施纳米硒对小麦籽粒硒含量及其品质性状的影响
2017-04-14李韬孙发宇龚盼王安袁林喜尹雪斌
李韬,孙发宇,龚盼,王安,袁林喜,尹雪斌
(1 扬州大学江苏省作物遗传生理国家重点实验室培育点/粮食作物现代产业技术协同创新中心/教育部植物功能基因组学重点实验室/小麦研究中心,江苏扬州 225009;2 苏州硒谷科技有限公司,江苏苏州 215123;3 中国科技大学苏州研究院,江苏苏州 215123)
施纳米硒对小麦籽粒硒含量及其品质性状的影响
李韬1,孙发宇1,龚盼1,王安1,袁林喜2,3,尹雪斌2,3
(1 扬州大学江苏省作物遗传生理国家重点实验室培育点/粮食作物现代产业技术协同创新中心/教育部植物功能基因组学重点实验室/小麦研究中心,江苏扬州 225009;2 苏州硒谷科技有限公司,江苏苏州 215123;3 中国科技大学苏州研究院,江苏苏州 215123)
【目的】研究基施纳米硒肥对不同小麦品种籽粒百粒重、硒含量、硒形态,其他矿质元素以及面粉糊化特性的影响,为利用纳米硒肥进行小麦硒生物强化提供参考。 【方法】选择 110 份小麦品种 (系),在扬州大学农学院网室种植,采用的施肥方式为基施纳米硒肥,设每千克土施 Se 0 (CK)、100 (Se100)、150 (Se150) mg 2 次重复,随机区组设计。小麦成熟期收获籽粒,测定百粒重。利用离子发射光谱 - 原子吸收仪测定了籽粒 Se、Ca、Mg、Cu、Fe、Mn、Zn 及 S 含量。选择高硒处理中硒含量前 10 名的品种,利用 LC-UV-AFS 测定了籽粒中不同形态硒的含量。随机挑选 59 个品种并利用 RVA 仪 (快速粘度分析仪) 测定其面粉糊化特性的特征值。利用MATLAB 和 SPSS 软件进行数据分析。 【结果】基施硒 0、100、150 mg/kg 土,小麦籽粒的百粒重均值分别为3.78、4.11 和 3.70 g,籽粒中总硒含量分别为 2.00、12.46 和 17.35 mg/kg。籽粒中的硒主要以有机态形式存在,以硒蛋氨酸含量最多,其次是硒甲基化半胱氨酸和少量的硒半胱氨酸,无机态硒仅微量存在于极少数品种中。基施硒肥对测定的矿质元素的吸收积累既有协同也有拮抗作用,对小麦面粉的糊化特性没有显著影响。糊化特性取决于基因型,不同品种间存在极显著差异。 【结论】硒肥可以提高小麦籽粒中的硒含量,但增幅因品种而异;硒强化后籽粒中硒主要以有机态形式存在;低浓度硒处理可以提高小麦籽粒的百粒重,且不会影响小麦的面粉糊化特性。
小麦;纳米硒;硒含量;硒形态;矿质元素;糊化特性
临床医疗试验已经证明,硒对人体健康有重要作用[1],人体缺硒会导致约 40 余种疾病[2-5]。中国营养学会推荐正常人体每日硒摄入量为 60~400 μg[6],但是中国人均摄硒量普遍不足[7]。通过提高作物和蔬菜中硒含量来进行食补是高效、安全、经济的补硒方式[8-9]。
植物中的含硒量因植物种类不同而差异较大,一般植物的正常含硒量低于 25 mg/kg,硒积累型植物可高达 15000 mg/kg[10-11]。虽然硒不是植物的必需元素,但却是有益元素[12],施硒不仅能提高植物中硒含量,也能加快植物的生长发育,提高植物的抗氧化能力和对环境的耐受性[13-17]。小麦是最具硒强化潜力的主粮作物之一[18-19],通过小麦的硒强化可以有效提高籽粒中的硒含量。在贫硒地区通过生物强化增加小麦籽粒中 (特别是小麦面粉中) 的硒含量是补硒的有效策略之一[10]。而且,在低硒土壤中增施硒肥可有效改善作物品质。施硒还可提高第一限制性氨基酸 — 赖氨酸含量 15%~16%[20],还能促进植物对硫的吸收[21]。
小麦粉的糊化特性是指小麦加工和面过程中面团的流体力学属性。小麦 RVA 谱 (特征值) 反映了小麦面粉的糊化特性,是衡量小麦面粉品质和食品加工品质的重要指标[22]。在 RVA 特征值中高峰粘度与面条的弹性、韧性和食用品质呈显著正相关,最终黏度与面条的滑爽性呈极显著正相关,与面条弹、韧性呈显著负相关[23-25]。然而小麦施硒能否影响小麦面粉糊化特性,国内外鲜有报道。
目前国内外对作物进行硒强化所施硒肥主要是硒酸钠和亚硒酸钠,大多采用喷施方式进行。纳米硒肥是一种缓释硒肥,根部基施或追施时其施用的安全浓度范围较硒酸钠或亚硒酸钠盐宽。本文主要利用纳米缓释硒肥对不同小麦品种进行强化并研究硒处理对籽粒中硒含量和形态、其他矿质元素含量以及对面粉糊化特性的影响,为利用纳米硒进行小麦硒强化提供部分参考依据,同时筛选具有硒强化潜力的小麦品种。
1 材料和方法
1.1 试验材料及试验设计
所选的材料包含我国的地方品种、育成品种和部分国外引进品种共 110 份。所有材料为春性或半冬性,均为遗传上的纯系。采用盆栽实验。纳米硒肥为苏州硒谷科技有限公司生产,硒含量3000 mg/kg。设 3 个施 Se 水平,即每公斤土施 0 (对照)、100 (Se100)、150 (Se150) mg 硒肥,每水平各重复 2 次,采用完全随机区组设计。播种前将硒肥和土充分混匀装盆,土壤中基础 (对照) 硒含量为0.016 mg/kg。小麦成熟后,以盆为单位进行收获,收获后立即进行脱粒,并将籽粒放在 65℃ 的烘箱中处理 10 个小时后用密封袋保存备用。
1.2 籽粒百粒重及元素含量测定
根据品种的地域来源和品种类型,共挑选了 74个具有代表性的品种,每个品种随机挑选 100 粒种子称重,重复 2 次。
称取 0.5 g 干燥后的籽粒,加入 5 mL 浓硝酸,3 mL水和两滴过氧化氢在微波消解仪上进行消解,消解液最后定容在 50 mL 的容量瓶中,用等离子发射光谱-原子吸收仪测定 Se、Ca、Mg、Cu、Fe、Mn、Zn和 S 含量。
1.3 硒形态及其含量的测定
从 Se150 处理中挑选籽粒总硒含量最高的 10 个品种,称取 0.1~0.2 g 样品于 10 mL 离心管中,加入5 mL Tris-HCl,摇匀,超声波处理 30 min,加入0.2 mL蛋白酶K,50℃ 气浴恒温振荡器中转速为 250 rpm,培养 18 h 后,加入 0.2 mL 蛋白酶 K,继续培养 6 h,加入 0.4 mL 蛋白酶 XIV,37℃,培养 18 h,再将离心管放入高速离心机,4℃,10000 rpm/min,离心30 min。上清液过 0.22 μm 水系膜后,利用液相色谱-原子荧光联用形态分析仪 (LC-UV-AFS) 测定样品中的硒形态。
1.4 RVA 特征值的测定
参照我国不同麦区小麦面粉糊化特性的变异程度[26],从 110 份材料中挑选 59 份典型材料进行 RVA特征值的测定。RVA (Rapid Viscosity Analyser) 参数使用澳大利亚新港公司生产的 Super3 型粘度仪,称3.5 g 面粉加 25 mL 蒸馏水进行测定。
1.5 数据分析
数据分析和作图采用 SPSS16 和 MATLAB 进行。
2 结果
2.1 施硒对小麦籽粒百粒重的影响
CK、Se100 和 Se150 处理小麦百粒重分别为3.78、4.11 和 3.70 g (图 1),Se100 处理小麦籽粒百粒重显著高于对照 (P < 0.001),Se150 处理小麦籽粒百粒重显著低于 Se100 (P < 0.001),但与对照差异不显著。

图1 不同施硒水平小麦籽粒百粒重Fig. 1 Wheat 100-kernel weight under different treatments of nano-selenium fertilizer
2.2 施硒处理对小麦籽粒硒含量的影响
CK、Se100 和 Se150 处理 110 个小麦品种的硒含量平均值分别为 2、12.93 和 17.14 mg/kg,不同施硒处理间差异均达到极显著 (P < 0.001)。对照处理籽粒硒含量的品种间差异不大 (图 2),但施硒处理籽粒硒含量品种间存在极显著差异 (P < 0.001),Se100 处理的硒含量介于 6.42~22.29 mg/kg,Se150 处理的硒含量介于 10.41~30.99 mg/kg,且品种与处理互作差异极显著 (P < 0.001)。说明不同品种籽粒富硒能力存在基因型差异,对施硒的响应也不同。相较对照,Se100 处理硒含量增幅介于 2.63~20.62 mg/kg,Se150 处理较 Se100 处理小麦籽粒硒含量有所上升,增幅最大的品种是 X359,其增加量为 19.63 mg/kg。硒含量与施硒量基本呈线性递增 (图 3),说明继续提高硒肥浓度仍然有提高籽粒硒含量的可能。
2.3 施硒处理小麦籽粒中的硒形态及其含量
在 Se150 处理条件下,筛选籽粒硒含量最高的10 个品种 (系) 进行硒形态的分析,结果表明,籽粒中硒主要以有机态硒的形式存在 (图 4),其中硒代蛋氨酸 (SeMet) 所占总硒含量的比例最高,变幅为66.98% (X359)~94.52% (陕 89150),甲基硒半胱氨酸(MeSeCys) 的含量次之,郑 9023 籽粒中 MeSeCys 比例高达 39%,其次为宁麦资 18,MeSeCys 占总硒量的 32.24%。10 个品种 (系) 中,有 5 个品种 (西麦1376、郑 9023、陕 89150、陕 107-6、X076) 没有发现有硒代胱氨酸 (SeCys) 形态的存在,但 X359 中SeCys 比例高达 27.41%,其次为优选 14 (13.22%),其余 3 个品种 SeCys 所占比例都小于 3%。SeMet、MeSeCys 和 SeCys 这 3 种有机硒的比例总和在 10 份品种中均高达 96.62% 及以上。除了有机硒之外,西麦 1376、郑 9023、合选 198 中也检测到少量无机硒的存在,这 3 个品种 Se4+所占比例分别为 0.3%、0.51% 和 0.36%。仅在西麦 1376 中发现了 Se6+,所占比例为 3.08%。
由于 10 份品种籽粒硒元素主要是以 SeMet 和MeSeCys 的形式为主,为了比较不同品种籽粒间SeMet 和 MeSeCys 是否存在显著差异,对籽粒中这两者的含量进行了方差分析,结果表明籽粒 MeSeCys含量在品种间有着极显著差异 (F = 3.55,P = 0.006),而 SeMet 在品种间差异不显著 (F = 1.08,P = 0.407)。
2.4 硒处理对小麦籽粒中其他元素含量的影响
施硒后籽粒各种元素含量的变化有多样性的特点,表现为不同品种同一元素或同一品种不同元素对施硒响应存在差异。以高硒处理 (Se150) 为例分析了硒处理对其他矿物质元素的影响。在 110 个品种中,籽粒各种元素对硒处理响应趋势 (增加或降低)一致的品种仅有 15 个 (表 1),其中 9 个品种在施硒后籽粒中 7 种元素含量均出现了不同程度的下降,代表品种有 Field-H、扬麦 2、安农 2 等;而扬麦17、宁资 8、罗麦 8 等 6 个品种施硒后籽粒中 7 种元素含量均有不同程度的增加。可见,元素之间的拮抗或协同主要因基因型而异。仅从元素营养角度考虑,施硒后籽粒 7 种元素含量均有所增加的 6 份材料具有较好的硒强化潜力。
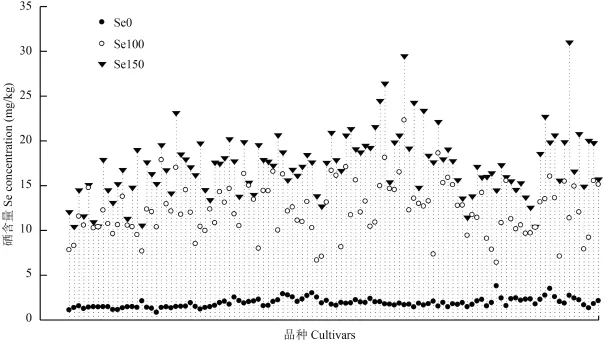
图2 不同施硒水平 110 个品种籽粒硒含量Fig. 2 Grain Se concentrations of 110 cultivars under different selenium addition rates
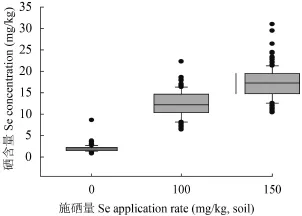
图3 不同硒处理 110 个品种的籽粒硒平均含量Fig. 3 Mean Se concentration in the grains of 110 cultivars under different selenium application rates
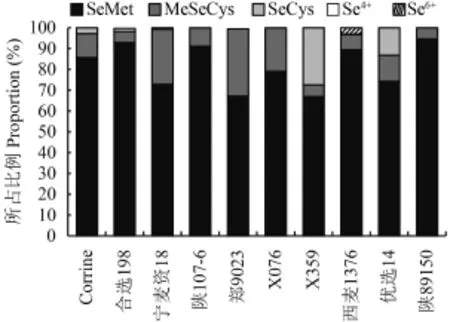
图4 小麦籽粒中不同形态硒所占比例Fig. 4 Proportion of different selenium forms in total selenium in wheat grain
2.5 硒处理对小麦面粉糊化特性 (RVA 参数) 的影响
RVA 各特征值在品种间达极显著差异 (表 2),但在对照与硒处理间 RVA 参数特征值差异都不显著,表明基施纳米硒处理不会影响小麦面粉的糊化特性。
3 讨论
3.1 基施纳米硒对小麦粒重的影响
试验结果表明,与对照相比,Se100 处理的籽粒百粒重显著提高,Se150 处理的籽粒百粒重与对照没有变化,说明基施纳米硒在低浓度水平下可以提高小麦籽粒的百粒重,这与已报道的在小麦[27]和水稻[28]中进行硒强化结果相似。但是随着硒浓度的升高大多数品种百粒重又会出现下降趋势,可能与高浓度硒抑制籽粒灌浆充实有关,会对植物产生毒害作用[12],但本试验中农艺性状方面无明显肉眼可见毒害症状。而“川麦 28”等品种在 Se150 下粒重仍有所增加。合选 198、鄂恩 1、扬辐 188、郑 9023 和 X325在 Se100 下粒重增幅较大,从产量角度而言具有较好的硒强化潜力。
3.2 基施纳米硒对小麦籽粒硒含量的影响
所有小麦品种 (系) 施硒后籽粒硒含量均有显著或极显著增加,这与前人研究的硒肥可以提高植物体内硒含量[29]的结果一致,但不同小麦品种籽粒总硒含量不同,因此可以通过筛选硒高效积累品种并进行外源硒强化,进而提高膳食中硒含量水平。根据国家质量技术监督局发布的 GB13105-1991《食品中硒限量卫生标准》规定,食品中含硒量(以硒计)成品粮 ≤ 0.3 mg/kg 的要求,施用纳米硒后,小麦籽粒中的总硒含量超过了标准,而对作物进行硒强化一般都会导致硒含量的超标[28,30-31],因此经过纳米硒处理后高硒含量的全麦面粉可以作为食品添加剂来利用。

表1 Se150 处理籽粒中营养元素含量变化一致的品种及其变幅Table 1 Cultivars showing consistent variations in nutrient contents under Se150 treatment
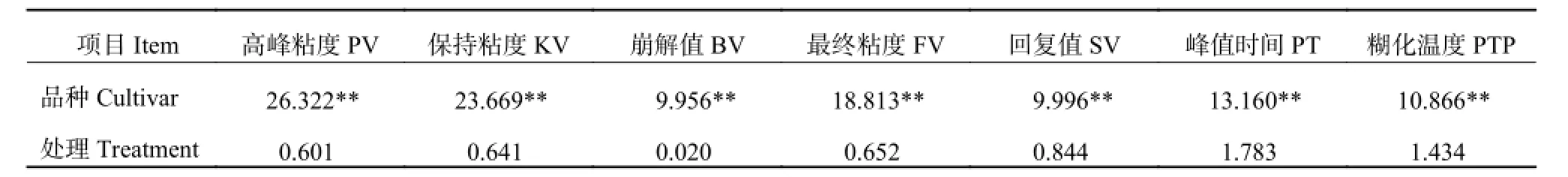
表2 施纳米硒小麦面粉各糊化特征值的 F 测验Table 2 F-test of wheat flour rapid viscosity analyser properties under different Se treatments
3.3 不同小麦品种在施硒条件下籽粒中硒形态的差异
不同小麦品种在硒处理后其籽粒中的硒形态和占比各异,这可能与不同品种在硒代谢的路径中的基因差异及其表达不同有关。在不同的硒形态中,通过植物富集转化的有机态硒更有利于人体吸收和利用。本研究结果表明硒代蛋氨酸 (SeMet) 是小麦籽粒中含量最多的有机含硒形态,和前人在其他作物上的研究结果基本一致[6],相较于无机硒具有更高的吸收和利用效率[32],作为抗氧化剂[33],在很多疾病的研究中都表现出较强的抗病能力,甚至在艾滋病[34]和神经元保护[35]的研究中也有重要作用;硒代半胱氨酸(SeCys) 是第 21 种人体标准氨基酸,也是合成谷胱甘肽氧化酶的中心氨基酸,谷胱甘肽氧化酶可以提高动植物的抗氧化性。硒半胱氨酸在改善认知功能下降、降低 tau 蛋白的过度磷酸化水平及缓解阿耳茨海默氏病等方面也有重要作用[36]。本研究中“X359”和“优选 14”中硒半胱氨酸含量较高,可以作为硒生物强化或育种利用的潜在品种 (系);甲基硒代半胱氨酸 (MeSeCys) 是一种天然的含硒氨基酸,是硒代半胱氨酸的甲基化衍生物,近年来它作为一种新型抗癌药物而被用于临床及科研实验研究中,在胃癌[37]、乳腺癌[38]、肝癌[39]等癌症的缓解和治疗方面的研究取得了很多进展。本研究表明,甲基硒代半胱氨酸在小麦品种中的差异是极显著的 (P < 0.01),因此筛选富含甲基硒代半胱氨酸的小麦品种对于国民预防癌症具有重要的意义。综合考虑籽粒总硒含量和硒形态特征,X359、郑 9023、合选 198 和宁麦资18 在硒强化或育种利用方面具有较大的应用潜力。
3.4 硒与硫元素之间的关系
硒处理会影响小麦籽粒中其他元素的积累,但不同元素对硒的响应不同。基施纳米硒肥会极显著提高籽粒的硫元素含量,相关性分析表明,Se100 和Se150 中硒和硫之间均存在极显著正相关 (r1= 0.516, r2= 0.499, P < 0.01),即硒和硫之间总体上 (大部分品种) 存在协同作用,但少部分品种施硒后硫含量有所降低 (表 1)。Zhu 等研究结果表明六价硒和硫之间存在拮抗作用[40],刘新伟等通过试验发现硫可以降低四价外源硒条件下小麦籽粒中硒含量[41]。因此,硒和硫的关系可能比较复杂,协同或拮抗关系很可能与基因型本身有关。
3.5 基施纳米硒对小麦面粉特性的影响
RVA 各参数中,高峰粘度、保持粘度、崩解值、回复值和最终粘度,是衡量淀粉糊化特性的最重要指标[42]。本研究结果表明,RVA 的几个特征值主要由基因型决定,基施纳米硒对 RVA 参数特征值影响不显著,说明基施纳米硒对小麦面粉糊化特性的影响可以不予考虑。
4 结论
1) 基施纳米硒肥可以显著提高小麦籽粒中的硒含量,增幅因品种而异,且低浓度硒肥可以提高小麦粒重,对小麦面粉的糊化特性没有显著影响。
2) 纳米硒强化后,小麦籽粒中的硒主要以有机态硒的形式存在,其中 SeMet 为主要存在形态,其次为 MeSeCys 和 SeCys,但含量和比例在品种间存在差异。
3) 综合考虑粒重、总硒含量和硒形态,“郑9023”和“合选 198”为理想的硒生物强化载体品种或可作为硒强化育种利用的亲本。
[1]Speckmann B, Grune T. Epigenetic effects of selenium and their implications for health[J]. Epigenetics, 2015, 10(3): 179-190.
[2]Zhang Z C, Zhang J S, Xiao J R. Selenoproteins and selenium status in bone physiology and pathology[J]. Biochimica et Biophysica Acta-General Subjects, 2014, 1840(11): 3246-3256.
[3]Fernandes A P, Gandin V. Selenium compounds as therapeutic agents in cancer[J]. Biochimica et Biophysica Acta-General Subjects, 2015, 1850(8): 1642-1660.
[4]Chen J S. An original discovery: selenium deficiency and Keshan disease (an endemic heart disease) [J]. Asia Pacific Journal of Clinical Nutrition, 2012, 21(3): 320-326.
[5]Kolesnikova L I, Karpova E A, Vlasov B Y, et al. Lipid peroxidationantioxidant defense system during toxic liver damage and its correction with a composite substance containing selenium and arabinogalactan [J]. Bulletin of Experimental Biology and Medicine, 2015, 159(3): 225-228.
[6]程义 勇. 《中国居民膳 食营养素参考摄入量 》2013修订版 简介[J].营养学报, 2014, (4): 313-317. Cheng Y Y. Brief introduction of the revised edition Reference intake of nutrients in dietary for Chinese people [J]. Acta Nutrimenta Sinica, 2014, (4): 313-317.
[7]Yu D, Liang D, Lei L, et al. Selenium geochemical distribution in the environment and predicted human daily dietary intake in northeastern Qinghai, China[J]. Environmental Science and Pollution Research, 2015, 22(15): 11224-11235.
[8]Ducsay L, Lozek O. Effect of selenium foliar application on its content in winter wheat grain[J]. Plant Soil and Environment, 2006, 52(2): 78-82.
[9]Chilimba A D C, Young S D, Black C R, et al. Agronomic biofortification of maize with selenium (Se) in Malawi[J]. Field Crops Research, 2012, 125: 118-128.
[10]李韬, 兰国防. 植物硒代谢机理及其以小麦为载体进行补硒的策略[J]. 麦类作物学报, 2012, 32(1): 173-177. Li T, Lan G F. Metabolism of selenium in plants and the strategies for selenium-biofortification using wheat[J]. Journal of Triticeae Crops, 2012, 32(1):173-177.
[11]Yuan L X, Zhu Y Y, Lin Z Q, et al. A novel selenocystineaccumulating plant in selenium-mine drainage area in Enshi, China[J]. Plos One, 2013, 8(6): e65615.
[12]White P J. Selenium accumulation by plants[J]. Annals of Botany, 2016, 117(2): 217-235.
[13]Djanaguiraman M, Devi D D, Shanker A K, et al. Seleninm - an antioxidative protectant in soybean during senescence[J]. Plant and Soil, 2005, 272(1-2): 77-86.
[14]Hartikainen H, Xue T L. The promotive effect of selenium on plant growth as triggered by ultraviolet irradiation[J]. Journal of Environmental Quality, 1999, 28(4): 1372-1375.
[15]Kong L A, Wang M, Bi D L. Selenium modulates the activities of antioxidant enzymes, osmotic homeostasis and promotes the growth of sorrel seedlings under salt stress[J]. Plant Growth Regulation, 2005, 45(2): 155-163.
[16]Hawrylak-Nowak B, Matraszek R, Szymanska M. Selenium modifies the effect of short-term chilling stress on cucumber plants[J]. Biological Trace Element Research, 2010, 138(1-3): 307-315.
[17]Chu J Z, Yao X Q, Zhang Z N. Responses of wheat seedlings to exogenous selenium supply under cold stress[J]. Biological Trace Element Research, 2010, 136(3): 355-363.
[18]Li H F, Mcgrath S P, Zhao F J. Selenium uptake, translocation andspeciation in wheat supplied with selenate or selenite[J]. New Phytologist, 2008, 178(1): 92-102.
[19]Lyons G, Ortiz-Monasterio I, Stangoulis J, et al. Selenium concentration in wheat grain: Is there sufficient genotypic variation to use in breeding?[J]. Plant and Soil, 2005, 269(1-2): 369-380.
[20]孙崇延, 李德安, 冯杰, 等. 施加硒化肥对麦粒的化学元素及氨基酸含量的影响[J]. 微量元素与健康研究, 1995, 12(3): 39-40, 45. Sun C Y, Li D A, Feng J, et al.The effect of selenium fertilizer on the concentration of the chemical elements and amino acids of wheat[J]. Trace Elements and Health Research, 1995, 12(3): 39-40, 45
[21]Boldrin P F, De Figueiredo M A, Yang Y, et al. Selenium promotes sulfur accumulation and plant growth in wheat (Triticum aestivum) [J]. Physiologia Plantarum, 2016, 158(1): 80-91.
[22]刘强, 田建珍, 李佳佳. 小麦粉粒度对其糊化特性影响的研究[J].现代面粉工业, 2012, (6): 16-20. Liu Q, Tian J Z, Li J J. Effects of wheat flour particle size on the gelatinization characteristics[J]. Modern Flour Milling Industry, 2012, (6): 16-20.
[23]蒋达, 张洪程, 刘艳阳, 等. 地力水平与施氮量对小麦淀粉RVA谱特征的影响[J]. 麦类作物学报, 2009, 29(1): 134-140. Jiang D, Zhang H C, Liu Y Y, et al. Effects of N application rate on RVA profile characters under different land fertility levels[J]. Journal of Triticeae Crops, 2009, 29(1): 134-140.
[24]Crosbie G M D, Dewan T. Wheat quality for Japanese flour milling and noodle industries[J]. Journal of Agriculture Western Australia, 1990, 31.
[25]Toykawah H, Rubenthaler G L, Powers J R, Schanus E G. Japanese noodle qualities.Ⅱ. Starch components[J]. Cereal Chemistry, 1989, 66(5): 387-391.
[26]阎俊, 张勇, 何中虎. 小麦品种糊化特性研究[J]. 中国农业科学, 2001, (1): 9-13. Yan J, Zhang Y, He Z H. Investigation on paste property of Chinese wheat[J]. Scientia Agricultura Sinica, 2001, (1): 9-13.
[27]Nawaz F, Ashraf M Y, Ahmad R, et al. Supplemental selenium improves wheat grain yield and quality through alterations in biochemical processes under normal and water deficit conditions[J]. Food Chemistry, 2015, 175: 350-357.
[28]Boldrin P F, Faquin V, Ramos S J, et al. Soil and foliar application of selenium in rice biofortification[J]. Journal of Food Composition and Analysis, 2013, 31(2): 238-244.
[29]Alfthan G, Eurola M, Ekholm P, et al. Effects of nationwide addition of selenium to fertilizers on foods, and animal and human health in Finland: From deficiency to optimal selenium status of the population[J]. Journal of Trace Elements in Medicine and Biology, 2015, 31: 142-147.
[30]Galinha C, Sanchez-Martinez M, Pacheco A M, et al. Characterization of selenium-enriched wheat by agronomic biofortification[J]. Journal of Food Science and Technology, 2015, 52(7): 4236-4245.
[31]Longchamp M, Castrec-Rouelle M, Biron P, et al. Variations in the accumulation, localization and rate of metabolization of selenium in mature Zea mays plants supplied with selenite or selenate[J]. Food Chemistry, 2015, 182: 128-135.
[32]Davis T Z, Tiwary A K, Stegelmeier B L, et al. Comparative oral dose toxicokinetics of sodium selenite and selenomethionine[J]. Journal of Applied Toxicology, 2017, 37: 231-238.
[33]Panchuk R R, Skorokhyd N R, Kozak Y S, et al. Antioxidants selenomethionine and D-pantethine decrease the negative side effects of doxorubicin in NL/Ly lymphoma-bearing mice[J]. Croatian Medical Journal, 2016, 57(2): 180-192.
[34]Watanabe L M, Barbosa Junior F, Jordao A A, et al. Influence of HIV infection and the use of antiretroviral therapy on selenium and selenomethionine concentrations and antioxidant protection[J]. Nutrition, 2016, 32(11-12): 1238-1242.
[35]窦贺荣, 李贤慧, 张新昌, 等. 补充硒蛋氨酸促进低氧/复氧损伤神经元存活作用的体外研究[J]. 营养学报, 2013, (4): 369-373. Dou H R, Li X H, Zhang X C, et al. Study on neuron survival during hypoxia/reoxygenation after selenomethionine supplement in vitro[J]. Acta Nutrimenta Sinica, 2013, (4): 369-373.
[36]Zhang Z H, Chen C, Wu Q Y, et al. Selenomethionine reduces the deposition of beta-amyloid plaques by modulating beta-secretase and enhancing selenoenzymatic activity in a mouse model of Alzheimer's disease[J]. Metallomics Integrated Biometal Science, 2016, 8:782-789.
[37]向琴, 邹金艳, 易三凤, 等. 甲基硒代半胱氨酸上调硒结合蛋白1的表达对胃癌SGC7901细胞增殖的影响[J]. 现代肿瘤医学, 2016, 24(15): 2343-2346. Xiang Q, Zou J Y, Yi S F, et al. Effect of methylselenocysteine in upregulation selenium -binding protein 1 for gastric carcinoma cells SGC7901 proliferation in vitro [J]. Modernoncology, 2016, 24(15):2343-2346.
[38]谢蒙蒙, 黄秋, 罗雅婕, 等. 硒-甲基硒代半胱氨酸对乳腺癌细胞端粒 酶 活 性 和hTERT基 因 表 达 的 影 响[J]. 营 养 学 报, 2014, (5):471-474. Xie M M, Huang Q, Luo Y J, et al. Effect of Se-methylselenocystein (MSC) on telomerast activity and hTERT gene experssion in human bresat cancer cells[J]. Acta Nutrimenta Sinica, 2014, (5): 471-474.
[39]王智. 硒-甲基硒代半胱氨酸诱导肝癌SMMC-7721细胞株的凋亡及机制探讨[J]. 江苏医药, 2012, (11): 1263-1266. Wang Z. A study on Se-methyselenocysteine inducing hepatocelluar carcinoma cell apoptosis and its mechanism[J]. Jiangsu Medical Journal, 2012, (11): 1263-1266.
[40]Zhu Y G, Pilon-Smits E A H, Zhao F J, et al. Selenium in higher plants: understanding mechanisms for biofortification and phytoremediation[J]. Trends in Plant Science, 2009, 14(8): 436-442.
[41]刘新伟, 段碧辉, 赵小虎, 等. 外源四价硒条件下硫对小麦硒吸收的影响机制[J]. 中国农业科学, 2015, (2): 241-250. Liu X W, Duan B H, Zhao X H, et al. Effects of sulfur on selenium uptake in wheat and its mechanism when amended with selenite[J]. Scientia Agricultura Sinica, 2015, (2): 241-250
[42]张勇, 何中虎. 我国春播小麦淀粉糊化特性研究[J]. 中国农业科学, 2002, (5): 471-475. Zhang Y,He Z H. Investigation on paste property of spring-sown Chinese wheat[J]. Scientia Agricultura Sinica, 2002, (5): 471-475.
Effects of nano-selenium fertilization on selenium concentration of wheat grains and quality-related traits
【Objectives】This article reported the influences of selenium application on the total and different forms of selenium concentration in wheat grains, which would clarify the potential of biofortification of Se in wheat through fertilization. 【Methods】Pot experiment was conducted and 110 wheat cultivars were collected as materials. The nano-Se was applied. Three levels of Se 0 (CK), 100 (Se100) and 150 mg/kg soil (Se 150) were designed and applied uniformly into soil before wheat planting. The 100-kernel weight was investigated. The concentrations of Se, Ca, Mg, Cu, Fe, Mn, Zn and S were measured using ICP. Using LC-UV-AFS method, the forms of Se were identified and quantified in the cultivars with total Se content in top ten, which were under treatment of Se150. Fifty-nine cultivars were randomly selected for paste viscosity evaluation using Rapid ViscoAnalyser. Data were analyzed using softwares Matlab2014 and SPSS16. 【Results】The mean 100-kernel weight for CK, Se100 and Se150 was 3.78, 4.11, and 3.70 g, respectively, and the total Se concentration wasaveraged 2.00, 12.46 and 17.35 mg/kg, respectively. Se was mainly existed in organic forms, of which selenomethionin (SeMet) was the predominant form, and then were methylated selenocysteine (MeSeCys) and selenocysteine (SeCys). Trace amounts of inorganic forms of Se4+and Se6+were also identified in some cultivars. Application of nano-Se fertilizer had either synergistically or antagonistically effects on the accumulation of the tested nutrients, but did not on the flour paste viscosity properties, which were mainly determined by genotypes of the cultivars.【Conclusions】Fertilization of nano-Se markedly increased the concentration of Se in wheat grains. Se in grains predominantly presented in organic forms of selenomethionin. Application of Se 100 mg/kg soil is effective in increment of 100-kernel weight, and application of Se 150 mg/kg soil will reduce it. Se fertilization had not significant impact on wheat flour paste viscosity properties.
wheat; nano-selenium fertilizer; selenium concentration; selenium form; flour paste viscosity;
LI Tao1, SUN Fa-yu1, GONG Pan1, WANG An1, YUAN Lin-xi2,3, YIN Xue-bin2,3
( 1 Jiangsu Provincial Key Laboratory of Crop Genetics and Physiology; Key Laboratory of Plant Functional Genomics of Ministry of Education, Wheat Research Center, Yangzhou University, Yangzhou, Jiangsu 225009, China; 2 Suzhou Selenium Valley Technology Co., Ltd, Suzhou, Jiangsu 215123, China; 3 Advanced Lab for Selenium and Human Health, Suzhou Institute of University of Science and Technology of China, Suzhou, Jiangsu 215123, China )
2016-05-30 接受日期:2016-09-07
江苏高校优势学科建设工程资助项目(PAPD);江苏省青年自然科学基金(BK2012202)资助。
李韬(1971—),男,甘肃会宁人,主要从事小麦遗传育种方面的研究。E-mail:taoli@yzu.edu.cn
