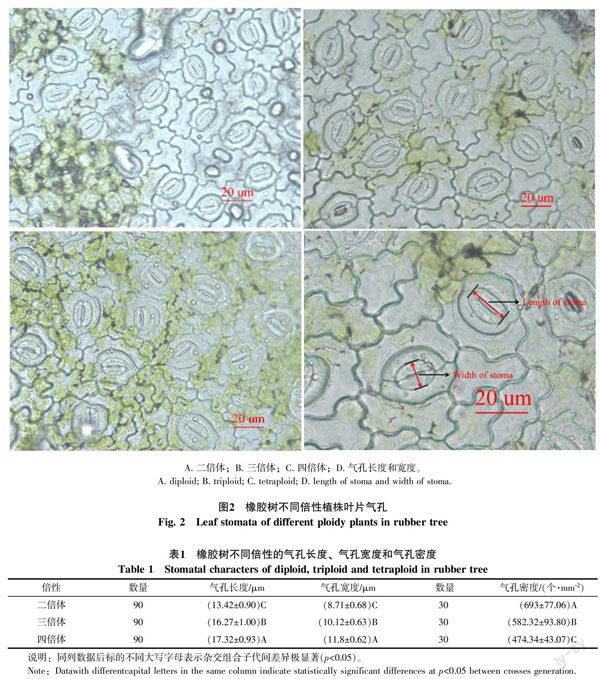
对已检测出的不同倍性植株,二倍体和三倍体分别随机取10株,每株取1个成熟叶片,四倍体植株取10个成熟叶片。采用指甲油法[29],在LeicaDMBL显微镜40×物镜下进行气孔性状观察。每片气孔装片随机选取3个视野使用LeicaDFC550显微镜照相机拍照。每个视野中随机取3个气孔,使用Image J图像分析软件测量气孔长度和宽度,不同倍性分别测量90个气孔。通过一张照片气孔数量和照片所拍摄区域的面积计算气孔密度。测量气孔密度的图像所拍摄区域的面积为0.028 812 mm2。
1.3 数据处理
数据整理在Microsoft Excel 2013中进行,使用IBM SPSS Statistics22中文版进行相关性分析、方差分析和多重比较,使用R语言绘制密度分布曲线。
2 结果与分析
2.1 流式细胞倍性检测
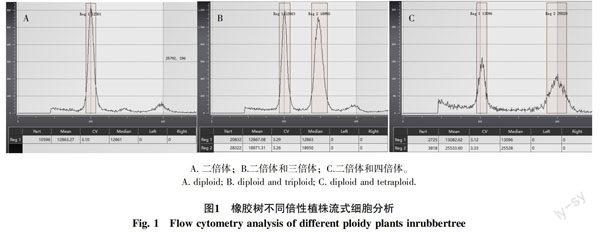
流式细胞倍性检测结果如图1所示,二倍体植株的DNA相对含量为200,三倍体植株的DNA相对含量为300,四倍体植株的DNA相对含量为400。流式细胞倍性检测可以准确鉴定出橡胶树的倍性。
2.2 不同倍性橡胶树叶片气孔性状差异
气孔性状观察结果(图2,表1)表明橡胶树不同倍性间的气孔长度、气孔宽度、气孔密度均存在极显著差异,二倍体、三倍体和四倍体的气孔长度范围分别为10.65~15.29、14.04~18.89、15.15~19.37 μm,气孔宽度范围分别为7.13~10.65、8.58~11.83、10.38~13.83 μm,气孔密度范围分别为520.62~867.69、402.53~764.80、381.79~555.32 个/mm2。表明随着橡胶树倍性的提高,气孔的长度和宽度均增大,而气孔密度减小。分别以每个视野内测量的3个气孔的长度、宽度的平均值为结果,计算气孔密度与气孔长度和宽度的相关性,结果表明气孔密度与气孔长度和宽度存在极显著负相关,相关系数分别为-0.55和-0.507。
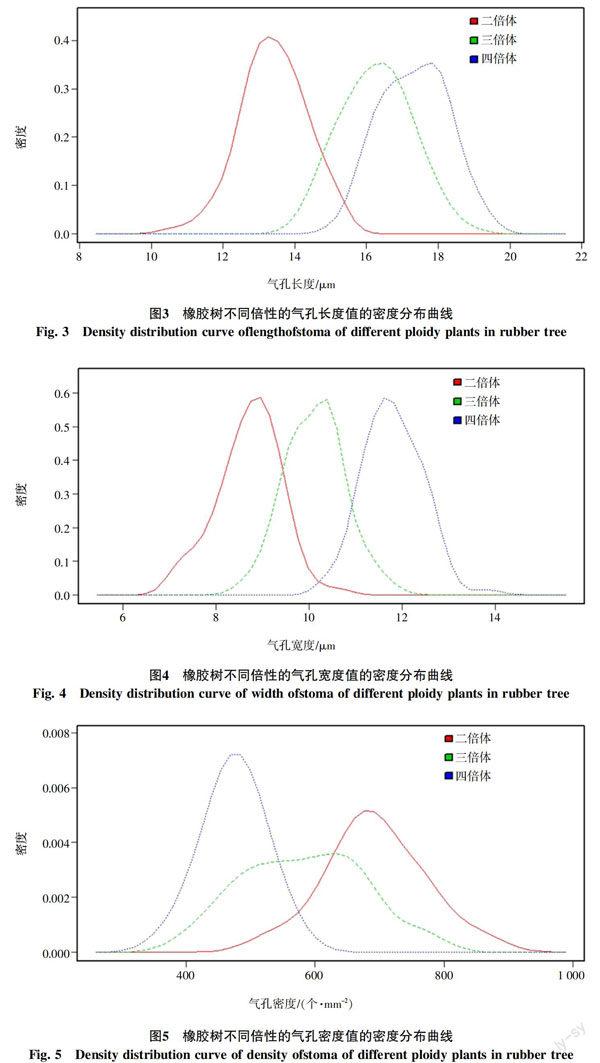
不同倍性橡胶树的气孔性状的密度分布(图3、4、5)表明,四倍体和二倍体在气孔长度值、宽度值和密度值区间上均有较大区别,以气孔长度15.15 μm、气孔宽度10.38 μm和气孔密度555.32 个/mm2为临界值,可以快速准确区分二、四倍体混合群体中的四倍体和二倍体。三倍体和二倍体、四倍体在气孔长度值、宽度值和密度值在分布区间上均存在较大的重复,但以气孔长度14.04 μm,气孔宽度8.58 μm和气孔密度764.80 个/mm2为临界值,可以筛除二、三、四倍体混合群体中的部分二倍体,提高多倍体鉴定的效率。
3 讨论
植物多倍体的气孔密度较二倍体小,气孔长度和宽度较二倍体大,是植物多倍体的重要特征之一。周钟毓等[27]观察经秋水仙碱处理和多次分离筛选与细胞学鉴定认为稳定典型的橡膠树多倍体(四倍体)的气孔,结果表明多倍体植株的气孔密度比二倍体植株小10%左右,气孔的长度、宽度比二倍体植株大10%,与其二倍体对照均存在显著或极显著差异。本研究对比了橡胶树二倍体、三倍体和四倍体的气孔性状差异,结果表明不同倍性间的气孔密度、气孔长度、宽度均存在极显著差异,随着橡胶树倍性的提高,气孔的长度和宽度均增大,而气孔密度减小,与周钟毓的研究结果相似。
植物产生多倍体的机制主要有体细胞染色体加倍和未减数配子(2n)发生两种,体细胞染色体加倍产生四倍体,2n配子发生产生三倍体和少量四倍体。橡胶树四倍体和二倍体的气孔性状差异较大,以气孔长度15.15 μm、气孔宽度10.38 μm和气孔密度555.32个/mm2为临界值,可以快速鉴定人工诱导体细胞加倍得到的二、四倍体混合群体中的四倍体。橡胶树三倍体和二倍体、四倍体在气孔长度值、宽度值和密度值在分布区间上均存在较大的重复,但以气孔长度14.04 μm,气孔宽度8.58 μm和气孔密度764.80个/mm2为临界值,可以筛除2n配子发生途径获得的二、三、四倍体混合群体中部分二倍体,提高多倍体的筛选效率。
植物多倍体的判别和鉴定方法从原理上根据其内在和外在的特征衍生而来[30]。在橡胶树的多倍体鉴定中,形态鉴定法、细胞学鉴定法、流式细胞检测等是最常用的多倍体鉴定方法。
形态鉴定法主要是根据多倍体在整个生长周期内的外部形态特征,初步鉴定是否为多倍体,在大规模的鉴定中可以进行初步筛选,减少工作量。形态鉴定法在橡胶树的多倍体(四倍体)鉴定中被大量应用。橡胶树四倍体的主要特征有:叶片较二倍体对照显著增厚,叶脉增粗一半以上,叶色浓绿,叶片与主动脉夹角变小[16];古铜期叶普遍表现出叶形指数小,侧脉条数少,叶脉检定角小[17];雌花和雄花比二倍体对照的大[31]。本研究中的三倍体和二倍体在形态特征上没有观察到明显区别,因此形态鉴定法在鉴定三倍体中并不适用。
细胞学鉴定法主要是指染色体计数法。橡胶树二倍体染色体条数为2n=36,三倍体为2n=54,四倍体2n=72。经过初步的形态鉴定后,采用染色体计数确认是常用的方法[5,14,31-37]。染色体计数法是倍性鉴定最直接、准确的方法,但其制片技术要求高,对部分植物有一定难度[38]。随着技术的进步,染色体计数法逐渐被更加高效、便捷的流式细胞仪检测方法替代。
流式细胞仪的工作原理是用染色剂对细胞进行染色后测定样品荧光强度,荧光强度与DNA含量成正比,DNA含量柱形图直接反映出不同倍性的细胞数,近年来在倍性分析中应用越来越广泛。张源源[25]首次通过流式细胞倍性检测法从橡胶树GT1自然授粉群体中检测出三倍体植株,并经染色体计数确认。
橡胶树多倍体较二倍体还有其它性状的差异。如黄有军等[36]观察到海垦2、PR107、RRIM600三个品系多倍体植株叶片主要形态特征为叶形指数、一级侧脉数和主脉导管列数比二倍体的小,栅栏组织、海绵组织和叶片厚度比二倍体的大。多倍体树皮较二倍体对照多列射线密度低,射线细胞口径较大,“网眼”比二倍体的宽、性状多样、不规则、排列较乱,围绕多列射线的乳管或轴向薄壁细胞排列变得混乱。陆永林等[31]测定多倍体花粉粒直径有41.0%超过50 μm,甚至有超过55 μm。赖杭桂等[20]认为直径在30~50 μm的为正常花粉,50 μm以上的为加倍花粉粒。以上性状差异因为准确性不高(叶形指数、一级侧脉数等)或难以直接观察(导管列数,花粉直径)等原因未被应用。
本研究根据不同倍性橡胶树气孔性状差异,提出以气孔长度、气孔宽度和气孔密度作为橡胶树多倍性混合群体中的多倍体的鉴别或提高筛选效率的指标,总结和讨论了橡胶树多倍体鉴定的方法及其应用,在橡胶树多倍体育种研究中具有一定的应用价值。
参考文献
[1] Jackson R C. Evolution and systematic significance of polyploidy[J]. Annu Rev Ecol Syst, 1976(7): 209-234.
[2] Cavalier-Smith T. Nuclear volume control by nucleoskeletal DNA, selection for cell volume and cell growth rate, and the solution of the DNA C-value paradox[J]. J Cell Sci. 1978, 34: 247-278.
[3] 康向阳. 林木多倍体育种研究进展[J]. 北京林业大学学报, 2003, 25(4): 70-74.
[4] 谭德冠, 庄南生, 黄华孙. 林木多倍体诱导的研究进展[J]. 华南热带农业大学学报, 2005, 11(1): 27-30.
[5] 李惠波, 周堂英, 宁连云, 等. 橡胶树新品种云研77-2和 77-4的细胞学鉴定及育种过程[J]. 热带亚热带植物学报, 2009, 17(6): 602-605.
[6] 白凤莹, 曾青青, 康 宁, 等. 毛白杨基因库优树倍性检测及性状对比分析[J]. 北京林业大学学报, 2015, 37(4): 113-119.
[7] 李 赟, 石荫坪, 束怀瑞, 等. 应用气孔性状对苹果与梨的倍性判别分析[J]. 果树科学, 1999, 6(1): 9-13.
[8] 戴洪义, 孙 敏, 商传明, 等. 葡萄的染色体倍性与气孔性状的关系及其判别分析[J]. 葡萄栽培与酿酒, 1990(2): 5-9.
[9] 张俊龙. 葡萄气孔、 花粉等与倍性的关系及倍性判别分析[J]. 甘肃科技, 2005, 21(5): 103-104.
[10] 巢 阳, 周 燕. 通过测量叶片气孔鉴定月季染色体倍性的研究[J]. 北京农学院学报, 2015, 30(3): 79-85.
[11] Luiz O T, Mendes A J. Teixeira. Colchicine induced polyploidy of Hevea rubber plants[J]. Bragantia, 1963, 22(30): 383-392.
[12] Shepherd H, Crisfield S B. Induction of polyploidy in Hevea brasiliensis[J]. Planter's Bulletin, 1969, 104: 248-256.
[13] Ong S H. Chromosome morphology at the pachytene stage in Hevea brasiliensis[J]. A preliminary report proceeding of international rubber conference, 1975(11): 3-12.
[14] 陸永林. 诱导产生三叶橡胶多倍体植株的研究[J]. 遗传, 1980, 2(6): 23-26.
[15] 郑学勤, 曾宪松, 陈向民, 等. 诱导橡胶多倍体与其细胞学研究续报Ⅰ[J]. 热带作物学报, 1980, 1(1): 27-31.
[16] 郑学勤, 曾宪松, 陈向民, 等. 诱导橡胶多倍体与其细胞学研究续报Ⅱ[J]. 热带作物学报, 1981, 2(1): 1-9.
[17] 凌绪柏, 陈广盛, 杨廷元. 提高橡胶树多倍体诱导率的方法研究[J]. 热带作物学报, 1988, 9(2): 1-9.
[18] 张树珍, 郑学勤, 曾宪松, 等. 橡胶新种质多倍体的诱导及其研究[J]. 热带作物科技, 1991(3): 35-37.
[19] 曾宪松, 张树珍, 张银东, 等. 离体培养橡胶树体细胞诱导纯多倍体无性系方法的研究初报[J]. 热带作物学报, 1997, 18(2): 15-20.
[20] 赖杭桂, 许少海, 李临生, 等. 巴西橡胶树性器官加倍技术的研究[J]. 热带作物学报, 1998, 19(4): 15-24.
[21] 赖杭桂, 凌绪柏. 橡胶树花芽不同发育阶段秋水仙素诱导花序变异效果的研究[J]. 热带作物学报, 1999, 20(4): 6-10.
[22] 胡东琼, 梁 茂, 吴云通. 橡胶树自然多倍体的选择与分离的研究初报[J]. 热带作物研究(热带农业科学), 1982, 2(3): 1-5.
[23] 刘阳平. 诱导橡胶树多倍体研究[J]. 福建热作科技, 1983(3): 12-16.
[24] 敖硕昌, 和丽岗, 肖桂秀, 等. 橡胶树高产抗寒材料云研77-2、 云研77-4的选育[J]. 云南热作科技, 1988, 21(2): 3-8.
[25] 张源源. 橡胶树座果规律及三倍体诱导研究[D]. 北京: 北京林业大学, 2013.
[26] 张源源, 康向阳, 李维国. 橡胶树多倍体诱导机制和途径[J]. 热带农业科学, 2014, 34(5): 44-49.
[27] 周钟毓, 詹赛荣, 陈传琴, 等. 巴西橡胶树多倍体植株的气孔特征[J]. 热带作物研究(热带农业科学), 1989, 9(4): 4-8.
[28] 康向阳, 王 君. 杨树多倍体诱导技术研究[M]. 北京: 科学出版社, 2010.
[29] Hamill S D, Smith M K, Dodd W A. In vitro induction of banana autotetraploids by colchicine treatment of micropropagated diploids[J]. Aust. J. Bot., 1992, 40: 887-896.
[30] 王丽艳, 梁国鲁. 植物多倍体的形成途径和鉴定方法[J]. 北方园艺, 2004(1): 61-62.
[31] 陆永林, 李渭廉, 冯雪原. 橡胶树多倍体开花习性及生殖器官形态的初步观察[J]. 热带作物学报, 1984, 5(1): 1-7.
[32] 郑学勤, 曾宪松, 陈向民, 等. 诱导橡胶三倍体新方法的研究[J]. 热带作物学报, 1983, 4(1): 1-4.
[33] 张树珍, 郑学勤, 曾宪松, 等. 橡胶新种质多倍体的诱导及其研究[J]. 热带作物科技, 1991(3): 35-37.
[34] 赖杭桂, 许少海, 李临生, 等. 巴西橡胶树性器官加倍技术的研究[J]. 热带作物学报, 1998, 19(4): 15-24
[35] 黄有军, 邱德勃, 王哲魁, 等. 橡胶树人工多倍体染色倍性研究[J]. 热带作物学报, 2002, 23(4): 7-12.
[36] 黄有军, 邱德勃, 王哲魁, 等. 橡胶树人工多倍体的形态性状与产胶特征[J]. 热带作物学报, 2003, 24(1): 8-12.
[37] 赖杭桂, 陈霞, 徐洪伟, 等. 橡胶树三倍体种质创制及生物学鉴定[J]. 热带作物学报, 2013, 34(6): 1 001-1 006.
[38] 陶抵輝, 李小红, 王利群, 等. 植物染色体倍性鉴定方法研究进展[J]. 生命科学研究, 2009, 13(5): 453-458.
