高脂日粮添加胆汁酸对草鱼组织脂肪酸组成的影响
2017-03-23陈昊杰田晶晶周继术
陈昊杰,田晶晶,周继术,吉 红
(西北农林科技大学 动物科技学院,陕西杨凌 712100)
高脂日粮添加胆汁酸对草鱼组织脂肪酸组成的影响
陈昊杰,田晶晶,周继术,吉 红
(西北农林科技大学 动物科技学院,陕西杨凌 712100)
为研究高脂日粮添加胆汁酸对草鱼组织脂肪酸组成的影响,以豆油为主要油脂源设计3组等氮实用日粮,分别饲喂96尾草鱼(初体质量69.86 g±6.24 g)8周,设置对照组(CT,蛋白350 g/kg,脂肪50 g/kg)、高脂组(HL,蛋白350 g/kg,脂肪70 g/kg)与胆汁酸组(BA,蛋白350 g/kg,脂肪70 g/kg,胆汁酸60 mg/kg)。结果表明,组织脂肪酸组成反映饲料脂肪酸组成。BA组腹腔脂肪组织中总脂肪酸、n-6多不饱和脂肪酸(PUFA)和亚油酸(LA,18∶2n-6)占油脂的质量百分比显著低于HL组,其在肝胰脏中趋势相反。与HL组相比,BA组肝胰脏中n-3 PUFA占总脂肪酸的质量百分比显著升高,肌肉中则降低。3组肝胰脏中饱和脂肪酸(SFA)占油脂的质量百分比无显著差异;BA组腹腔脂肪组织中SFA占油脂的质量百分比显著低于HL组。BA组组织与饲料脂肪酸组成的相关性显著高于HL组,HL组显著高于CT组。综上所述,胆汁酸可能调控组织间脂肪酸的转运与蓄积,并提高组织与饲料脂肪酸组成的相关性。
高脂;胆汁酸;草鱼;脂肪酸组成
胆汁酸是在肝脏中由胆固醇合成的一类两性甾醇类化合物[1],是胆汁的重要组成部分。胆汁酸具有较强的表面活性,可有效乳化脂肪,改善不溶于水的物质(如胆固醇)在水中的溶解度,还能提高脂肪酶的活力,促进脂肪的消化吸收[2]。许多研究还发现胆汁酸可作为信号分子,通过与胆汁酸受体(如法尼醇X受体FXR)结合来调控代谢[3-4]。同时,胆汁酸可以提高动物免疫力,减少动物对细菌内毒素的吸收[5],具有较强的抗菌作用,可调节机体的肠道微生物区系[6],抑制肠道细菌的过度增殖,维护消化道健康。
脂肪不仅是细胞膜的组成部分,还能促进亲脂性营养物质的吸收,为机体提供能量及必需脂肪酸,是一种重要的营养素[7]。日粮中适宜的脂肪水平能促进机体生长,节约蛋白[8-9],但随着集约化养殖业的发展,为适应养殖品种快速生长的生产性需求,生产中往往使用高能量配合饲料,反而造成养殖鱼类体内脂肪的过度蓄积[8,10],严重影响鱼体的品质及商业价值[11-12]。另外,鱼体的脂肪酸组成反映饲料的脂肪酸组成[13-14],不同油脂源或脂肪水平会导致鱼体组织脂肪酸组成出现差异[14]。同时,不同组织具有不同的生理功能,如腹腔脂肪组织是脂肪储存的主要部位,肝脏是鱼类脂质代谢的主要组织,肌肉主要用于提供可食部分满足人类的消费需求,因此,不同组织与饲料脂肪酸组成的相关性也有一定差异[15-16]。
草鱼(Ctenopharyngodonidella)是一种原产于中国的草食性淡水鱼类,目前养殖面积广泛,中国2014年的产量达到538万t,占中国淡水养殖鱼类总产量的18.31%[17]。研究表明,相比于多数肉食性鱼类,草鱼对能量的需求及利用能力较弱[18],日粮适宜脂肪水平为40~50 g/kg[8,19],饲喂高脂日粮(脂肪水平60 g/kg以上)往往造成鱼体脂肪过度蓄积[20],特别是在肝胰脏[21]。胆汁酸能促进脂肪的消化吸收利用,在脂肪组织的分解利用过程中产生的游离脂肪酸也可被肌肉和肝胰脏摄取和利用[22]。已有研究发现胆汁酸对罗氏沼虾(Macrobrachiumrosenbergii)[22]和大菱鲆(Scophthalmusmaximus)[23]的肌肉脂肪酸组成有显著影响,但对淡水鱼类(如草鱼)组织脂肪酸组成的影响还未见报道。
因此,本研究在高脂日粮中添加胆汁酸饲喂草鱼,通过对草鱼组织中脂肪酸占总脂肪酸和组织油脂的质量百分比,以及组织-饲料脂肪酸组成的相关性进行分析,以研究高脂日粮添加胆汁酸对草鱼组织脂肪酸组成的影响,从而为胆汁酸影响水产动物脂肪代谢的研究提供参考资料。
1 材料与方法
1.1 试验饲料及原料
胆汁酸由山东龙昌动物保健品有限公司提供,质量分数大于950 g/kg,其中w(猪脱氧胆酸)∶w(鹅脱氧胆酸)∶w(猪胆酸)=7∶2 ∶1。其余饲料原料均购自华秦农牧有限公司(陕西杨凌)。鱼粉、肉骨粉、豆粕、菜粕、棉粕等作为主要蛋白源,豆油为主要脂肪源,添加10 g/kg的三氧化二铬(Cr2O3)作外源指示剂。配制3组等氮实用日粮,分别为对照组(CT,蛋白350 g/kg,脂肪50 g/kg)、高脂组(HL,蛋白350 g/kg,脂肪70 g/kg)和胆汁酸组(BA,蛋白350 g/kg,脂肪70 g/kg,胆汁酸60 mg/kg),试验饲料配方及常规成分见表1,脂肪酸组成分析见表2。所有原料彻底粉碎,与豆油、一定量的水混合后倒入制粒机,制成直径2.5 mm的颗粒,在通风阴凉处干燥24 h后置于-20 ℃冰箱保存。
1.2 饲养管理
草鱼幼鱼由陕西省安康市汉滨区流水镇某商业渔场提供。草鱼在水泥池(长4.75 m,宽1.65 m,高0.80 m)中,用商品饲料(蛋白350 g/kg,脂肪70 g/kg,通威股份有限公司提供,四川成都)暂养1个月。饥饿24 h后,挑选规格整齐、体质健壮的96尾幼鱼(初体质量69.86 g±6.24 g)随机分至6个缸(215 L,直径0.75 m,高度0.70 m),6个缸再随机分为3组,每组饲喂1种饲料。每缸配备加热棒维持水温28 ℃左右,光周期12 h/12 h。每天投喂3次至饱食,分别在8:30、12:30、16:30进行,为期8周。养殖期间水质测定结果为:溶氧(14.83±0.50) mg/L;pH 7.27±0.24,氨氮(0.10±0.04) mg/L。
1.3 样品采集
采样过程严格按照西北农林科技大学动物管理委员会的要求执行,尊重动物福利与道德规范。采样前,草鱼饥饿24 h后使用MS-222 (50 mg/L)麻醉。每缸解剖6尾鱼,取肝胰脏、腹腔脂肪组织、肌肉于-20 ℃保存,用于组织脂肪酸组成分析。
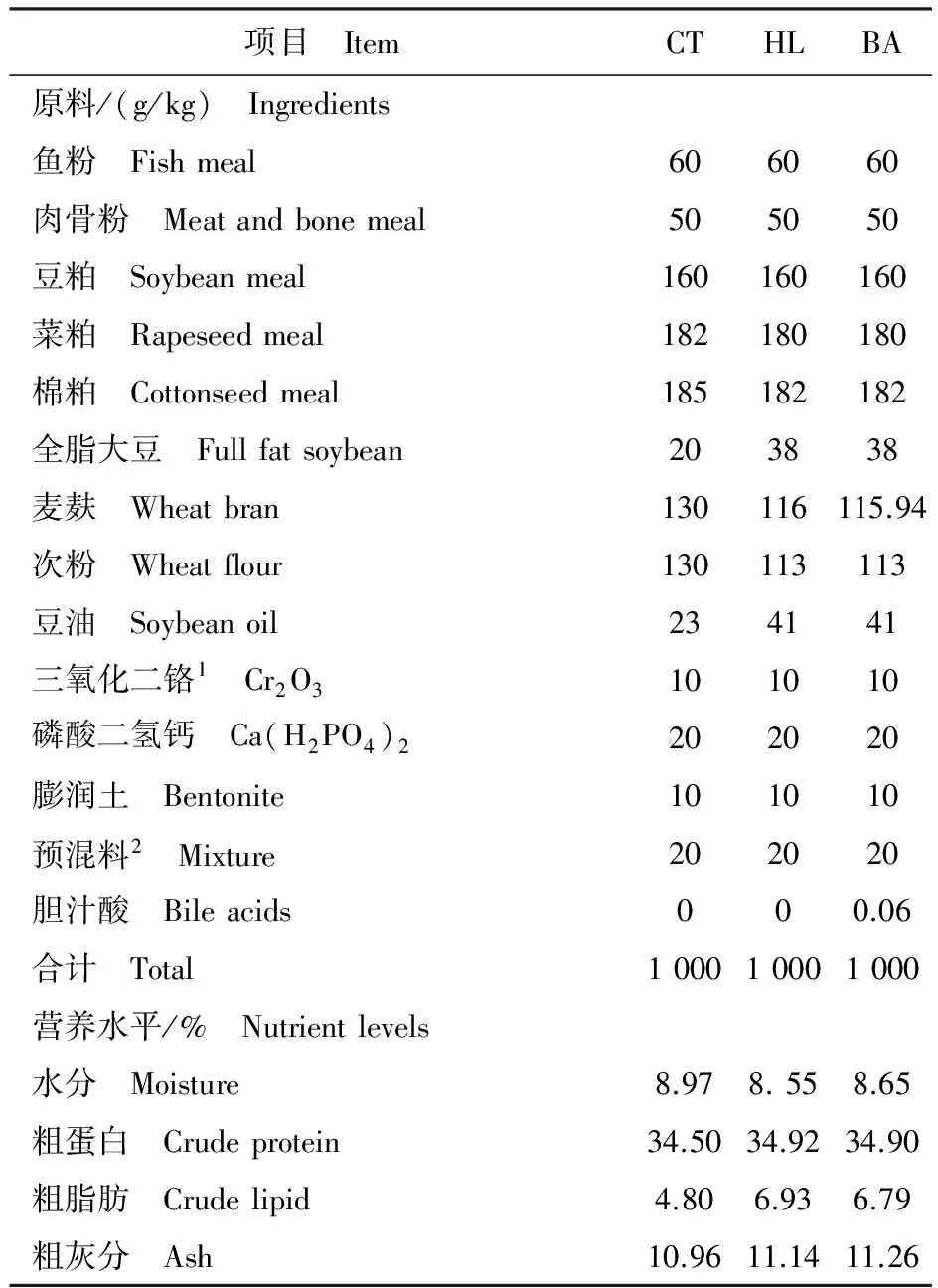
表1 饲料配方及营养水平Table 1 Formulation and nutrient levels of the experimental diets
注:1. 指示剂;2. 预混料含有10 g/kg的维生素和10 g/kg的矿物质。1 kg预混料包括维生素A 67 IU,维生素D 16.2 IU,维生素E 7.4 g,维生素K 340 mg,维生素B1670 mg,维生素B21 000 mg,维生素B6800 mg,维生素B121.4 mg,维生素C 10 g,泛酸2.65 g,叶酸 330 mg,烟酰胺5.35 g,氯化胆碱35 g,生物素34 mg,肌醇8 g,铁14 g,铜350 mg,锌4 g,锰1.4 mg,镁10 g,钴30 mg,碘40 mg,硒35 mg。
Note: 1.Inert marker; 2.Mixture contained 10 g/kg vitamin and 10 g/kg mineral; Ingredients including/1 kg vitamin A 67 IU,vitamin D 16.2 IU,vitamin E 7.4 g,vitamin K 340 mg,vitamin B1670 mg,vitamin B21 000 mg,vitamin B6800 mg,vitamin B121.4 mg,vitamin C 10 g,D-pantothenic acid 2.65 g,folic acid 330 mg,nicotinamide 5.35 g,choline chloride 35 g,biotin 34 mg,inositol 8 g,Fe 14 g,Cu 350 mg,Zn 4 g,Mn 1.4 mg,Mg 10 g,Co 30 mg,I 40 mg,Se 35 mg.
1.4 脂肪酸组成及相关性分析
根据Folch等[24]研究方法,称取饲料或组织5~10 g于50 mL离心管,加入溶液[V(氯仿)∶V(甲醇)=2∶1] 20 mL,高速匀浆,静置2 h,用定量滤纸过滤;然后加入4 mL蒸馏水,3 000 r/min 离心5 min,弃上清,下层40 ℃水浴蒸干;接着吸取约50 μL油脂并称量,记录粗提油质量。
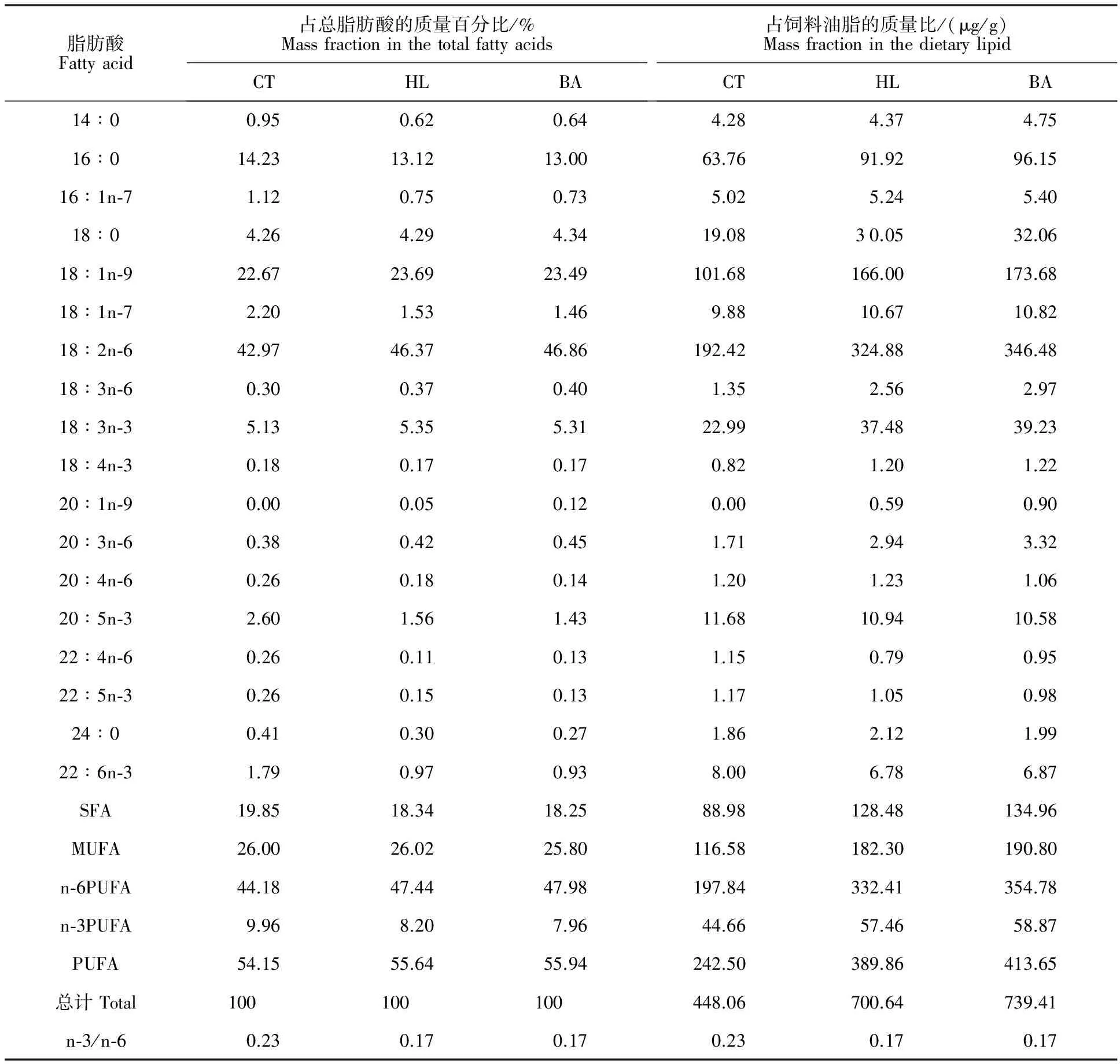
表2 饲料的脂肪酸组成Table 2 Fatty acid profiles of the experimental diets
注:SFA.饱和脂肪酸;MUFA.单不饱和脂肪酸;PUFA.多不饱和脂肪酸。下同。
Note:SFA,saturated fatty acid; MUFA,monounsaturated fatty acid;PUFA,polyunsaturated fatty acid.The same as below.
之后加入1 mL正己烷(色谱纯),再加入1 mL KOH-甲醇溶液(0.4 mol/L),进行30 min的甲酯化;最后加入2 mL去离子水,待分层后提取上层溶液,-20 ℃保存。
脂肪酸组成使用气相色谱仪(安捷伦7820a,安捷伦科技,美国)进行测定。脂肪酸甲酯通过与脂肪酸甲酯混合标准品(47015-U,Sigma-Aldrich,美国)进行比较鉴定。测定前取20 μL样品与5 μL十七烷酸甲酯(10 mg/mL,H4515,Sigma-Aldrich,美国)混匀,然后取1 μL混合物上机检测。每个样品测定3次。各类脂肪酸占总脂肪酸的质量百分比按面积归一化法计算,以百分比形式呈现。根据十七烷酸甲酯占总脂肪酸的质量百分比,计算饲料、草鱼肝胰脏与腹腔脂肪组织油脂中各类脂肪酸所占的质量百分比,公式如下:
MF(μg/g)=[M17(μg)×PF/P17]/Moil(g)式中,MF表示饲料或草鱼组织每克油脂中所含某一脂肪酸的质量;M17表示上机检测样品中十七烷酸甲酯的质量;PF表示饲料或组织中某一脂肪酸占总脂肪酸的质量百分比;P17表示上机检测样品中十七烷酸甲酯占总脂肪酸的质量百分比;Moil表示粗提油的质量。
此外,根据Tian等[15]研究方法,各类脂肪酸占总脂肪酸的质量百分比通过CORREL函数计算出皮尔森相关系数,以研究组织与饲料脂肪酸组成的相关性。同时,定义R并作图,以期获得不同脂肪酸在不同组织中的变化情况,计算公式如下:
R=log2(PT/PD)
式中,R表示组织脂肪酸和饲料脂肪酸的相关性程度;PT表示组织中某一脂肪酸占总脂肪酸的质量百分比;PD表示饲料中相应脂肪酸占总脂肪酸的质量百分比。
1.5 数据处理
试验数据采用“平均值±标准差”表示,使用SPSS18.0软件(Chicago,IL,USA)对脂肪酸组成数据进行单因素方差分析,对皮尔森相关系数进行组织和饲料的双因素方差分析,并用Duncan’s多重比较组间差异。
2 结果与分析
2.1 草鱼肝胰脏的脂肪酸组成分析
如表3所示,SFA方面,HL和BA组的SFA、16∶0和18∶0占总脂肪酸的质量百分比显著低于CT组 (P<0.05),但3组的SFA、14∶0、16∶0、18∶0及24∶0占油脂的质量比均无显著性差异(P>0.05)。MUFA方面,HL和BA组的16∶1n-7占总脂肪酸和油脂的质量比均低于CT组,20∶1n-9变化趋势相反。BA组MUFA和油酸(OA,18∶1n-9)占总脂肪酸的质量百分比显著低于CT组(P<0.05),而其占油脂的质量比在3组间均无显著性差异(P>0.05)。n-6 PUFA方面,HL与BA组的n-6 PUFA、亚油酸(LA,18∶2n-6)、18∶3n-6和20∶3n-6占总脂肪酸和油脂的质量比均显著高于CT组(P<0.05)。与HL组相比,BA组的n-6 PUFA、18∶3n-6、20∶3n-6及花生四烯酸(ARA,20∶4n-6)占油脂的质量比较高(P<0.05)。n-3 PUFA方面,3组间亚麻酸(LNA,18∶3n-3)占总脂肪酸和油脂的质量比均无显著性差异(P>0.05)。BA组的n-3 PUFA、22∶5n-3及二十二碳六烯酸(DHA,22∶6n-3)占油脂的质量比均显著高于HL组(P<0.05)。此外,HL与BA组的PUFA占总脂肪酸的质量百分比显著高于CT组,n-3/n-6呈现出相反的趋势(P<0.05)。BA组PUFA占油脂的质量比显著高于HL组(P<0.05)。
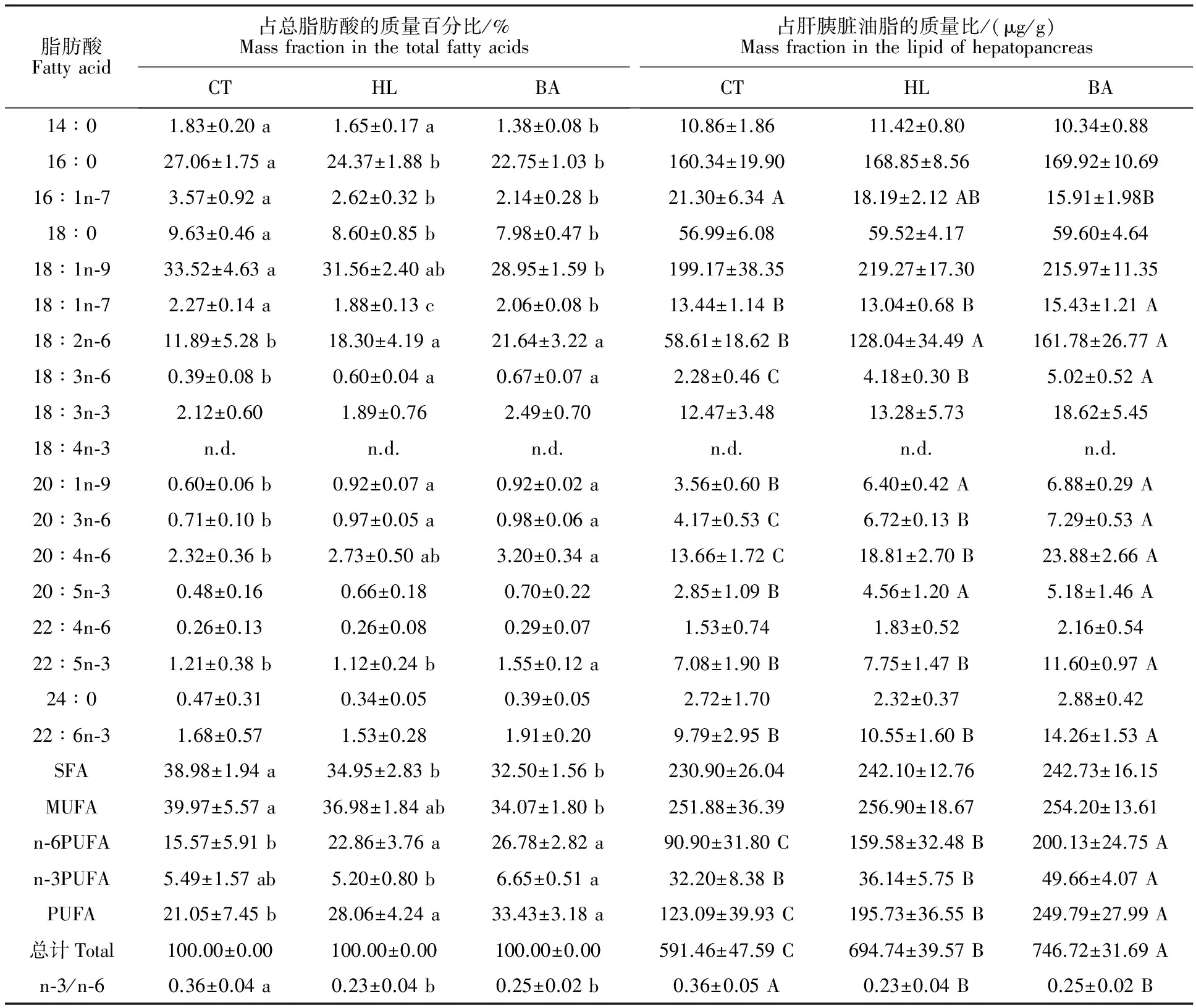
表3 草鱼肝胰脏的脂肪酸组成Table 3 Fatty acid profilesof the hepatopancreas in the grass carp
注:n.d.,未检测出脂肪酸。同行上标不同表示差异显著(P<0.05) (n=12)。脂肪酸占总脂肪酸的质量百分比差异使用小写字母,占肝胰脏油脂的质量比差异使用大写字母。下同。
Note:n.d.,Not detected. Different superscripts in the same row denote significant differences (P<0.05) (n=12). Differences of mass fraction used lowercase in the total fatty acids,and used uppercase in the lipid of hepatopancreas.The same as below.
2.2 草鱼腹腔脂肪组织的脂肪酸组成分析
如表4所示,SFA方面,HL和BA组的SFA、14∶0、16∶0、18∶0与24∶0占总脂肪酸和油脂的质量比均显著低于CT组 (P<0.05),其中BA组的SFA、14∶0和16∶0占油脂的质量比显著低于HL组(P<0.05)。MUFA方面,HL与BA组的MUFA和OA占总脂肪酸的质量百分比显著高于CT组,占油脂的质量比却低于CT组(P<0.05)。n-6 PUFA方面,HL与BA组n-6 PUFA、LA、18∶3n-6、20∶3n-6和ARA占总脂肪酸的质量百分比均显著高于CT组(P<0.05);而BA组n-6 PUFA和LA占油脂的质量比却显著低于HL组(P<0.05),ARA则高于HL组(P<0.05)。n-3 PUFA方面,HL和BA组的二十碳五烯酸(EPA,20∶5n-3)与DHA占总脂肪酸和油脂的质量比均显著低于CT组(P<0.05)。此外,BA组总脂肪酸、n-3 PUFA和LNA占油脂的质量比显著低于HL组(P<0.05)。n-3/n-6的变化情况与肝胰脏一致,表现为HL和BA组显著低于CT组(P<0.05)。
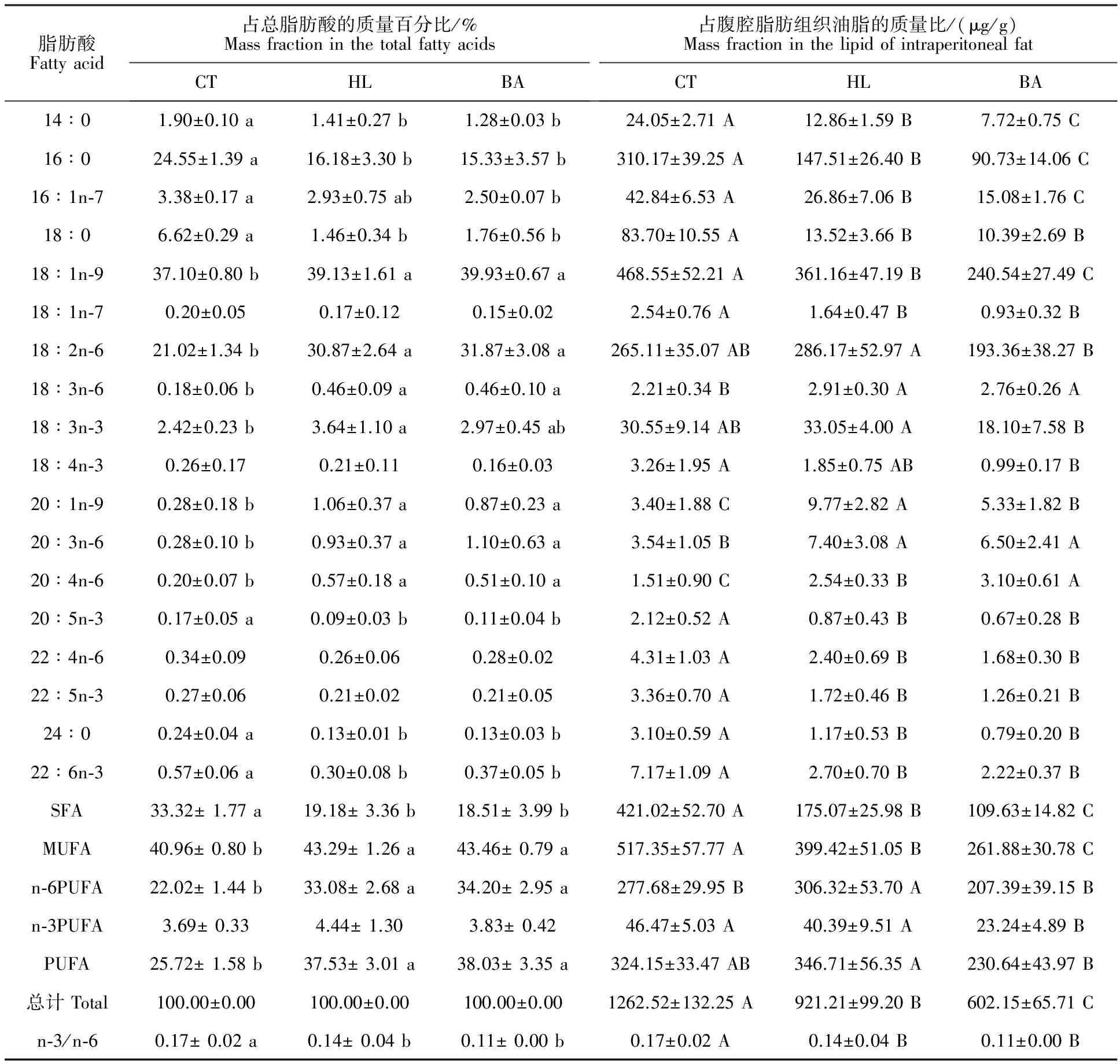
表4 草鱼腹腔脂肪组织的脂肪酸组成Table 4 Fatty acid profiles of the intraperitoneal fat in the grass carp
注:脂肪酸占总脂肪酸的质量百分比差异使用小写字母,占腹腔脂肪组织油脂的质量比差异使用大写字母。
Note: Differences of mass fraction used lowercase in the total fatty acids,and used uppercase in the lipid of intraperitoneal fat.
2.3 草鱼肌肉的脂肪酸组成分析
如表5所示,SFA方面,HL和BA组的SFA占总脂肪酸的质量百分比显著低于CT组(P<0.05),其中BA组SFA、16∶0、18∶0和24∶0占总脂肪酸的质量比显著低于HL组(P<0.05)。MUFA方面,HL组MUFA和OA占总脂肪酸的质量百分比显著低于CT与BA组(P<0.05)。n-6 PUFA方面,BA组n-6 PUFA和LA占总脂肪酸的质量百分比显著高于CT与HL组,而18∶3n-6、20∶3n-6、ARA及22∶4n-6变化趋势相反(P<0.05)。n-3 PUFA方面,BA组22∶5n-3、DHA及n-3 PUFA占总脂肪酸的质量百分比显著低于CT和HL组(P<0.05)。此外,HL与BA组的PUFA占总脂肪酸的质量比显著高于CT组,BA组的n-3/n-6显著低于HL组,HL组显著低于CT组(P<0.05)。
2.4 组织与饲料脂肪酸组成的相关性分析
如表6所示,BA组组织与饲料脂肪酸组成的相关性显著高于HL组,而HL组相关性显著高于CT组(P<0.05)。其中BA组肌肉与饲料的脂肪酸组成相关性显著高于HL组,而HL组显著高于CT组(P<0.05);HL与BA组肝胰脏、腹腔脂肪组织与饲料的脂肪酸组成相关性显著高于CT组(P<0.05)。排除饲料的影响,肌肉-饲料脂肪酸组成的相关性最高,肝胰脏与饲料的最低(P<0.05)。CT与BA组中,组织与饲料脂肪酸组成的相关性顺序为肌肉>腹腔脂肪组织>肝胰脏(P<0.05);HL组中,肌肉和腹腔脂肪组织与饲料脂肪酸组成的相关性显著高于肝胰脏(P<0.05)。
2.5R值分析饲料与鱼体脂肪酸组成的关系
图1~3分别反映草鱼肝胰脏、腹腔脂肪组织及肌肉与饲料脂肪酸组成的关系。如果饲料中的脂肪酸完全沉积在组织中,没有经过任何脂肪酸代谢(从头合成、分解等),折线会与X轴重合。若R>0,则该种脂肪酸在组织中优先沉积;若R<0,则该种脂肪酸在组织中被特异性代谢。结果显示,各组在同一组织中均呈现出相似的波动。与CT组相比,HL与BA组肝胰脏中LA、18∶3n-6、ARA、EPA、22∶4n-6及DHA的R值较高;腹腔脂肪组织中LA、18∶3n-6、20∶3n-6及ARA的R值较高。与CT和HL组相比,BA组肌肉中18∶3n-6、20∶3n-6及EPA的R值较低。3个组织中,16∶1n-7、20∶3n-6、ARA及22∶4n-6的R值较高,LA、LNA和EPA的R值较低。此外,肌肉中DHA的R值也较高。

表5 草鱼肌肉的脂肪酸组成Table 5 Fatty acid profiles of the muscle in the grass carp
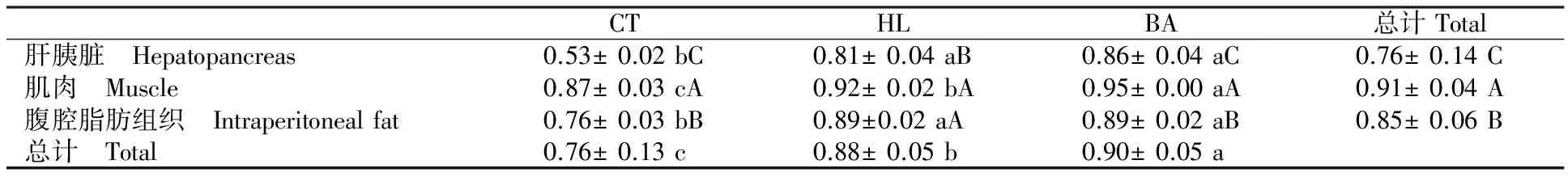
表6 组织与饲料脂肪酸组成的皮尔森相关系数Table 6 The Pearson correlation coefficient of the fatty acid profiles between the tissues and diets
注:每行差异使用小写字母,每列差异使用大写字母。
Note:The horizontal used lowercase and the vertical used uppercase.
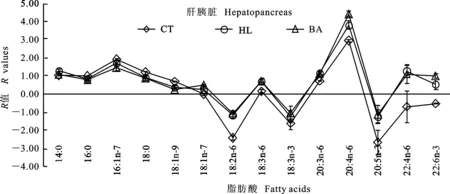
图1 草鱼肝胰脏脂肪酸组成的R值分析Fig.1 R values analysis of the fatty acid profiles in the hepatopancreas of grass carp
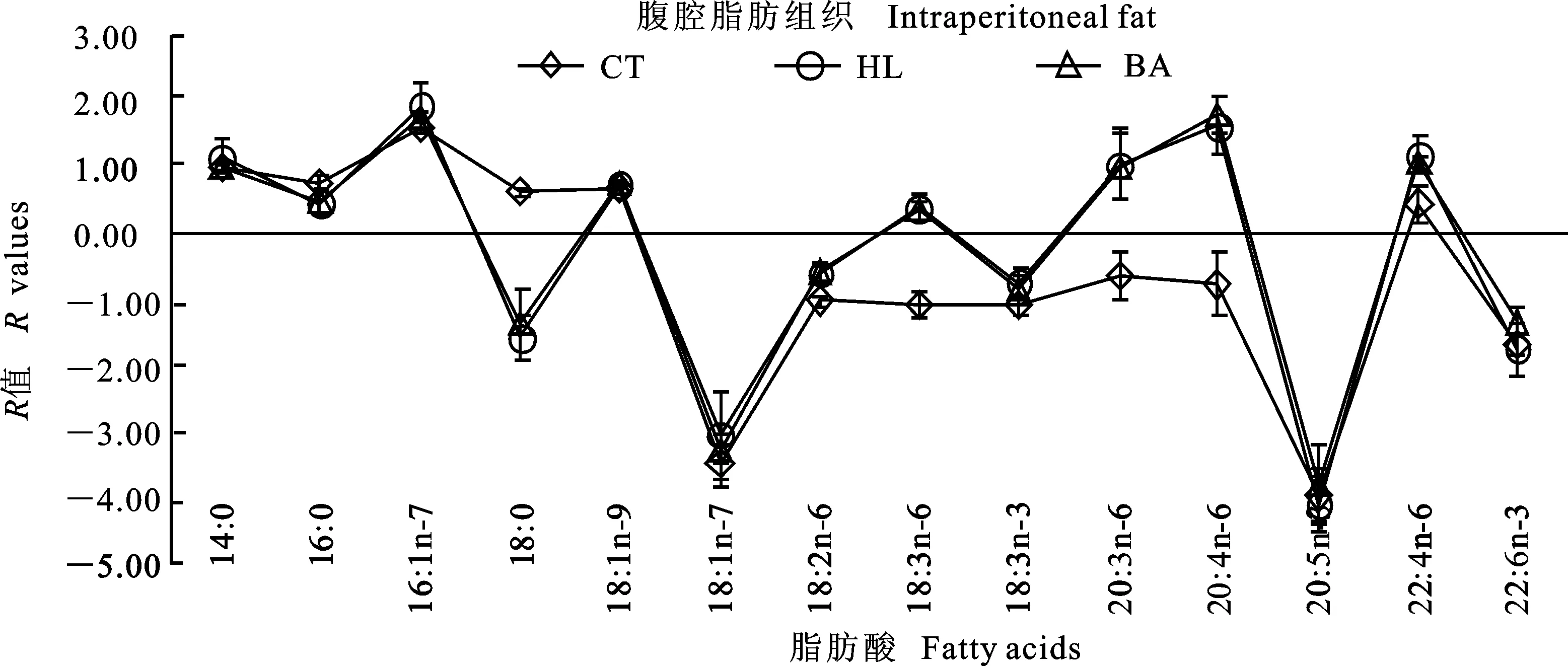
图2 草鱼腹腔脂肪组织脂肪酸组成的R值分析Fig.2 R values analysis of the fatty acid profiles in the intraperitoneal fat of grass carp
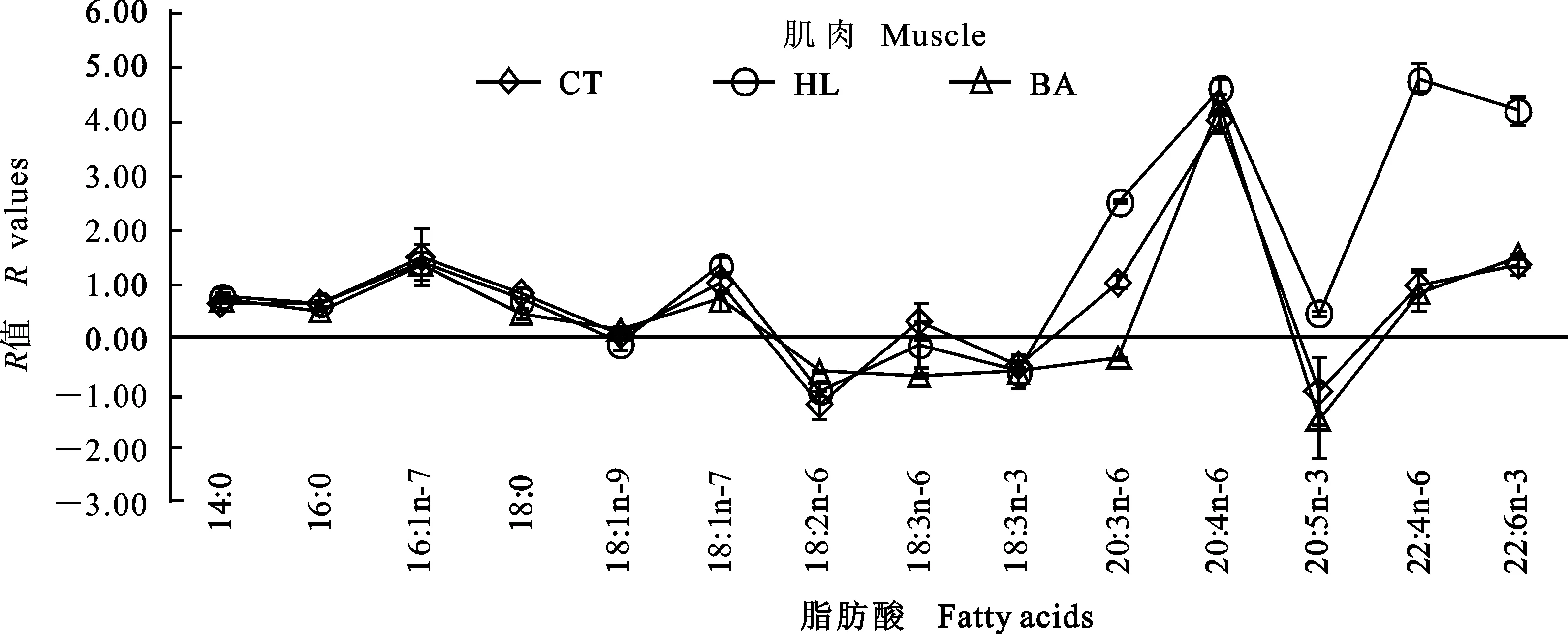
图3 草鱼肌肉脂肪酸组成的R值分析Fig.3 R values analysis of the fatty acid profiles in the muscle of grass carp
3 讨 论
脂肪是动物体必需脂肪酸和能量的主要来源,在鱼类营养中发挥重要作用[7]。三大营养物质中脂肪含有的能量最高。蛋白质虽然是水产动物饲料中最重要的营养物质,但因蛋白源成本较高,生产中往往通过提高饲料中的脂肪含量来节约蛋白[8-9]。然而,饲料中过高的脂肪水平会造成鱼体脂肪过度蓄积[8,10],且不同组织中脂肪蓄积程度不同,根据生理功能的不同,组织与饲料脂肪酸组成的相关性也不尽相同[15-16],因此不同的脂肪含量可能造成组织中脂肪酸占组织油脂的质量比的差异。胆汁酸可以促进脂肪的消化吸收利用,产生的游离脂肪酸能被其他组织利用[22],也能影响不同组织中脂肪酸占组织油脂的质量比。
3.1 草鱼组织的脂肪酸组成
鱼体组织的脂肪酸组成反映饲料的脂肪酸组成[13-14]。本研究中,组织脂肪酸组成与饲料脂肪酸组成表现出较高的相关性。本试验使用豆油作为主要油脂源,其富含LA[25],因此CT组饲料中LA占总脂肪酸(饲料油脂)的质量比为42.97% (192.42 μg/g),而HL和BA组分别为46.37% (324.88 μg/g)和46.86% (346.48 μg/g)。本研究中,HL与BA组组织(肝胰脏、腹腔脂肪组织和肌肉)中n-6 PUFA和LA占总脂肪酸的质量百分比均显著高于CT组(P<0.05),与饲料的结果相似。然而,BA组腹腔脂肪组织的总脂肪酸、n-6 PUFA和LA占油脂的质量比低于HL组,肝胰脏中却呈现相反的趋势。脂肪酸是过氧化物酶体增殖物激活受体α (PPARα)的天然配体[26]。脂肪酸信号通路(如LA)可以活化PPARα[27],而PPARα能抑制胆固醇7α-羟化酶(CYP7A1)的表达[28],从而抑制内源性胆汁酸的合成。BA组外源性添加的胆汁酸超过机体需求,在抑制胆汁酸合成的过程中,PPARα的活化可能需要更多的配体,因而机体从脂肪组织中分解更多的游离脂肪酸并转运至肝胰脏。淡水鱼类可通过延长与去饱和作用将C18PUFA转化为高不饱和脂肪酸(HUFA),如LA可转化为ARA,LNA可转化为EPA和DHA[7]。本研究发现,BA组肝胰脏及腹腔脂肪组织的18∶3n-6、20∶3n-6和ARA占总脂肪酸的质量百分比显著高于CT组(P<0.05),这可能是LA通过延长与去饱和作用转化生成。另外,BA组肝胰脏中18∶3n-6、20∶3n-6和ARA占油脂的质量比显著高于HL组(P<0.05),表明由脂肪组织转运至肝胰脏的LA进行转化。
本研究饲料中鱼粉含量相同,而饲料中的n-3 HUFA(如EPA,DHA)主要由鱼粉提供[29]。CT组饲料中EPA和DHA占总脂肪酸与饲料油脂的质量比均高于HL和BA组。HL与BA组腹腔脂肪组织的EPA和DHA占总脂肪酸与油脂的质量比均显著低于CT组(P<0.05),与饲料中的结果相似。与HL组相比,BA组肝胰脏的EPA、DHA和n-3 PUFA占总脂肪酸的质量百分比呈升高趋势,肌肉中则降低。王恒[22]报道饲喂胆汁酸后,罗氏沼虾肌肉中EPA、DHA及n-3 PUFA占总脂肪酸的质量百分比呈下降趋势;孙建珍等[23]研究发现,胆汁酸显著降低大菱鲆肌肉中EPA、DHA和n-3 PUFA占总脂肪酸的质量百分比,与本研究结果一致,表明胆汁酸可能促进n-3 HUFA在草鱼组织间的调配与利用。
SFA方面,HL和BA组组织(肝胰脏、腹腔脂肪组织和肌肉)中SFA占总脂肪酸的质量百分比显著低于CT组(P<0.05),与饲料的结果吻合。然而,HL和BA组饲料中SFA(如14∶0、16∶0)占饲料油脂的质量百分比高于CT组,3组草鱼肝胰脏中SFA占油脂的质量比却没有显著差异(P>0.05),这说明SFA在肝胰脏中的沉积有限,多余的SFA可能在肝胰脏中进行转化或者β-氧化[25]。另外,BA组腹腔脂肪组织中SFA占油脂的质量比显著低于HL组(P<0.05),表明饲料中添加胆汁酸可降低SFA在腹腔脂肪组织中的沉积。此外,BA组肌肉中SFA(如16∶0、18∶0和24∶0)占总脂肪酸的质量百分比显著低于HL组(P<0.05),与王恒[22]的结果矛盾。
3.2 草鱼组织与饲料脂肪酸组成的相关性
本研究对不同组织和饲料脂肪酸组成的相关性也进行分析。BA组组织与饲料脂肪酸组成的相关性最高(0.90),其次为HL组(0.88),最低为CT组(0.76)(P<0.05)。低脂日粮中的脂肪被吸收后用于代谢的比例较高,以维持机体的正常需要,造成相关性较低;高脂日粮中的脂肪被吸收后用于沉积的比例较高,因此相关性高。胆汁酸能提高脂肪酶的活力,促进脂肪的消化吸收[2]。本研究还发现,BA组草鱼的肠道脂肪酶活力高于HL组,而HL组高于CT组(未发表资料),与组织与饲料脂肪酸组成相关性的变化趋势一致,说明胆汁酸可能通过促进脂肪的消化吸收,提高其利用率,从而使更多脂肪酸沉积在组织中,最终提高组织与饲料脂肪酸组成的相关性。在组织方面,肌肉与饲料脂肪酸组成的相关性最高(0.91),其次为腹腔脂肪组织(0.85),最低为肝胰脏(0.76) (P<0.05)。Tian等[15]在松浦镜鲤(CyprinusspecularisSongpu)上的研究发现肌肉、腹腔脂肪组织与饲料脂肪酸组成的相关性显著高于肝胰脏,与本研究结果相似。肝胰脏作为重要的代谢组织,具有更强的脂肪酸代谢(去饱和/延长,β-氧化)能力,能通过自身调节以维持肝胰脏的脂肪酸组成[25],因而导致其相关性较低。腹腔脂肪组织是脂肪沉积的首要部位,是能量蓄积的主要场所,饲料中的脂肪被鱼体摄入后,经消化吸收转运,除用于分解供能,大部分转化为甘油三酯或者膜磷脂储存在脂肪组织中,这可能是导致其与饲料脂肪酸组成相关性高的原因[16]。肌肉与饲料脂肪酸组成的相关性高,且HL与BA组显著高于CT组,这可能是由于饲料中的脂肪在体内的异位过度蓄积进而导致其相关性增加。
R值反映组织脂肪酸变化的程度。同一组织中,3组均呈现出相似的波动,说明脂肪酸在同一组织中的代谢方式相同。草鱼的必需脂肪酸为LA和LNA[7]。本研究中LA的R值较低,20∶3n-6、ARA和22∶4n-6的R值较高,这很大程度上是因为LA通过去饱和或延长作用进行转化。肝胰脏和肌肉中LNA、EPA的R值较低,DHAR值较高,这可能是因为LNA转化为终产物DHA。脂肪酸能够影响肌肉的风味和营养特性,作为可食部分,肌肉中DHA的沉积有利于满足人类营养的需要。然而,腹腔脂肪组织中EPA和DHA的R值较低,原因可能为LNA 在脂肪组织内转化为HUFA的能力最低,也可能与饲料中这类脂肪酸的转运具有选择性有关[16]。另外有趣的是,3个组织中16∶1n-7的R值均较高,而不是OA,表明草鱼的16∶0比18∶0更优先进行Δ-9去饱和作用,这与松浦镜鲤上的研究结果一致[15]。
4 结 论
综上所述,组织的脂肪酸组成反映饲料的脂肪酸组成,但外源性添加胆汁酸可能调控组织间脂肪酸的转运与蓄积。胆汁酸还可能促进n-3 HUFA在草鱼组织间的调配与利用,降低腹腔脂肪组织中SFA的沉积,维持肝胰脏中SFA的稳定。另一方面,胆汁酸可提高组织与饲料脂肪酸组成的相关性,这可能是通过促进脂肪的消化吸收实现的。组织与饲料脂肪酸组成的相关性紧密,但组织间脂肪酸的沉积存在差异。其中肌肉的相关性最高,肝胰脏的最低,这可能与组织的生理功能差异有关。
Reference:
[1] HOFMANN A F,HAGEY L R.Bile acids:chemistry,pathochemistry,biology,pathobiology,and therapeutics[J].CellularandMolecularLifeSciences,2008,65(16):2461-2483.
[2] ROMANSKI K W.The role and mechanism of action of bile acids within the digestive system-bile acids in the liver and bile[J].AdvancesinClinicalandExperimentalMedicine,2007,16(6):793-799.
[3] LI Y,JADHAV K,ZHANG Y.Bile acid receptors in non-alcoholic fatty liver disease[J].BiochemicalPharmacology,2013,86(11):1517-1524.
[4] LANGHI C,PEDRAZ-CUESTA E,HARO D,etal.Regulation of human class I alcohol dehydrogenases by bile acids[J].JournalofLipidResearch,2013,54(9):2475-2484.
[5] POUPON R E,LINDOR K D,PARES A,etal.Combined analysis of the effect of treatment with ursodeoxycholic acid on histologic progression in primary biliary cirrhosis[J].JournalofHepatology,2003,39(1):12-16.
[6] BAJOR A,GILLBERG P G,ABRAHAMSSON H.Bile acids:short and long term effects in the intestine[J].ScandinavianJournalofGastroenterology,2010,45(6):645-664.
[7] TOCHER D R.Fatty acid requirements in ontogeny of marine and freshwater fish[J].AquacultureResearch,2010,41(5):717-732.
[8] DU Z Y,LIU Y J,TIAN L X,etal.Effect of dietary lipid level on growth,feed utilization and body composition by juvenile grass carp (Ctenopharyngodonidella)[J].AquacultureNutrition,2005,11(2):139-146.
[9] WANG J T,LIU Y J,TIAN L X,etal.Effect of dietary lipid level on growth performance,lipid deposition,hepatic lipogenesis in juvenile cobia (Rachycentroncanadum)[J].Aquaculture,2005,249(1):439-447.
[10] BORGES P,OLIVEIRA B,CASAL S,etal.Dietary lipid level affects growth performance and nutrient utilisation of Senegalese sole (Soleasenegalensis) juveniles[J].BritishJournalofNutrition,2009,102(7):1007-1014.
[11] DIAS J,ALVAREZ M J,DIEZ A,etal.Regulation of hepatic lipogenesis by dietary protein/energy in juvenile European seabass (Dicentrarchuslabrax)[J].Aquaculture,1998,161(1):169-186.
[12] RUEDA-JASSO R,CONCEICAO L E,DIAS J,etal.Effect of dietary non-protein energy levels on condition and oxidative status of Senegalese sole (Soleasenegalensis) juveniles[J].Aquaculture,2004,231(1):417-433.
[13] TOCHER D R,BELL J G,DICK J R,etal.Effects of dietary vegetable oil on atlantic salmon hepatocyte fatty acid desaturation and liver fatty acid compositions[J].Lipids,2003,38(7):723-732.
[14] 韩光明,王爱民,徐 跑,等.饲料脂肪水平对吉富罗非鱼体脂沉积及脂肪酸组成的影响[J].中国水产科学,2011,18(2):338-349.
HAN G M,WANG A M,XU P,etal.Effects of dietary lipid levels on fat deposition and fatty acid profiles of GIFT,Oreochromisniloticus[J].JournalofFisherySciencesofChina,2011,18(2):338-349 (in Chinese with English abstract).
[15] TIAN J J,LEI C X,JI H.Influence of dietary linoleic acid (18∶2n-6) and α-linolenic acid (18∶3n-3) ratio on fatty acid composition of different tissues in freshwater fish Songpu mirror carp,Cyprinuscarpio[J].AquacultureResearch,2015,doi:10.1111/are.12832.
[16] 田晶晶,萧培珍,吉 红,等.实用饲料对养殖草鱼体组织脂肪酸组成的影响[J].饲料工业,2013,34(20):22-27.
TIAN J J,XIAO P ZH,JI H,etal.Influence of practical diets on tissue fatty acid composition in reared grass carp (Ctenopharyngodonidellus) [J].FeedIndustryMagazine,2013,34(20):22-27 (in Chinese with English abstract).
[17] 农业部渔业渔政管理局.2014中国渔业统计年鉴[M].北京:中国农业出版社,2015.
FISHERY BUREAU OF DEPARTMENT OF AGRICULTURE.Fishery Yearbook in China in 2014[M].Beijing:Chinese Agriculture Press,2015 (in Chinese).
[18] DU Z Y,CLOUET P,HUANG L M,etal.Utilization of different dietary lipid sources at high level in herbivorous grass carp (Ctenopharyngodonidella):mechanism related to hepatic fatty acid oxidation[J].AquacultureNutrition,2008,14(1):77-92.
[19] LI C,LIU P,JIH,etal.Dietary n-3 highly unsaturated fatty acids affect the biological and serum biochemical parameters,tissue fatty acid profile,antioxidation status and expression of lipid-metabolism-related genes in grass carp,Ctenopharyngodonidellus[J].AquacultureNutrition,2015,21(3),373-383.
[20] DU Z Y,CLOUET P,ZHENG W H,etal.Biochemical hepatic alterations and body lipid composition in the herbivorous grass carp (Ctenopharyngodonidella) fed high-fat diets [J].BritishJournalofNutrition,2006,95(5):905-915.
[21] GUO X Z,LIANG X F,FANG L,etal.Effects of dietary non-protein energy source levels on growth performance,body composition and lipid metabolism in herbivorous grass carp (Ctenopharyngodonidella)[J].AquacultureResearch,2015,46(5):1197-1208.
[22] 王 恒.不同脂肪源及胆汁酸对罗氏沼虾生长性能及脂肪酸组成的影响[D].江苏扬州:扬州大学,2010.
WANG H.Effect of different lipid sources and bile acid on growth performance and fatty acid composition inMacrobrachiumrosenbergii[D].Yangzhou Jiangsu:Yangzhou University,2010 (in Chinese with English abstract).
[23] 孙建珍,王际英,马晶晶,等.不同脂肪水平下添加胆汁酸对大菱鲆(Scophthalmusmaximus)幼鱼生长、体组成和脂肪代谢的影响[J].海洋与湖沼,2014,45(3):617-625.
SUN J ZH,WANG J Y,MA J J,etal.Effects of dietary bile acids on growth,body composition and lipid metabolism of juvenile turbot (Scophthalmusmaximus) at different lipid levels [J].OceanologiaetLimnologiaSinica,2014,45(3):617-625 (in Chinese with English abstract).
[24] FOLCH J,LEES M,SLOANE-STANLEY G H.A simple method for the isolation and purification of total lipids from animal tissues[J].TheJournalofBiologicalChemistry,1957,226(1):497-509.
[25] TURCHINI G M,TORSTENSEN B E,WING-KEONG N.Fish oil replacement in finfish nutrition[J].ReviewsinAquaculture,2009,1(1):10-57.
[26] YOONM.The role of PPARα in lipid metabolism and obesity:focusing on the effects of estrogen on PPARα actions[J].PharmacologicalResearch,2009,60(3):151-159.
[27] FUCHS C,KIENBACHER C,KERSHAW E E,etal.P357impaired PPARα signalling influences bile acid homeostasis in ATGL liver KO mice[J].JournalofHepatology,2014,60(1):S186.
[28] MARRAPODIM,CHIANG JY L.Peroxisome proliferator-activated receptor α (PPARα) and agonist inhibit cholesterol 7α-hydroxylasegene( CYP7A1) transcription[J].JournalofLipidResearch,2000,41(4):514-520.
[29] 宋永康,林 虬,陈人弼.鱼粉脂肪酸组成的分析研究[J].台湾海峡,2004,23(1):88-93.
SONG Y K,LIN Q,CHEN R B.Study on fatty acid composition in fishmeals by gas-chromatography [J].JournalofOceanographyinTaiwanStrait,2004,23(1):88-93 (in Chinese with English abstract).
(责任编辑:顾玉兰 Responsible editor:GU Yulan)
High-lipid Diet Supplemented with Bile Acids Affects the Tissue Fatty Acid Profile in Grass Carp,Ctenopharyngodonidella
CHEN Haojie,TIAN Jingjing,ZHAO Jishu and JI Hong
(College of Animal Science and Technology,Northwest A&F University,Yangling Shaanxi 712100,China)
To investigate the effects of high-lipid diet supplemented with bile acids on tissue fatty acid profile in grass carp,three isonitrogenous practical diets(soybean oil served as the primary lipid source),including control group (CT,protein 350 g/kg,lipid 50 g/kg),high-lipid group (HL,protein 350 g/kg,lipid 70 g/kg) and bile-acids group (BA,protein 350 g/kg,lipid 70 g/kg,bile acids 60 mg/kg),were formulated to feed 96 grass carp (initial body mass:69.86 g±6.24 g) for 8 weeks. The fatty acid profiles of tissues reflected those of the diets. BA group showed lower mass fraction of total fatty acids,n-6 polyunsaturated fatty acid (PUFA) and linoleic acid (LA,18∶2n-6) in the lipid of intraperitoneal fat,compared with HL group,with an inverse trend in the lipid of hepatopancreas. BA group increased the mass fraction of n-3 PUFA in the total fatty acids in the hepatopancreas,but decreased in the muscle,compared with HL group.Besides,in the lipid of intraperitoneal fat,the mass fraction of saturated fatty acid (SFA) in BA group was lower than that in HL group,whereas those in the lipid of hepatopancreas were similar among all groups. BA group showed the highest tissue-diet fatty acid correlation,followed by HL group,CT group was the lowest. Overall,bile acids might regulate the transportation and accumulation of fatty acids in the tissues. It also increased the fatty acid correlation between tissues and diets.
High-lipid; Bile acids;Ctenopharyngodonidella; Fatty acid profile.
CHEN Haojie,male,master student. Research area: aquatic animal nutrition and feed science. E-mail: chj19910209@foxmail.com
JI Hong,male,professor,doctoral supervisor. Research area: aquatic animal nutrition and feed science. E-mail: jihong@nwsuaf.edu.cn
2016-04-18
2016-05-12
陈昊杰,男,硕士研究生,研究方向为水产动物营养与饲料学。E-mail: chj19910209@foxmail.com
吉 红,男,教授,博士生导师,研究方向为水产动物营养与饲料学。E-mail: jihong@nwsuaf.edu.cn
日期:2016-12-20
S963.73+9
A
1004-1389(2017)01-0014-11
网络出版地址:http://www.cnki.net/kcms/detail/61.1220.S.20161220.1640.004.html
Received 2016-04-18 Returned 2016-05-12
