杨树木质部特异启动子诱导活性分析
2017-05-17李杰雒雅婧张爽杜克久
李杰 雒雅婧 张爽 杜克久


摘要:以融合pMDCesAP-GUS、pJCesAP-GUS、pDMDCesAP-GUS的转基因杨树作为试验材料,分别对其根、茎、叶进行创伤及ABA(脱落酸)激素诱导处理,通过定量检测转基因杨树不同器官的GUS酶活性,分析不同启动子对创伤及ABA激素诱导的响应情况。结果表明,创伤处理转基因杨树12 h时,PMDCesAP杨树根的GUS酶活力达到最高点。ABA激素处理转基因杨树,pBI121在根、叶中的GUS酶活力的趋势是先升高后降低;处理3 h时,GUS酶活力最高;pBI121的茎在ABA激素处理前GUS酶活力是最高的;PMDCesAP的根、茎在ABA激素处理前GUS酶活力处于最大值。
关键词:杨树;木质部特异启动子;创伤诱导;脱落酸诱导;GUS活性
中图分类号:S718.46 文献标志码:A 文章编号:1002—1302(2016)01—0063—05
启动子是直接调控基因特异性表达的重要元件,它驱动基因表达,并决定基因表达的强度、时间和空间等特异性。由于组成型启动子驱动的基因在植物不同组织器官中均有不同程度的表达,应用中逐渐暴露出导致转基因植株较野生型植株生长缓慢、植株变矮、产量下降等问题,因此研究和利用组织器官特异性启动子和诱导型启动子具有非常重要的意义。
笔者所在实验室前期分别从3倍体毛白杨-93(Populustomentosa Carr.-“93”)、加杨-1(Populus canadensis Moench.-“1”)中分离了具有木质部组织特异性的MDCe-sAP启动子和JCesAP启动子。通过生物信息学分析表明,2个启动子序列均含有与损伤防御有关的顺式作用元件,还有大量其他与激素、水分等诱导有关的调控元件。本试验以融合pMDCesAP-GUS、pJCesAP-GUS、pDMDCesAP-GUS的轉基因杨树作为试验材料,分别对其根、茎、叶进行创伤及ABA(脱落酸,abscisic acid)处理,通过定量检测转基因杨树不同器官的GUS酶活性,分析不同启动子对创伤及ABA诱导的响应情况,从而为进一步分析启动子中决定组织特异性及诱导性的调控元件奠定基础。
1材料与方法
1.1试验材料
前期已通过农杆菌介导法将3个笔者所在实验室构建的植物表达载体pMDCesAP-GUS(含MDCesAP启动子-GUS融合基因)、pJCesAP-GUS(含JCesAP启动子-GUS融合基因)、pDMDCesAP-GUS(含2个串联MDCesAP启动子-GUS融合基因)的转基因毛白杨作为试验材料;同时将转入pBll21(含35S启动子-GUS报告基因)的毛白杨作为阳性对照、转入根癌农杆菌(Agrobacterium tumefaciens)AGLI的毛白杨作为阴性对照,未转基因的毛白杨组培苗为CK。
1.2材料处理
创伤处理:无菌水清洗以上杨树组培苗,用滤纸吸干表面水分,用剪刀将根剪成长1.5 cm的小段(除尽根部组织培养基);采用镊子对杨树苗茎段部分造成机械创伤;用剪刀将叶片剪下,剪掉叶脉,留下纯叶片,剪成1 cm×1 cm的小块。每种样品的不同部位分别称质量0.1 g,放人1.5 mL的离心管,25℃光照条件下分别处理0、3、6、12、24 h。
ABA激素诱导处理:将以上杨树苗按照创伤处理的方法进行处理,于1.5 mL离心管中加入3倍体积的1 mmol/L的ABA将其浸泡。
1.3转基因杨树GUS报告基因荧光定量检测
取转基因植株根、茎、叶,每份样品取0.1 g,加入3倍体积的GUS提取缓冲液,研磨成粉末,于12000 r/min、4℃离心10 min,取上清液,加入2 mmol/L的GUS反应底物4-MUG,迅速充分混匀后,用0.2 mol/L Na2CO3终止反应。利用RF-5301PC荧光仪,激发波长365 nm,发射波长455 nm,测定蛋白含量,以1 min水解4-MUG生成1 nmol/L4-MU的酶量为1个酶活力单位(U),以每mg蛋白的酶活力表示GUS活性(U/mg),各试验重复3次,取平均值。
2结果与分析
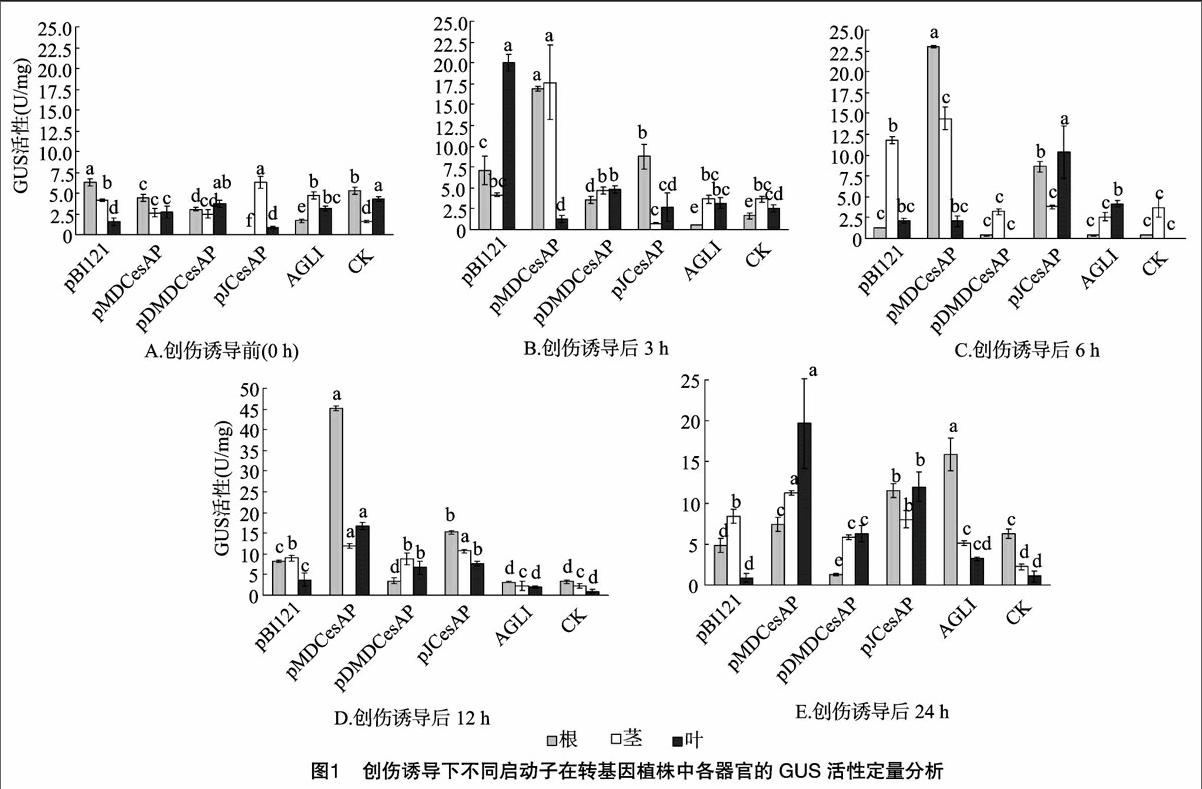
2.1创伤诱导处理转基因杨树的特异性表达分析
2.1.1同一启动子在相同处理时间不同组织器官的酶活性变化 由图1-A可知,创伤处理0 h时,在根中,pMDCesAP、pDMDCesAP和pJCesAP的GUS活性很弱,仅为阳性对照pBll21的70.9%、48.7%和0.7%,pMDCesAP、pDMDCesAP和pJCesAP有显著差异性(P<0.05)。在茎中,pJCesAP的GUS活性很强,比阳性对照pBll21增加了52.2%;pMDCe-sAP、pDMDCesAP的GUS活性很弱,仅为阳性对照pBll21的64.1%和59.0%,且pJCesAP与pMDCesAP和pDMDCesAP有显著差异,pMDCesAP和pDMDCesAP之间的差异不显著。在叶中,pMDCesAP和pDMDCesAP的GUS活性很强,为阳性对照pBll21的1.78倍和2.43倍;pJCesAP的GUS活性很弱,仅为阳性对照pBll21的57.9%;pMDCesAP与pDMDCe-sAP和pJCesAP有显著差异,pJCesAP与阳性对照pBI121之间没有显著差异。
由图1-B可知,创伤处理3 h时,在根中,pMDCesAP和pJCesAP的GUS活性很强,为阳性对照pBI121的2.39倍和1.24倍;pDMDCesAP的活性很弱,仅为阳性对照pBI121的50.9%,pMDCesAP、pDMDCesAP和pJCesAP有显著差异性(P<0.05)。在茎中,pJCesAP的GUS活性很弱,仅为阳性对照pBI121的18.5%,pMDCesAP、pDMDCesAP的活性很强,为阳性对照pBI121的4.22和1.1倍,pMDCesAP与pDMDCe-sAP和pJCesAP有显著差异,pDMDCesAP和pJCesAP与阳性对照pBI121之间无显著差异。在叶中,pMDCesAP、pDMDCe-sAP和pJCesAP的GUS活性均很弱,仅为阳性对照pBll21的6.4%、23.9%和13.2%,DDMDCesAP与DMDCesAP和pJCe-sAP有显著差异,pMDCesAP和pJCesAP之间没有显著差异。
由图1-C可知,创伤处理6 h时,在根中,pMDCesAP和pJCesAP的GUS活性很强,为阳性对照pBll21的17.70倍和6.63倍;pDMDCesAP的活性很弱,仅为阳性对照pBll21的2.9%,pMDCesAP、pDMDCesAP和pJCesAP之间有显著差异性,pDMDCesAP与阳性对照pBI121没有显著差异。在茎中,pMDCesAP的GUS活性很强,为阳性对照pBll21的1.22倍,pDMDCesAP和pJCesAP的活性很弱,仅为阳性对照pBI121的27.4%和32.1%;pMDCesAP、pDMDCesAP和pJCesAP有显著差异性,pDMDCesAP和pJCesAP没有显著差异。在叶中,pJCesAP的GUS活性很強,为阳性对照pBI121的4.91倍,pMDCesAP的GUS活性变化不大,pDMDCesAP的活性很弱,仅为阳性对照pBI121的14.2%;pJCesAP与pMDCesAP和pDMDCesAP有显著差异,pMDCesAP与pJCesAP之间有差异性,与阳性对照pBI121之间差异不明显,pDMDCesAP与pJCesAP有显著差异,与阳性对照pBll21和pMDCesAP没有显著差异。
由图1-D可知,在根中,创伤处理12 h时,pMDCesAP和pJCesAP的GUS活性很强,为阳性对照pBI121的5.53倍和1.86倍,而pDMDCesAP的活性则为阳性对照pBI121的42.8%;pMDCesAP、pDMDCesAP和pJCesAP相互问有显著差异性。在茎中,pMDCesAP、pDMDCesAP和pJCesAP的GUS活性变化不明显;pMDCesAP和pJCesAP之间没有显著差异,但与pDMDCesAP有显著差异;pDMDCesAP与阳性对照pBI121也没有显著差异。在叶中,pMDCesAP、pDMDCesAP和pJCesAP的GUS活性均在增加,为阳性对照pBI121的4.42倍、1.78倍和2.04倍;pDMDCesAP和pJCesAP之间没有显著差异,但均与pMDCesAP有显著差异。
由图1-E可知,在根中,创伤处理24 h时,pMDCesAP和pJCesAP的GUS活性在增加,是阳性对照pBI121的1.52倍和2.37倍;pDMDCesAP的活性在减少,为阳性对照pBI121的26.3%,pMDCesAP、pDMDCesAP和pJCesAP相互问有显著差异性(P<0.05)。在茎中,pMDCesAP的GUS活性比阳性对照pBI121增加了33.4%;pDMDCesAP和pJCesAP的活性在减少,为阳性对照pBI121的69.0%和95.3%;pMDCesAP、pDMDCesAP和pJCesAP相互间有显著差异性,pJCesAP与阳性对照pBll21没有显著差异。在叶中,pMDCesAP、pDMDCe-sAP和pJCesAP的GUS活性很强,是阳性对照pBll21的21.9倍、6.9倍和13.3倍;pMDCesAP、pDMDCesAP和pJCesAP相互间有显著差异性。
2.1.2不同启动子的相同器官随处理时间不同的酶活性变化将各转基因杨树的根部做创伤诱导处理,结果见图2-A。pBll21随处理时间的增加,GUS酶活力在创伤处理6 h时处于低谷;pMDCesAP随处理时间的增加,GUS酶活力呈先增加后降低的趋势,在创伤处理12 h时,GUS酶活力最高,是处理0 h时最低酶活力的10.15倍;pDMDCesAP随处理时间的增加,GUS酶活力总体变化不大,在创伤处理6 h时达到低谷;pJCesAP随处理时间的增加,GUS酶活力整体呈增加趋势,处理12 h时的GUS酶活力达到峰值;AGLI随处理时间的增加,GUS酶活在创伤处理24 h达到峰值;CK随处理时间的增加,GUS酶活力增长不明显,在创伤处理6 h时达到低谷。
将各转基因杨树的茎部做创伤诱导处理,结果见图2一B。pBll21随处理时间的增加,GUS酶活力呈先增加后降低的趋势,在创伤处理3 h时处于峰值,是处理0 h时最低酶活力的2.85倍;pMDCesAP随处理时间的增加,GUS酶活力呈先增加后降低的趋势,在创伤处理6 h时GUS酶活力最高,是处理0 h时最低酶活力的6.7倍;pDMDCesAP随处理时间的增加,在创伤处理12 h时达到峰值;pJCesAP随处理时间的增加,GUS酶活力基本呈先降低后增加的趋势,创伤处理12 h时的GUS酶活力达到峰值;AGLI随处理时间的增加,GUS酶活力呈先降低后增加的趋势,在创伤处理24 h时达到峰值;CK随处理时间的增加,GUS酶活力整体增长不明显,在处理0 h时达到低谷。
将各转基因杨树的叶部做创伤诱导处理,结果见图2-C。pBI121随处理时间的增加,GUS酶活力总体呈先增加后降低的趋势,在创伤处理3 h时达到峰值,是处理24 h时最低酶活力的22.3倍;pMDCesAP随处理时间的增加,GUS酶活力呈先降低后增加的趋势,在创伤处理24 h时GUS酶活力最高,是处理3 h时最低酶活力的15.4倍;pDMDCesAP随处理时间的增加,创伤处理6 h时达到低谷;pJCesAP随处理时间的增加,GUS酶活力基本呈增加趋势,处理24 h时的GUS酶活力达到峰值;AGLI随处理时间的增加,GUS酶活力增长不明显;CK随处理时间的增加,在处理6 h时基因基本处于沉默状态。
2.2 ABA激素诱导各转基因杨树的特异性表达分析
2.2.1同一启动子相同处理时间在不同组织器官的酶活性变化由图3-A可知,ABA诱导处理0 h时,在根中,pMDC-esAP、pDMDCesAP的GUS活性很强,为阳性对照pBI121的4.75倍和2.75倍;pJCesAP的GUS活性很弱,仅为阳性对照pBI121的15.2%,pMDCesAP、pDMDCesAP和pJCesAP相互问有显著差异性(P<0.05)。在茎中,pMDCesAP、pDMDCe-sAP和pJCesAP的活性均很弱,仅为阳性对照pBI121的14.0%、16.1%和9.0%,pMDCesAP与pJCesAP有显著差异,pMDCesAP与pDMDCesAP、pJCesAP之间没有显著差异。在叶中,pMDCesAP、pDMDCesAP和pJCesAP的活性均很弱,仅为阳性对照pBI121的0.2%、55.5%和21.3%,pMDCesAP、pDMDCesAP和pJCesAP相互问有显著差异性。
由图3-B可知,ABA诱导处理3 h时,在根中,pMDCe-sAP和pJCesAP的GUS活性很强,为阳性对照pBI121的1.20倍和2.12倍;pDMDCesAP的活性很弱,仅为阳性对照pBI121的13.1%,pMDCesAP、pDMDCesAP和pJCesAP有显著差异性(P<0.05),pMDCesAP与阳性对照pBI121无显著差异。在茎中,pMDCesAP的GUS活性与阳性对照pBI121的变化不大,pDMDCesAP、pJCesAP的活性很强,为阳性对照pBI121的4.68倍和6.57倍,pMDCesAP、pDMDCesAP和pJCesAP相互间有显著差异(P<0.05),pMDCesAP與阳性对照pBI121之间无显著差异。在叶中,pMDCesAP、pDMDCesAP和pJCesAP的GUS活性均很弱,仅为阳性对照pBI121的10.3%、9.9%和11.7%,pDMDCesAP、pMDCesAP和pJCesAP之间没有显著差异。
由图3-C可知,ABA诱导处理6 h时,在根中,pMDCe-sAP和pJCesAP的GUS活性很强,为阳性对照pBI121的1.22倍和2.23倍;pDMDCesAP的活性与阳性对照pBI121的的活性相差不大,pJCesAP与pMDCesAP、pDMDCesAP之间有显著差异性(P<0.05),pMDCesAP与pDMDCesAP没有显著差异。在茎中,pMDCesAP的GUS活性在减少,为阳性对照pBI121的51.8%,pDMDCesAP和pJCesAP的活性在增加,是阳性对照pBI121的1.43倍和3.51倍,pMDCesAP、pDMDCe-sAP和pJCesAP相互间有显著差异性(P<0.05),pMDCe-sAP、pDMDCesAP与pJCesAP没有显著差异。在叶中,pMDC-esAP、pDMDCesAP和pJCesAP GUS活性均很弱,仅为阳性对照pBI121的15.2%、18.1%和19.7%,pMDCesAP、pDMDCe-sAP、pJCesAP与阳性对照pBI121有显著差异(P<0.05);pMDCesAP、pDMDCesAP和pJCesAP相互间差异不显著。
由图3-D可知,ABA诱导处理12 h时,在根中,pMDCe-sAP和pJCesAP的GUS活性很强,为阳性对照pBll21的3.52倍和5.91倍,而pDMDCesAP的活性仅为阳性对照pBI121的49.8%,pMDCesAP、pDMDCesAP和pJCesAP相互间有显著性差异(P<0.05);pDMDCesAP和pJCesAP之间没有显著差异。在茎中,pMDCesAP、pDMDCesAP和pJCesAP的GUS活性均减少,为阳性对照pBI121的14.0%、31.7%和57.8%,pMDCesAP、pDMDCesAP和pJCesAP相互间有显著差异。在叶中,pJCesAP的活性在增加,是阳性对照pBll21的1.33倍;pMDCesAP和pDMDCesAP的GUS活性在减少,为阳性对照pBI121的4.4%和81.9%,pMDCesAP、pDMDCesAP和pJCesAP相互间有显著差异性(P<0.05)。
由图3-E可知,ABA诱导处理24 h时,在根中,pMDCe-sAP、pDMDCesAP和pJCesAP的GUS活性均在增加,是阳性对照pBI121的2.99倍、3.02倍和1.28倍;pJCesAP与pM-DCesAP和pDMDCesAP之间有显著差异性,pMDCesAP和pDMDCesAP没有显著差异,pJCesAP与阳性对照pBI121之间也没有显著差异。在茎中,pMDCesAP、pDMDCesAP和pJCe-sAP的GUS活性均为增加,是阳性对照pBI121的1.21倍、1.75倍和2.12倍;pMDCesAP、pDMDCesAP和pJCesAP相互间有显著差异性,pMDCesAP与阳性对照pBI121之间没有显著差异。在叶中,pMDCesAP、pDMDCesAP和pJCesAP的GUS活性均很强,是阳性对照pBI121的6.19倍、3.41倍和1.91倍,pMDCesAP、pDMDCesAP和pJCesAP相互间有显著差异性。
2.2.2不同启动子的相同器官随处理时间不同的酶活性变化 将各转基因杨树的根部做ABA诱导处理,结果(图4-A)表明,pBI121随处理时间的增加,GUS酶活力呈先增加后降低的趋势,在ABA处理3 h时GUS酶活力达到高峰,是处于低谷的处理24 h时的6.29倍;pMDCesAP随处理时间的增加,GUS酶活力总体上呈降低的趋势,ABA处理0 h时GUS酶活力最高,是处理24 h时最低酶活力的2.97倍;pDMDCe-sAP随处理时间的增加,GUS酶活力在ABA处理0 h时达到顶峰,处理12 h时达到低谷;pJCesAP随处理时间的增加,GUS酶活力相对于处理0 h时均是增加的,在ABA处理3 h时,GUS酶活力达到峰值,是处理0 h酶活力的46.8倍;AGLI随处理时间的增加,GUS酶活在ABA处理12 h达到峰值;CK随处理时间的增加,GUS酶活力呈先增加后降低的趋势,在处理6 h时达到峰值,处理24 h时到达低谷。
将各转基因杨树的茎部做ABA诱导处理,结果(图4-B)表明,pBI121随处理时间的增加,GUS酶活力呈先降低后增加再降低的趋势,在ABA处理0 h时GUS酶活力达到高峰,处理3、6、24 h时GUS酶活力相差不大且均很低,在处理12 h时GUS酶活力却增加了;pMDCesAP随处理时间的增加,GUS酶活力总体上没有发生大的变化,ABA处理0 h相对较高;pDMDCesAP随处理的增加,GUS酶活力总体上呈先增加后降低的趋势,在ABA处理3 h时达到高峰,是处理6 h的2.98倍;pJCesAP随处理时间的增加,GUS酶活力总体呈先增加后降低的趋势,在ABA处理3 h时达到高峰,是处理0 h时酶活力低谷的5.6倍;AGLI随处理时间的增加,GUS酶活在ABA处理12 h达到峰值;CK随处理时间的增加,GUS酶活力总体呈先降低后增加的趋势,在处理3 h时达到最低谷。
将各转基因杨树的叶部做ABA诱导处理,结果(图4-C)表明,pBI121随处理时间的增加,GUS酶活力呈先增加再降低的趋势,在ABA处理3 h时达到高峰,在处理24 h时处于低谷;pMDCesAP随处理时间的增加,GUS酶活力总体上呈增加的趋势,ABA处理24 h达到高点;pDMDCesAP随处理的增加,GUS酶活力总体上呈先降低后增加的趋势,在ABA处理0 h时达到高峰,是处理3 h时活力低谷的4.3倍;pJCesAP随处理时间的增加,在ABA处理12 h时GUS酶活力达到高峰;AGLI随处理时间的增加,GUS酶活呈先增加后降低的趋势;CK随处理时间的增加,ABA处理3 h时GUS酶活力处于高峰,处理6 h时达到最低谷。
3讨论与结论
笔者所在实验室前期分离的MDCesAP启动子、JCesAP启动子以及经过改造的DMDCesAP复合启动子经功能检测均具有木质部组织特异性。通过生物信息学分析表明,3个启动子序列均含有与损伤防御有关的顺式作用元件,还有大量其他与激素、水分等诱导有关的调控元件。为了研究三者是否同时具有响应伤害及ABA处理的诱导启动子特性,进行了相关试验。结果表明:在创伤处理3 h后,pMDCesAP根部的GUS酶活力均大于阳性对照pBI121根部的GUS酶活力,尤其在处理6 h时,GUS酶活力为阳性对照pBI121的17.7倍;pMDCesAP茎部的GUS酶活力自创伤处理3 h后,与阳性对照pBI121相比也表现出较高的活性;但在创伤处理24 h时,pMDCesAP叶部的GUS酶活力明显升高,并高于根及茎中的GUS酶活力。由此可见,MDCesAP在创伤处理后3—12 h之间,表现出明显的木质部组织特异性及创伤诱导活性。创伤处理下pDMDCesAP的GUS酶活性变化规律不明显。在创伤处理0 h时,pJCesAP在茎中的GUS酶活性很強,比阳性对照pBI121增加了52.2%,但随处理时间的增加,GUS酶活力基本呈先降低后增加的趋势,创伤处理12 h时的GUS酶活力达到峰值,且仅在处理0、3 h表现出木质部组织特异性。另外,3个启动子在创伤处理24 h时,组织特异性表现均不明显,但pMDCesAP、pJCesAP在根、茎、叶中的GUS酶活力均大于阳性对照pBI121相应部位的GUS酶活力。
在ABA处理试验中,处理0~24 h后,pMDCesAP根部的GUS酶活力均大于阳性对照pBI121根部的GUS酶活力,尤其在处理0 h时,GUS酶活力为阳性对照pBI121的4.75倍。由此可见,MDCesAP在ABA处理后,均表现出明显的木质部组织特异性及创伤诱导活性。ABA处理下,pDMDCesAP的GUS酶活性在0 h最强,是阳性对照pBI121的2.75倍,随后呈现下降、增加、下降、增加的规律性变化。ABA处理3~12 h时,pJCesAP在根中的GUS酶活力均高于阳性对照pBI121,在3、6 h时,茎中的GUS酶活力也高于阳性对照pBI121,且表现出木质部组织特异性。另外,3个启动子在创伤处理24 h时,组织特异性表现均不明显,但在根、茎、叶中的GUS酶活力均大于阳性对照pBI121相应部位的GUS酶活力。
植物诱导抗性是新兴的抗病虫害策略,在病虫害防治中有着广阔的应用前景,日益受到人们的关注。组织特异性及诱导型启动子调控抗性基因的表达策略已成为植物抗性基因工程育种研究中的一大热点。但是天然的特异型启动子大多表达水平不高,不能满足人们的多种需要。因此,对现有的天然启动子进行人工改造,利用不同种类启动子中特异性顺式调控元件及增强子序列去构建高活性的、同时受多种因素调控的复合式启动子将是一个十分重要的解决问题途径。本试验结果说明,在启动子中,决定组织特异性相关的顺式作用元件与决定诱导特性的其他顺式作用元件既可以共同协助,又可以互相抑制,从而调节目的基因在特定的时间、空间及一定环境条件下表达,其具体的作用模式有待进一步研究。
