苹果树腐烂病菌生长发育相关突变体筛选及侧翼序列分析
2017-03-23程明明高小宁黄丽丽
冯 浩,程明明,张 冕,高小宁,黄丽丽
(西北农林科技大学 植物保护学院,陕西杨凌 712100)
苹果树腐烂病菌生长发育相关突变体筛选及侧翼序列分析
冯 浩,程明明,张 冕,高小宁,黄丽丽
(西北农林科技大学 植物保护学院,陕西杨凌 712100)
由弱寄生菌苹果黑腐皮壳属真菌(ValsamaliMiyabe et Yamada)引起的苹果树腐烂病是苹果枝干上的重大病害。探究病菌生长发育的分子机制对于新农药靶标位点的开发具有重要理论意义。在前期建立的苹果树腐烂病菌ATMT突变体库的基础上,利用PDA培养基常规培养法,对野生型菌株03-8及230株供试突变体菌株的菌落形态、大小以及繁殖体产生情况进行观察分析,筛选得到表型发生变化的突变体共67株(占突变体总数的29.13%)。其中11株突变体的菌落大小异常(4.78%);25株突变体的菌落颜色发生变化(10.87%);18株突变体的菌落边缘变为不规则(7.83%);9株突变体的繁殖体最初形成时间推迟至50 d 左右(3.91%);17株(7.39%)突变体的繁殖体产生数量发生显著变化。利用TAIL-PCR对10株表型变化的突变体进行侧翼序列扩增,获得3条T-DNA侧翼序列,序列分析发现1个为细胞色素氧化酶1(cytochrome oxidase 1),其他2个均为含有核糖体L34 (Ribosomal L34)的假想蛋白。筛选获得67个生长发育受到影响的突变体,并鉴定得到2个候选的调控病菌生长发育的相关基因,为揭示苹果树腐烂病菌生长发育的分子机理奠定基础。
苹果树腐烂病;Valsamali;突变体;生长发育;侧翼序列
由弱寄生菌苹果黑腐皮壳属真菌(ValsamaliMiyabe et Yamada)引起的苹果树腐烂病是苹果枝干上的重大病害[1]。主要引起苹果树枝干皮层腐烂,被称为苹果树的“癌症”,给苹果产业造成巨大的经济损失。病原菌生长发育与侵染致病环节息息相关,因此,从分子水平探究影响病原菌生长发育的关键因子能够为全面揭示病菌致病分子机制奠定基础,有利于新靶标位点的发现和新农药的开发。
在植物病原真菌功能基因组研究中,反向遗传学方法是研究功能基因最常用的方法。通过ATMT 技术构建T-DNA真菌插入突变体库,筛选致病表型突变体,克隆相关突变基因,进而通过正向和反向遗传学等手段进行致病相关基因功能的研究。目前,利用该方法已经对多种植物病原真菌的致病相关基因进行了研究并取得重要进展。稻瘟病菌(Magnaportheoryzae)、大丽轮枝菌(Verticilliumdahliae)、链格孢(Alternariaalternata)、灰葡萄孢(Botrytiscinerea)及菜豆间座壳(Diaporthephaseolorum)等病菌的ATMT突变体库相继被构建,并鉴定到一 批与生长速率、产孢能力及致病力相关的重要基因[2-10]。可见,通过筛选病菌ATMT突变体库中生长发育相关突变体,进而克隆突变基因,并通过正向和反向遗传学等手段进行功能研究,有利于对病菌生长发育及致病机制的深入理解。
此前研究中,西北农林科技大学植物保护学院果树病害病原生物学及综合防治研究室已经建立了苹果树腐烂病菌ATMT突变体库,为苹果树腐烂病菌功能基因研究提供丰富的材料[11]。黄定宣等[12]对突变体库中230株突变体表型进行筛选,鉴定到阿魏酸酯酶等致病相关基因,但并未开展生长发育相关基因的鉴定工作。并且该突变体库包括8 000多个突变体,急需深入挖掘突变体资源,以便全面解析病菌的生长发育机制。为此,本研究从突变体库中选取230株突变体菌株,通过表型分析筛选生长发育受到影响的突变体,并通过侧翼序列的扩增对相关基因进行分离和初步注释,以期为揭示苹果树腐烂病菌生长发育调控机制奠定基础,并为新农药靶标位点的开发及病害有效防治提供理论依据。
1 材料与方法
1.1 菌 株
苹果树腐烂病菌野生型菌株03-8以及230株苹果树腐烂病菌ATMT突变体均由西北农林科技大学植物保护学院果树病害综合治理研究室提供。
1.2 培养基
PDA培养基、LB培养基。
1.3 试剂和引物合成
氨苄青霉素(Amp)购自Sigma(美国),用量为 100 mg/L;PMD19 T-Simple连接酶(TaKaRa,中国大连)、Taq酶(TaKaRa,中国大连)等购自宝生物工程(大连)有限公司;胶回收试剂盒购自Biomiga(美国)。
1 mol/L Tris-HCl(pH 8.0)、0.5 mol/L EDTA(pH 8.0)、50×TAE缓冲液母液(pH 8.5)及EB染色液等。
引物合成均由上海生工生物工程技术服务有限公司完成(表1)。

表1 TAIL-PCR扩增引物Table 1 Primer list for TAIL-PCR
1.4 菌株的活化
苹果树腐烂病菌野生型菌株03-8及230株突变体均在常规PDA培养基上25 ℃黑暗培养3 d。
1.5 菌落大小的测定
在活化菌株的菌落边缘取直径7 mm的菌饼,接种于PDA培养基上25 ℃黑暗培养3 d。以十字交叉法测定菌落中心到边缘的大小,各菌株均重复3次,每个重复以野生型菌株作对照,试验重复3次。运用SPSS 16.0软件,对所测定的数据进行t检验。
1.6 菌落形态的记录
活化菌株于25 ℃黑暗培养5 d后,观察记录菌落的颜色/质地、菌落边缘、气生菌丝疏密程度。每个菌株设3个重复,试验重复3次。
1.7 繁殖体形成时间及数量统计
将黑暗培养5 d后的培养皿置于24 h光照条件下继续培养,同时监测繁殖体形成时间,并于培养60 d后统计突变体和野生型菌株繁殖体个数。每个菌株设3个重复培养皿(d=9 cm),试验重复3次。
1.8 侧翼序列的扩增及比对分析
利用CTAB法对10个突变体菌株进行DNA提取。采用热不对称交错PCR(TAIL-PCR)对表型变异的突变体菌株的右侧翼序列进行扩增,参照Liu等[13]方法,参照Tsuji 等[14]和Mullins等[15]引物。PCR反应体系均为:10×Buffer 2 μL、Mg2+1.2 μL、dNTPs 0.4 μL、特异引物(10 μmol/L) 0.2 μL、AD混合引物(10 μmo/L;)1 μL、模板1 μL、Taq酶 0.3 μL、用灭菌双蒸水补足20 μL。上一轮反应产物稀释1 000倍之后作为下一轮的模板。第1轮PCR反应条件为:95 ℃预变性5 min,94 ℃ 1 min,62 ℃ 1 min,72 ℃ 2 min为1个循环,进行5个循环;94 ℃ 1 min,25 ℃ 1 min,以0.2 ℃/s的速度升温至72 ℃,72 ℃ 2 min为1个循环,进行2个循环;94 ℃ 30 s,64 ℃ 30 s,72 ℃ 2 min,94 ℃ 30 s,64 ℃ 30 s,72 ℃ 2 min,94 ℃ 30 s,45 ℃ 30 s,72 ℃ 2 min为1个循环,进行12个循环;72 ℃延伸5 min,最后16 ℃保温。第2轮PCR反应条件为:95 ℃预变性5 min,94 ℃ 1 min,62 ℃ 1 min,72 ℃ 2 min为1个循环,进行5个循环;94 ℃ 30 s,64 ℃ 30 s,72 ℃ 2 min,94 ℃ 30 s,64 ℃ 30 s,72 ℃ 2 min,94 ℃ 30 s,45 ℃ 30 s,72 ℃ 2 min为1个循环,进行10个循环;94 ℃ 30 s,45 ℃ 1 min,72 ℃ 2 min为1个循环,进行5个循环;72 ℃延伸5 min,最后16 ℃保温。第3轮PCR反应条件为:94 ℃预变性5 min;94 ℃ 45 s,45 ℃ 45 s,72 ℃ 2 min为1个循环,进行35个循环;72 ℃延伸5 min,16 ℃保温。
以10个苹果树腐烂病菌突变体基因组 DNA 为模板,对其 T-DNA 插入位点基因右侧的序列进行扩增、回收、测序。测序结果经过DNAMAN(Version 7)进行序列比对,获侧翼序列,并对序列进行tblastx(http://blast.ncbi.nlm.nih.gov/Blast.cgi)注释分析。
2 结果与分析
2.1 突变体菌落大小的测定
通过十字交叉法测量野生型菌株03-8及230株突变体菌株3 d的菌落大小,筛选得到11株菌落大小与野生型菌株03-8有显著差异的菌株,其中3个菌株3 d的菌落大小相对于03-8显著减小(表2,图1)
2.2 突变体菌落形态的观察
通过对野生型菌株及230株供试突变体菌株进行菌落形态(包括菌落颜色、形状及气生菌丝的疏密程度)的观察,筛选得到菌落颜色有变异的菌株25株,其中18株变异为菌落正面灰白色,背面黑褐色,7株变异为菌落正背面均为黄褐色,并且18株菌落边缘变异为不规则类型(表3,图2)。

表2 3 d菌落大小显著变化的菌株Table 2 Strains with significant changes in colony size on the 3rd day
注:同列数据后不同小写字母表示在0.05水平差异显著(t检验)。
Note:Different lowercase letters following data in the column indicate significantly difference at 0.05.
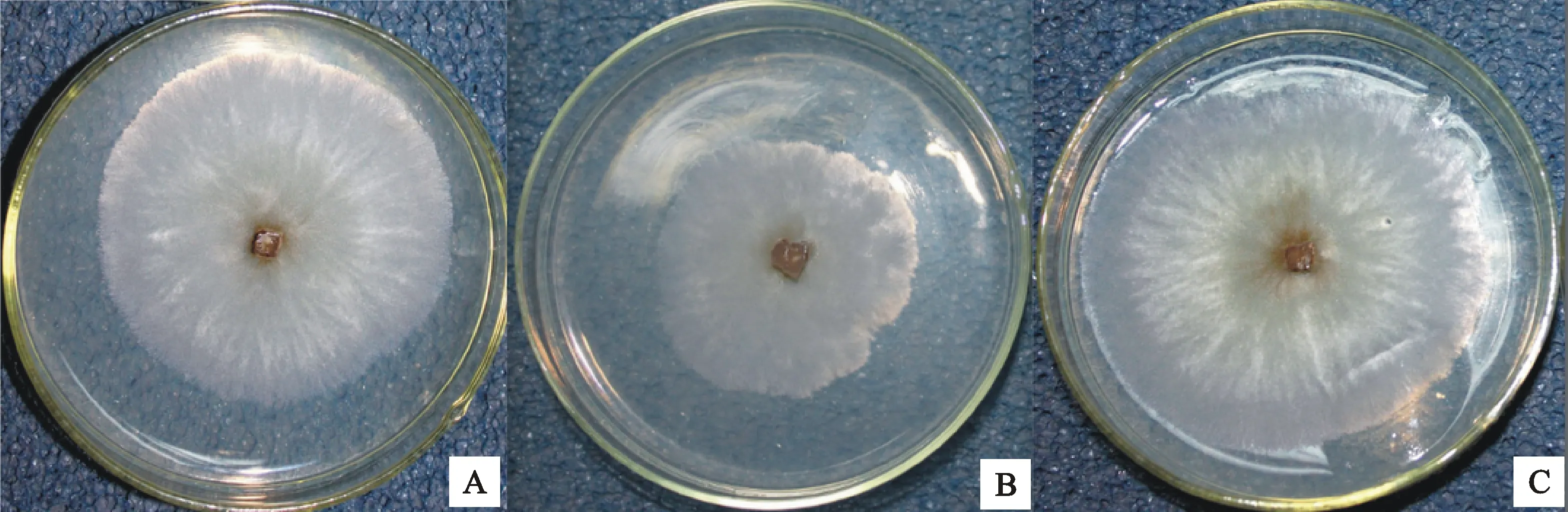
A.野生型菌株03-8 WT strain 03-8; B.3 d菌落大小显著减小的菌株 Strain which colony size reduced significantly on the 3rd day; C.3 d菌落大小显著增大的菌株 Strain with significant increase of colony size on the 3rd day

图1 野生型菌株与菌落大小显著变化的突变体菌株(PDA,25 ℃,3 d)Fig.1 WT and mutant strains with significant changes in colony size(PDA,25 ℃,3 d)
(续表2 Continued table 2)
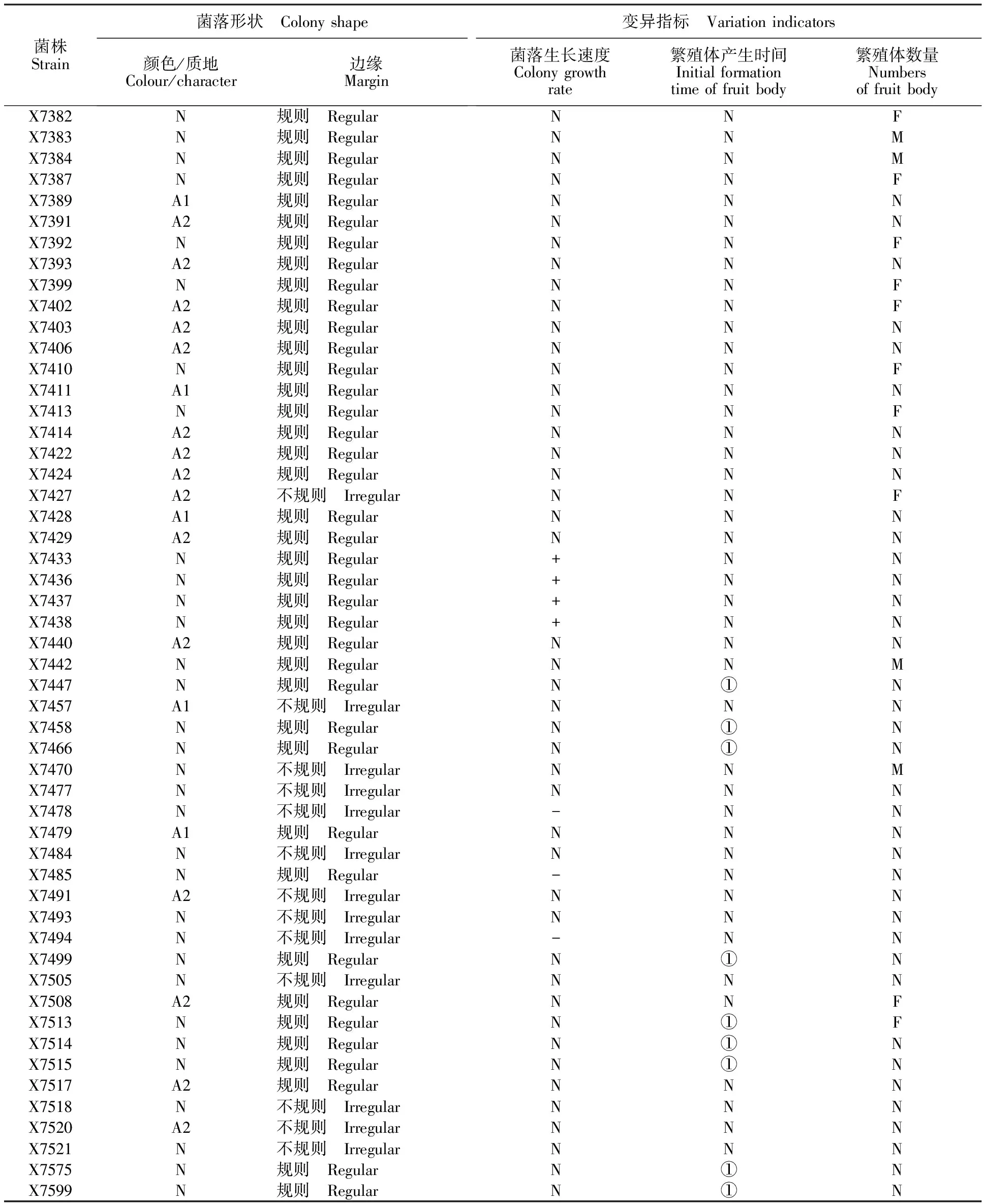
菌株Strain菌落形状 Colonyshape颜色/质地Colour/character边缘Margin变异指标 Variationindicators菌落生长速度Colonygrowthrate繁殖体产生时间Initialformationtimeoffruitbody繁殖体数量NumbersoffruitbodyX7382N规则 RegularNNFX7383N规则 RegularNNMX7384N规则 RegularNNMX7387N规则 RegularNNFX7389A1规则 RegularNNNX7391A2规则 RegularNNNX7392N规则 RegularNNFX7393A2规则 RegularNNNX7399N规则 RegularNNFX7402A2规则 RegularNNFX7403A2规则 RegularNNNX7406A2规则 RegularNNNX7410N规则 RegularNNFX7411A1规则 RegularNNNX7413N规则 RegularNNFX7414A2规则 RegularNNNX7422A2规则 RegularNNNX7424A2规则 RegularNNNX7427A2不规则 IrregularNNFX7428A1规则 RegularNNNX7429A2规则 RegularNNNX7433N规则 Regular+NNX7436N规则 Regular+NNX7437N规则 Regular+NNX7438N规则 Regular+NNX7440A2规则 RegularNNNX7442N规则 RegularNNMX7447N规则 RegularN①NX7457A1不规则 IrregularNNNX7458N规则 RegularN①NX7466N规则 RegularN①NX7470N不规则 IrregularNNMX7477N不规则 IrregularNNNX7478N不规则 Irregular-NNX7479A1规则 RegularNNNX7484N不规则 IrregularNNNX7485N规则 Regular-NNX7491A2不规则 IrregularNNNX7493N不规则 IrregularNNNX7494N不规则 Irregular-NNX7499N规则 RegularN①NX7505N不规则 IrregularNNNX7508A2规则 RegularNNFX7513N规则 RegularN①FX7514N规则 RegularN①NX7515N规则 RegularN①NX7517A2规则 RegularNNNX7518N不规则 IrregularNNNX7520A2不规则 IrregularNNNX7521N不规则 IrregularNNNX7575N规则 RegularN①NX7599N规则 RegularN①N
注:N 代表突变体所有生物学特性与野生型无显著差别;A1代表菌落颜色质地异常,正面灰白色背面黑褐色;A2代表菌落颜色质地异常,正背面均为褐色;+代表培养3 d的菌落大小比野生型菌株显著增大;-代表培养3 d的菌落大小比野生型菌株显著减小;①表示繁殖体初始形成时间推迟至50 d左右;M代表繁殖体增多,是野生型菌株03-8的2倍以上,超过每皿400个;F代表繁殖体减少,不足野生型菌株03-8的一半,即少于每皿100个。
Note:N means that no significant difference was found between mutants and wild type strains; A1 means there is obvious difference of colonial colour,grey to white on front and brown to dark on back; A2 means there is obvious difference of colonial colour,both front and back is brown; + means the colony of strains becomes much bigger than the wild type after 3 d cultivation; - means the colony of strains become smaller obviously than the wild type after 3 d cultivation;① means the time for generation of propagules is postponed almost 50 d. M means generations of propagules increase to more than double times of the wild type 03-8,with more than 400 per petri dish; F means generation of propagules reduce to less than half of the wild type 03-8,with less than 100 per petri dish.
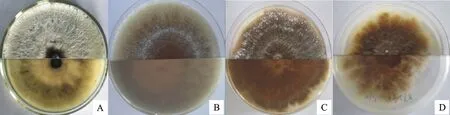
A.野生型菌株03-8,正面乳白色,背面黄褐色 WT 03-8,the front is milky white,and the back is yellow to brown;B.正面背面均为黄褐色 Both front and back are yellow to brown;C.正面灰白色,背面黑褐色 Front is gray to white and the back is brown to dark;D.边缘不规则 Margin is irregular
图2 4种菌落形态代表型菌株(PDA,25 ℃)
Fig.2 Four kinds of colony morphology on representative strain(PDA,25 ℃)
2.3 突变体繁殖体的产生情况
野生型菌株03-8在24 h光照条件下接种于PDA培养基上培养15 d左右即可见黑色的繁殖体。统计230株供试菌株繁殖体形成时间发现,大部分突变体在接种于PDA培养基上15~20 d产生繁殖体。 10株突变体的繁殖体最初形成时间延迟,其中9株推迟50 d左右出现繁殖体。通过野生型菌株03-8及230株突变体接种于PDA培养基上光照培养60 d时产生的繁殖体数量统计发现:野生型菌株03-8在接种60 d时繁殖体为每皿200个(d=9 cm),5株突变体的繁殖体产生量超过每皿400个,比野生型菌株03-8多1倍以上,12株突变体的繁殖体产生量少于野生型菌株03-8的一半,繁殖体少于每皿100个(表3,图3)。
2.4 侧翼序列扩增及序列分析
本试验使用AD简并引物和3个特异性引物HS-1、HS-2、HS-3对23株突变体插入位点的右侧序列分别进行扩增,经过20 g/L琼脂糖凝胶电泳、胶回收、连接转化、菌落PCR验证后送公司测序(图4~5)。与野生型菌株03-8基因组序列比对后,最终得到3个突变体插入位点的右侧序列(表4)。序列经过tblastx注释分析表明,突变体X7174的侧翼序列被注释为细胞色素氧化酶1(cytochrome oxidase 1),X7177、和X7156的侧翼序列注释为含有核糖体L34 (Ribosomal L34)的假想蛋白。
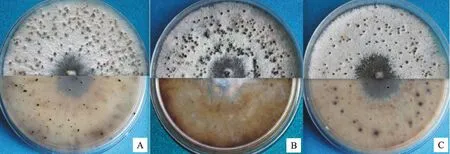
A.野生型菌株03-8 WT strain 03-8;B.繁殖体明显增多的突变体菌株 Mutants with significant increase of fruit bodies;C.繁殖体显著减少的突变体菌株 Mutants with significant decrease of fruit bodies
图3 野生型菌株以及突变体繁殖体产生情况(PDA,25 ℃,60 d)
Fig.3 Fruit body formation of wild strain and mutants(PDA,25 ℃,60 d)
3 讨论与结论
苹果树腐烂病是苹果生产上的第一大病害,严重威胁着苹果产业的健康发展。目前,已经对病菌的种类、生物学特性、致病力分化、侵染过程、致病机制及发生流行规律等方面开展了系统研究[16-23]。然而关于病菌的生长发育机制尚未全面了解,通过探究病菌生长发育机制对于农药新的靶标位点的发现及病害有效防控策略的制定具有重要意义。
随着农杆菌介导的遗传转化体系(Agrobacteriumtumefaciens-mediated transformation,ATMT)在植物病原真菌中的成功应用,建立病菌ATMT突变体库并结合TAIL-PCR技术分离鉴定相关基因成为研究病菌重要基因功能的突破口[24-26]。并利用该技术成功克隆到与病原真菌致病性与抗药性等重要生物学性状的相关基因并证实其生物学功能[27-28]。本试验共筛选实验室ATMT突变体库内菌株230株,大部分菌株在菌落形态、菌株生长速率以及繁殖体产生情况上与野生型菌株03-8并没有显著变化,只有少数突变体菌株表现明显的差异,这种现象出现的可能原因是:首先,外源T-DNA插入病原基因组位点是随机的,其插入位点可能是病原菌某些与表型相关基因的关键位点,也可能是基因组中不会引起表型明显变化的位点,由于是随机插入,插入到关键位点的几率将很小,所以大部分突变体菌株与野生型菌株表型相似;其次,表型的改变不仅会受到外源基因的影响,还同时会受到病原菌本身的株系、遗传转化体系以及插入拷贝数等诸多因素的影响。

1. X7174第2轮 Second application of X7174; 2~3. X7174第3轮 The third application of X7174; 4.X7177第2轮 The second application of X7177; 5~6.X7177第3轮 The third application of X7177;M.marker DL2000
图4 TAIL-PCR第2轮与第3轮对比电泳
Fig.4 Electrophoresis of second and third application of TAIL-PCR

1~2. X7174; 3~4. X7177;M.marker DL2000

菌株Strain突变体Mutant基因注释Geneannotation表型变化PhenotypicchangeX7174C01/VM00381细胞色素氧化酶1Cytochromeoxidase1生长速度增快IncreasedgrowthrateX7177NTP9J3XB014含有核糖体L34的假想蛋白Hypotheticalprotein(RibosomalL34)生长速度增快IncreasedgrowthrateX7156NTP9J3XB014含有核糖体L34的假想蛋白Hypotheticalprotein(RibosomalL34)生长速度增快Increasedgrowthrate
对本次筛选到的10株菌株进行侧翼序列扩增,得到3条侧翼序列条带,其中有1条侧翼序列基因注释为细胞色素氧化酶 1(cyt 1),另一条注释为含有核糖体L34 (Ribosomal L34)的假想蛋白。细胞色素氧化酶(cytochrome oxidase)是具有电子传递链末端的酶,是呼吸电子传递链的第4个中心酶复合物,又被称为复合物IV(Complex IV),氰化物、硫化氢、叠氮化合物和CO都能够与细胞色素氧化酶结合,并抑制其活性,从而造成细胞的“化学窒息”。后续工作中还应通过Southern 杂交等方法确定突变体T-DNA拷贝数,基于全基因组信息获得相应基因的全长序列,并利用基因敲除技术对目标基因进行功能验证,明确目的基因在苹果树腐烂病菌生长发育中的作用。
Reference:
[1] WANG X L,WEI J L,HUANG L L,etal.Re-evaluation of pathogens causing Valsa canker on apple in China[J].Mycologia,2011,103(2):317-324.
[2] 贺春萍,林春花,廖奇亨,等.稻瘟病菌 T-DNA插入突变体库构建及致病相关突变体筛选[J].热带作物学报,2007,28(1):80-84.
HE CH P,LIN CH H,LIAO Q H,etal.Construction of a T-DNA insertion mutant library ofMagnaporthegriseaand identification of pathogenicity-related mutants[J].ChineseJournalofTropicalCrops,2007,28(1):80-84(in Chinese with English abstract).
[3] 徐后娟,徐荣燕,李多川.链格孢ATMT农杆菌介导转化体系的构建及弱毒突变株验证[J].基因组学与应用生物学,2009,28(6):1056-1062.
XU H J,XU R Y,LI D CH.Agrobacterium tumefaciens-mediated transformation ofAlternariaalternataand identification of hypovirulent mutant[J].GenomicsandAppliedBiology,2009,28(6):1056-1062(in Chinese with English abstract).
[4] 吴小燕,王教瑜,张 震,等.稻瘟病菌致病性增强突变体B11的基因分析[J].中国水稻科学,2009,23(6):611-615.
WU X Y,WANG J Y,ZHANG ZH,etal.Analysis on gene locus ofMagnaportheoryzaeB11,a pathogenicity-enhanced mutant[J].ChineseJournalofRiceScience,2009,23(6):611-615(in Chinese with English abstract).
[5] 冯 娟,朱廷恒,崔志峰,等.农杆菌介导的灰葡萄孢T-DNA 插入突变体库构建及插入位点分析[J].微生物学报,2010,50(2):169-173.
FENG J,ZHU T H,CUI ZH F,etal.Construction of T-DNA insertion mutants ofBotrytiscinereaviaAgrobacteriumtumefaciensmediated transformation and sequence analysis of insertion site[J].ActaMicrobiologicaSinica,2010,50(2):169-173(in Chinese with English abstract).
[6] 徐荣旗,汪佳泥,陈捷胤,等.棉花黄萎病菌 T-DNA插入突变体表型特征和侧翼序列分析[J].中国农业科学,2010,43(3):489-496.
XU R Q,WANG J N,CHEN J Y,etal.Analysis of T-DNA insertional flanking sequence and mutant phenotypic characteristics inVerticilliumdahliae[J].ScientiaAgriculturaSinica,2010,43(3):489-496(in Chinese with English abstract).
[7] JEON J,PARK S Y,CHI M H,etal.Genome-wide functional analysis of pathogenicity genes in the rice blast fungus[J].NatureGenetics,2007,39(4):561-565.
[8] GAO F,ZHOU B J,LI G Y,etal.A glutamic acid-Rich protein identified inVerticilliumdahliaefrom an insertional mutagenesis affects microsclerotial formation and pathogenicity[J].PLoSONE,2010,5(12):1-10.
[9] MARUTHACHALAM K,KLOSTERMA S J,KANG S,etal.Identification of pathogenicity-related genes in the vascular wilt fungusVerticilliumdahliabyAgrobacteriumtumefaciens-mediated T-DNA insertional mutagenesis[J].MolecularBiotechnology,2011,49(3):209-221.
[10] SEBASTIANES F L S,LACAVA P T,FAVAROL C L,etal.Genetic transformation ofDiaporthephaseolorum,an endophytic fungus found in mangrove forests,mediated byAgrobacteriumtumefaciens[J].CurrentGenetics,2012,58(1):21-33.
[11] HU Y,DAI Q Q,LIU Y Y,etal.Agrobacteriumtumefaciens-mediated transformation of the causative agent of Valsa canker of apple treeValsamalivar.mali[J].CurrentMicrobiology,2014,68(6):769-776.
[12] 黄定宣,胡 杨,孙光超,等.苹果树腐烂病菌T-DNA插入突变体表型及致病突变体研究[J].西北农林科技大学学报(自然科学版),2014,42(7):113-121.
HUANG D X,HU Y,SUN G CH,etal.Phenotype and pathogenicity ofValsamaliT-DNA insertion mutants[J].JournalofNorthwestA&FUniversity(NatureScienceEdition),2014,42(7):113-121(in Chinese with English abstract).
[13] LIU Y G,WHITTIER R F.Thermal asymmetric interlaced PCR:automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking[J].Genomics,1995,25(3):674-681.
[14] TSUJI G,FUJII S,FUJIHARAN,etal.Agrobacteriumtumefaciens-mediated transformation for random insertional mutagenesis inColletotrichumlagenarium[J].JournalofGeneralPlantPathology,2003,69(4):230-239.
[15] MULLINS E D,CHEN X,ROMAIN E P,etal.Agrobacterium-mediated transformation ofFusariumoxysporum:An efficient tool forinsertional mutagenesis and gene transfer[J].Phytopathology,2001,91(2):173-180.
[16] 臧 睿,黄丽丽,康振生,等.陕西苹果树腐烂病菌(Cytosporaspp.)不同分离株的生物学特性和致病性研究[J].植物病理学报,2007,37(4):343-351.
ZANG R,HUANG L L,KANG ZH SH,etal.Biological characteristics and pathogenicity of different isolates ofCytosporaspp.isolated from apple trees in Shaanxi province[J].ActaPhytopathologicaSinica,2007,37(4):343-351(in Chinese with English abstract).
[17] 臧 睿,黄丽丽.苹果树腐烂病菌分生孢子萌发及其影响条件研究[J].西北农业学报,2007,16(1):64-67.
ZANG R,HUANG L L.Study on the pycindiospore germination of apple tree Valsa canker pathogen[J].ActaAgriculturaeBoreali-occidentalisSinica,2007,16(1):64-67(in Chinese with English abstract).
[18] 杜战涛,李正鹏,高小宁,等.陕西省苹果树腐烂病周年消长及分生孢子传播规律研究[J].果树学报,2013,30(5):681-687.
DU ZH T,LI ZH P,GAO X N,etal.Study on the conidia dispersal and the disease dynamics of apple treecanker caused byValsamalivar.maliin Shaanxi[J].JournalofFruitScience,2013,30(5):681-687(in Chinese with English abstract).
[19] KE X W,HUANG L L,GAO X N,etal.Histological and cytological investigations of the infection and colonization of apple bark byValsamalivar.mali[J].AustralasianPlantPathology,2013,42(1):85-93.
[20] KE X W,YIN ZH Y,SONG N,etal.Transcriptome profiling to identify genes involved in pathogenicity ofValsamalion apple tree[J].FungalGeneticsandBiology,2014,68:31-38.
[21] YIN ZH Y,LIU H Q,LI ZH P,etal.Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark[J].NewPhytologist,2015,208(4):1202-1216.
[22] LI ZH P,YIN ZH Y,FAN Y Y,etal.Candidate effector proteins of the necrotrophic apple canker pathogenValsamalican suppress BAX-induced PCD[J].FrontierinPlantScience,2015,6:579.
[23] 宋 娜,戴青青,宋 娜,等.苹果树腐烂病菌 GTP-环化水解酶Ⅱ基因敲除载体构建及其突变体的表型分析[J].中国农业科学,2014,47(15):2980-2989.
SONG N,DAI Q Q,SONG N,etal.Construction of knockout vector of GTPcyclohydrolaseⅡ gene and mutant’s biological characteristics ofValsamali[J].ScientiaAgriculturaSinica,2014,47(15):2980-2989(in Chinese with English abstract).
[24] RHO H S,KANG S,LEE Y H.Agrobacteriumtumefaciens-mediated transformation of the plant pathogenic fungusMagnaporthegrisea[J].MolecularCells,2001,12(3):407-411.
[25] FRANDSEN R J N.A guide to binary vectors and strategies for targeted genome modication in fungi usingAgrobacteriumtumefaciens-mediated transformation[J].JournalofMicrobiologyMethod,2011,87(3):247-262.
[26] MICHIELSE C B,HOOYKAAS P J,VAN DEN HONDELC A M J J,etal.Agrobacterium-mediated transformation as a tool for functional genomics in fungi[J].CurrentGenetics,2005,48(1):1-17.
[27] BLAISE F,REMY E,MEYER M,etal.A critical assessment ofAgrobacteriumtumefaciens-mediated transformation as a tool for pathogenicity gene discovery in the phytopathogenic fungusLeptosphaeriamaculans[J].FungalGeneticsandBiology,2007,44(2):123-138.
[28] ZHANG Y J,ZHAO J J,XIE M,etal.Agrobacteriumtumefaciens-mediated transformation in the entomopathogenic fungusLecanicilliumlecaniiand development of benzimidazole fungicide resistant strains[J].JournalofMicrobiologicalMethods,2014,105:168-173.
(责任编辑:潘学燕 Responsible editor:PAN Xueyan)
Screening of Growth-Related Mutants ofValsamaliand Analysis of Flanking Sequence
FENG Hao,CHENG Mingming,ZHANG Mian,GAO Xiaoning and HUANG Lili
(College of Plant Protection,Northwest A&F University,Yangling Shaanxi 712100,China)
Apple valsa canker,caused by weakly parasitic fungus ofValsamali,is one of destructive diseases in apple. Research on molecular mechanisms of growth and development ofV.maliis of great importance in excogitating fungicide with new target-site. In this study,phenotypes including colonial morphology,size of colony and generation of propagules of 230 mutants with wild strain 03-8 were analyzed by using conventional PDA medium culture method. 67 mutants which accounted for 29.13% of obvious phenotypes changes were screened out. Among which,11(4.78%)strains were abnormal in size of colony; 25(10.87%)strains had obvious changes in colour of colony; 18(7.83%)strains had irregular colony margin; Propagules generation of 9(3.91%)strains was postponed about 50 days,futherly,the propagules quantity of 17 strains(7.39%)changed significantly. The flanking sequences of 10 mutant strains were amplified using TALI-PCR. Three flanking sequences of T-DNA were acquired,one of three is the cytochrome oxidase 1 and the other two ones were ribosome L34 (Ribosomal L34) hypothetical proteins. In brief,67 mutants were screened out,and two candidate genes related with the growth and development ofV.maliwere identified. The results will lay a foundation for revealing the molecular mechanism of pathogen growth and development.
Apple valsa canker;Valsamali; Mutant; Growth and development; Flanking sequence
FENG Hao,male,lecture. Research area: pathogenesis mechanism of fungi.E-mail:xiaosong04005@163.com
HUANG Lili,female,professor. Research area: pathogen biology and integrated control of fruit trees diseases.E-mail: huanglili@nwsuaf.edu.cn
2016-03-29
2016-04-30
陕西省自然科学基础研究计划青年人才项目(2016JQ3015)。
冯 浩,男,讲师,从事病菌致病机理研究。E-mail:xiaosong04005@163.com
黄丽丽,女,教授,主要从事果树病害病原生物学及病害综合防控研究。E-mail: huanglili@nwsuaf.edu.cn
日期:2016-12-20
S432.4+4
A
1004-1389(2017)01-0144-08
网络出版地址:http://www.cnki.net/kcms/detail/61.1220.S.20161220.1645.037.html
Received 2016-03-29 Returned 2016-04-30
Foundation item Basic Research Plan in Natural Science of Shaanxi Province of China(No. 2016JQ3015 ).
