基于地形因素的高寒草甸土壤温湿度和物种多样性与初级生产力关系研究
2017-03-09牛钰杰周建伟杨思维王贵珍杜国祯花立民
牛钰杰,周建伟,杨思维,王贵珍,刘 丽,杜国祯,花立民,*
1 甘肃农业大学 草业学院/草业生态系统教育部重点实验室/中-美草地畜牧业可持续发展研究中心, 兰州 730070 2 兰州大学生命科学学院, 兰州 730030
山地是青藏高原的典型地形[1]。作为特殊的生境岛屿,山体地形决定了水热资源的再分配并影响着物种多样性[2]。高寒草甸作为青藏高原最主要草地类型,主要分布于山地之间[1],地形是物种多样性和土壤水热资源变化的关键非生物因子。不同地形下草地物种多样性和土壤水热与生态系统功能的关系成为当今生态学研究热点之一[3- 4],初级生产力作为综合反映生态系统服务功能的指标得到生态学家广泛认可[5],物种多样性作为丰富度[6]、多度[7]、优势度[8]和异质性测度[9]的综合统计量,不同多样性大小,代表群落物种间不同组成结构。植物群落在漫长的演化过程中形成的动态稳定群落结构,是对土壤水热等环境因子的综合响应[10]。水热资源分配成为影响植物多样性进而导致生产力变化的主要原因[10]。因此,生态系统初级生产力受到群落物种组成和土壤因素的共同影响[11- 12]。
山地坡向和海拔是山地高寒草甸最主要的地形因素,直接影响太阳辐射和降水的空间再分配[13],进而导致土壤水分和温度的差异性分布[14]。国内外学者在开展生产力和多样性关系研究时,一是通过较大地理尺度和组织层次的研究来消除地形因素的干扰[3,10],忽略了小尺度山体单元中土壤温湿度和群落多样性的具体差异;二是通过小尺度均质生境和人工群落来模拟研究[11,15],忽略了由于尺度增大导致的较强空间异质性对土壤水热和多样性与生产力关系的影响。三是在山地研究中,只对山体不同坡向或海拔进行同一空间层次水平上的研究,取样在不同山体间进行,不能对山地整体的空间层次或坡向和海拔层次进行系统研究[12,16]。因此,这些研究得出了不一致的研究结果,且彼此间参考性不高[3,10,15,17]。尽管已有的小尺度山体生态学研究表明,地形是生境中土壤温湿度异质性分布的主要影响因素,且得出坡向是物种多样性和土壤含水量差异分布的主导因素[16,18]。但土壤温湿度和物种多样性的哪一组分对高寒草甸生态系统山地整体层次以及局部不同坡向和海拔层次下的生产力影响更为明显,仍然不甚明确[17,19- 20]。小尺度山体作为山地草甸生态系统管理的最基本地形单元,消除了地理区域不同带来的气候差异,成为坡向与海拔共同作用下水热资源和物种多样性与生产力关系研究的理想区域。故本研究选取青藏高原东北缘祁连山东缘马牙雪山支脉的代表性小尺度高寒草甸山体为研究对象。排除地理因素差异的干扰,只考虑坡向和海拔两维变量,进行土壤温湿度和物种多样性与初级生产力关系的研究,并从整个支脉山体以及坡向和海拔两个层次,研究土壤温湿度和生物多样性哪一个组分对初级生产力的影响更为明显这一科学问题,本研究结果希望能够为大尺度山地生态学研究提供参考,并为科学、合理地利用小尺度山体高寒草甸提供科学依据。
1 材料与方法
1.1 研究区概况
本研究选取青藏高原东北缘甘肃省天祝藏族自治县祁连山东缘马牙雪山一个支脉山体,沿山脊线从山底到山顶的长度3.2 km,海拔2850—3020 m。平均坡度为8—10°,草地面积大约为150 hm2。草地类型为典型高寒草甸。地理坐标为37°09′—37°14′N,102°40′—102°47′E,年均气温-0.5℃,>0℃的年积温为1380℃。近5年来,年均降水量416 mm,主要集中在7—9 月,占全年降雨量的76%,年潜在蒸发量1592 mm。无绝对无霜期,植物生长期120—140 d。土壤类型为高山草甸土,土层厚度40—50 cm。不同坡向植被类型均属于高寒草甸类,但因为微生境不同,又存在明显群落差异,阴坡为珠芽蓼(Polygonumviviparum)+线叶嵩草(Kobresiacapillifolia)群落,山脊为线叶嵩草(Kobresiacapillifolia)+球花蒿(Artemisiasmithii)群落,阳坡为洽草(Koeleriacristata)+赖草(Leymussecalinus)群落,阶地为垂穗披碱草(Elymusnutans)+紫花针茅(Stipapurpurea)群落。
1.2 试验设计
系统考察研究区域内山地植被和地形,选取位于祁连山东缘、坡度和年际放牧强度基本一致的整个高寒草甸支脉山体为研究对象。消除大气候、放牧和坡度的干扰。将整个山地划分为4个样区,阴坡(Shadow aspect,SHA)、山脊(Ridge,RI)和阳坡(Sunny aspect,SUA)、阶地(Bottom land,BL)。阶地海拔2850 m,系统将试验山地海拔平均分为3个海拔段,低海拔区(2860—2910 m)(Low elevation,LO)、中海拔区(2910—2960 m)(Medium elevation,ME)、高海拔区(2960—3020 m)(High elevation,HI)(图1)。采用大样本系统取样的方法,从山底开始,以垂直于山脊线的方向,设置样带(包括阴坡、山脊和阳坡)。海拔每上升5 m,设置一条样带,直到山顶,共设置24条样带。在每条样带的阴坡和阳坡的上、中、下坡位各设置1个样方,在山脊设置1个样方,1条样带内共7个样方,山体共168个样方。在阶地位置平行设置3条样带,每个样带7个样方,共21个样方。本研究中,样方大小0.5 m×0.5 m,总计样方189个。调查所有样方内植物群落结构、土壤温湿度。选取物种丰富度、多度、均匀度和优势度反映物种多样性。首先以整个山体为研究对象,分析土壤温湿度和物种多样性对初级生产力(用生物量表示)的影响。再进一步考虑山体的不同坡向和海拔,研究土壤温湿度和物种多样性与初级生产力的关系。

图1 研究样地设置Fig.1 Design of the experiment plot
1.3 植物群落结构调查与土壤温湿度测定
于2012年8月中旬植物旺盛生长期进行植被群落调查。在研究样地用系统取样的方法开展植物群落调查和土壤温湿度测定。调查内容和方法包括,群落物种丰富度用0.25 m2样方,以其中出现的物种数表示该样方物种丰富度;群落内物种盖度用0.25 m2样方,针刺100次,记录每一物种盖度;群落内物种频度在每个样方附近随机抛样圆20次,记录该样点每一物种出现频率。分物种将植株地上部分齐地面剪取,并记录每一物种的个体数量,表示该物种的多度。在实际调查取样时,密丛型以不可再分的有主茎植物属于一株。而匍匐茎型植物按已扎根植物算为一株。样方内剪取的植物装入信封带回实验室后置于70℃烘箱内烘干至恒重,获得干重,以表示地上生物量。使用群落物种盖度和频度的平均值表示物种重要值,用最大值标准化法计算物种相对重要值。计算样方群落的物种均匀度(Evenness指数)[9]和优势度(Berger-Parker指数)[8]。计算公式为:
Berger-Parker指数:
B=Nmax/N
式中,S为每个样方总的物种数;N为S个全部物种的相对重要值之和;Ni为第i个物种的相对重要值,Nmax为相对重要值最大值。
在植物群落调查的样方内,用托普TSZ-IIW型土壤水分温度测量仪测定0—10、10—20 cm和20—30 cm土壤温湿度,重复5次的平均值表示该样方点该土层深度的温湿度,将该样方点3层土壤的温湿度取平均值,表示该样方表层土壤(0—30 cm)的温湿度。并用Garmin手持GPS记录样方点的经纬度和海拔。
1.4 数据分析
首先对物种丰富度、多度、均匀度、优势度、土壤温度和湿度做不同坡向和海拔下样方间的多重比较分析(LSD法)。采用线性回归法分析土壤温湿度、物种多样性(丰富度、 多度、均匀度、优势度)与初级生产力之间的关系。为了排除土壤温湿度和物种多样性之间的相互干扰,进而影响与初级生产力的关系,一是控制土壤温湿度,对物种多样性与生产力进行偏相关分析,设定P值为0.05。二是控制物种多样性,对土壤温湿度与生产力进行偏相关分析。在排除某一因子影响后,用偏相关决定系数R2,解释土壤温湿度和物种多样性对初级生产力的贡献大小。所有统计分析均使用SPSS 19.0软件进行。
2 结果与分析
2.1 山地土壤温湿度、物种多样性和初级生产力比较
样方调查结果显示(表1),土壤温度在中等海拔的阳坡最高,土壤湿度在高海拔的阴坡最高,物种丰富度在中等海拔的阴坡最高,多度在高海拔的山脊最高,均匀度在低海拔的阳坡最高,优势度在高海拔的阴坡最高,初级生产力在中等海拔的阴坡最高。阴坡土壤温度低且变异系数小,湿度高且变异系数大,土壤温度与湿度显著负相关(R2=0.19P=0.00)。阳坡土壤湿度低且变异系数小,温度高变异系数大,温度与湿度无显著相关关系(R2=0.04P=0.11);山脊和阶地土壤温度与湿度相比阴阳坡处于中间水平,但阶地土壤温度与湿度显著负相关(R2=0.72P=0.00)(图2)。

表1 山地土壤温湿度、物种多样性和初级生产力比较
括号内为纵向比较,括号外为横向比较;显著水平:P<0.05

图2 不同样区位置及土壤温湿度关系Fig.2 Relationship between soil temperature and moisture in different sampling plots
2.2 整个山地土壤温湿度和物种多样性与初级生产力的关系
根据研究区189个样方数据建立的土壤温湿度、物种多样性与初级生产力之间的回归模型(图3)可知,对于整个山体,初级生产力随物种多度的增加而显著增加(R2=0.07P=0.01),其余指标均未与初级生产力表现出显著相关关系。

图3 山地土壤温湿度和物种多样性与初级生产力的关系Fig.3 Relationship between soil temperature, soil moisture, species diversity and primary productivity in the entire hill
2.3 不同坡向下土壤温湿度和物种多样性与初级生产力的关系
由图4可知,阴坡初级生产力随物种丰富度的增加而显著增加(R2=0.13P=0.00),其余多样性指标与初级生产力未表现显著相关关系。山脊初级生产力也随物种丰富度的增加而显著增加(R2=0.12P=0.02),其余多样性指标与初级生产力未表现显著相关关系。阳坡初级生产力随物种多度的增加而显著增加(R2=0.12P=0.04),其余多样性指标与初级生产力未表现显著相关关系。阶地初级生产力随均匀度的增加而显著增加(R2=0.28P=0.04),随优势度的增加而显著降低(R2=0.49P=0.03),其余多样性指标与初级生产力未表现显著相关关系。

图4 不同坡向物种多样性与初级生产力的关系Fig.4 Relationship between species diversity and primary productivity on different slope aspects of the hill
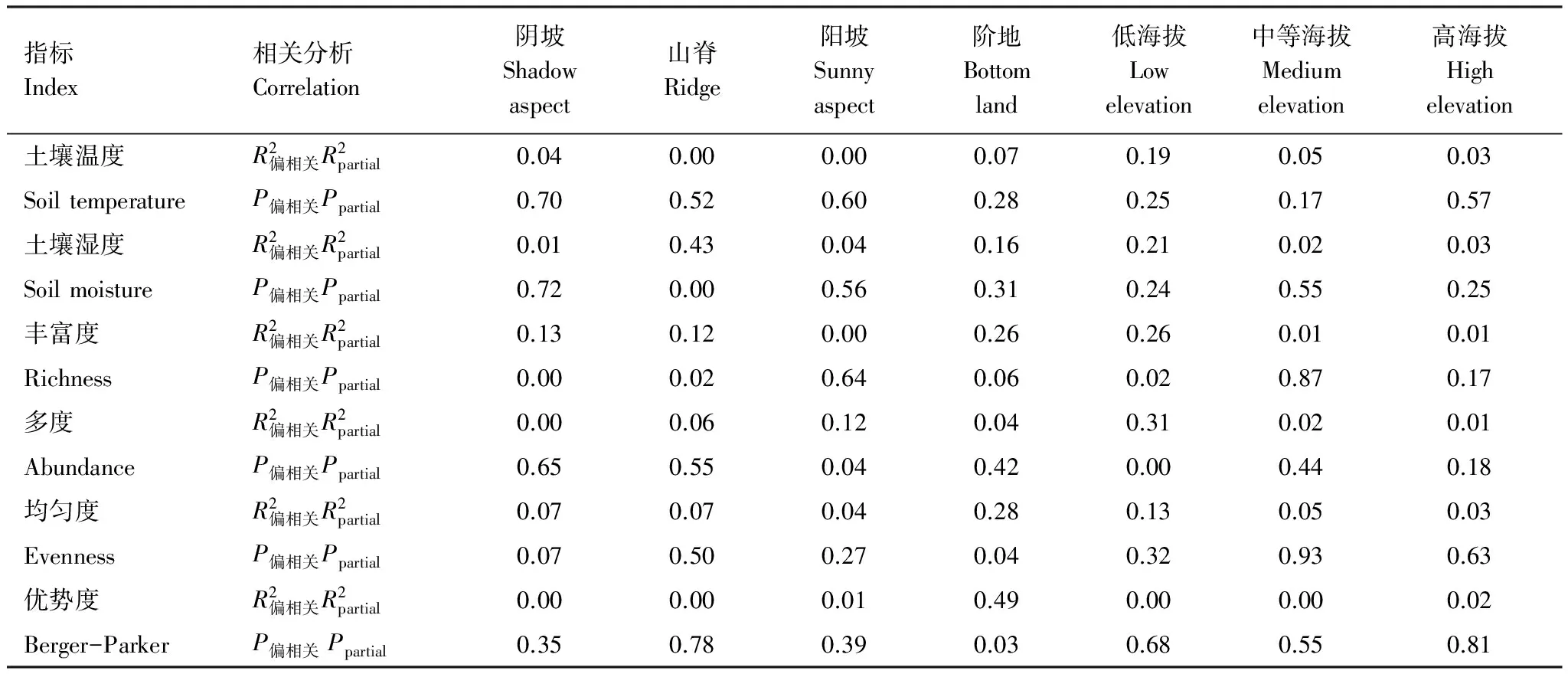
由图5可知,只有山脊初级生产力随土壤湿度的增加而显著增加,其余坡向均未与土壤温度和湿度表现显著相关关系。基于偏相关R2值可知(表2),除山脊初级生产力受土壤湿度的影响较大外,其余坡向均是物种多样性对群落初级生产力的影响较大。
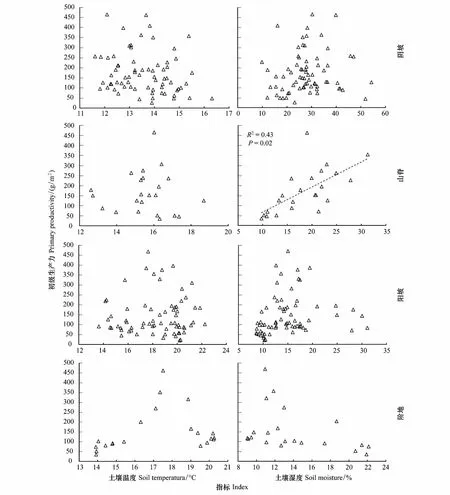
图5 不同坡向土壤温湿度与初级生产力的关系Fig.5 Relationship between soil temperature, soil moisture and primary productivity on different slope aspects of the hill
2.4 不同海拔段土壤温湿度和物种多样性与初级生产力的关系
由图6对于不同海拔段分析可知,低海拔区群落初级生产力随物种多度的增加而显著增加(R2=0.31P=0.00),随物种丰富度也显著增加(R2=0.19P=0.02),其余物种多样性指标未与初级生产力表现显著相关关系。在中等海拔区和高海拔区物种多样性指标均未与初级生产力表现显著相关关系。由图7可知,在不同海拔段土壤温湿度均未与群落初级生产力表现出显著相关关系。基于偏相关R2值可知(表2),除低海拔区初级生产力受物种多度和丰富度的影响较大外,其余海拔段土壤温湿度和物种多样性对群落初级生产力的影响均较低。
3 讨论与结论
3.1 山体层次物种多度是影响高寒草甸初级生产力变化的主要原因
本研究首先以山地高寒草甸整体为研究单元,发现初级生产力仅随物种多度的增加而显著增加。虽然土壤温湿度在不同的坡向和海拔表现出极强的空间变异性,但均未与初级生产力表现出显著相关关系。此结论与一些研究者在大尺度研究中指出的环境因子(水热资源)对生产力的影响强于物种多样性的观点不一致[11,19,21],究其原因,可能来自两个方面。第一,研究尺度大小的差异[22]。在环境梯度跨度较大的空间尺度下取样,水热条件发生极大变化,甚至可以忽略地形因素引起的差异,气候因素成为主导的影响因子,进而对生产力产生决定性作用。本研究,基于小尺度山体发现虽然不同坡向和海拔的土壤温湿度呈现出差异化的分布规律,但以山地整体为研究单元,土壤温湿度等因素对初级生产力的影响较弱。在小尺度山体,地形虽然引起了土壤水热因子的变化,但物种多样性的变化更为明显(表1),群落内物种组成结构的调整更为积极,更多的物种可以利用微小的环境异质性产生的资源差异而获得生存,群落内部物种的调整可以使不同的植物获取营养库中各自所需的营养,进一步表明小尺度山体高寒草甸群落初级生产力受生物群落内部调节的作用较大;第二,人类活动干扰,以往研究者往往在公路或铁路沿线进行取样[12],取样位置影响取样结果。放牧作为青藏高原草地最主要的人为干扰方式,在地理区域跨度较大的不同区域进行取样时,没有考虑放牧差异对物种多样性和生产力造成的干扰,进而影响其研究结果。在小尺度山地草甸研究中,不同的研究者在不同的坡向和海拔段得出不同的结果[10,12,15- 17]。但却缺乏跨坡向与海拔的山体尺度研究,因而不能在山地单元中得出整体概括性的研究结果。本研究以小尺度山地整体为研究单元进行大样本系统取样,避开大尺度取样的气候和差异性放牧影响,仅考虑地形引起的环境因子变化下土壤温湿度和多样性对初级生产力的影响,发现物种多度是造成初级生产力差异的最主要影响因素。物种多度的区域分布主要由环境资源特性、物种生态位和植物扩散过程等因素决定[23]。韩大勇和杨允菲[24]研究认为在生境尺度上,物种多度和物种分布呈显著正相关关系。即局部多度高的物种一般趋于广布,而局部多度低的物种趋于受限分布[23]。植物多度可以实现不同生境间的连接。本研究在生境尺度上进行,植物群落属于局域种群,研究区以密丛型莎草科和根茎疏丛型禾本科植物分布为主,山地生境资源可以提供极高物种多度的共存,且相互竞争达到动态稳定,因此,在山体层次,植物多度可以跨不同坡向与海拔生境影响初级生产力的发挥。
表2不同坡向和海拔下土壤温湿度、物种多样性与初级生产力的关系
Table2Relationshipsbetweensoiltemperature,soilmoisture,speciesdiversityandprimaryproductivityondifferentslopeaspectswithdifferentaltitudeofthehill
指标Index相关分析Correlation阴坡Shadowaspect山脊Ridge阳坡Sunnyaspect阶地Bottomland低海拔Lowelevation中等海拔Mediumelevation高海拔Highelevation土壤温度R2偏相关R2partial0.040.000.000.070.190.050.03SoiltemperatureP偏相关Ppartial0.700.520.600.280.250.170.57土壤湿度R2偏相关R2partial0.010.430.040.160.210.020.03SoilmoistureP偏相关Ppartial0.720.000.560.310.240.550.25丰富度R2偏相关R2partial0.130.120.000.260.260.010.01RichnessP偏相关Ppartial0.000.020.640.060.020.870.17多度R2偏相关R2partial0.000.060.120.040.310.020.01AbundanceP偏相关Ppartial0.650.550.040.420.000.440.18均匀度R2偏相关R2partial0.070.070.040.280.130.050.03EvennessP偏相关Ppartial0.070.500.270.040.320.930.63优势度R2偏相关R2partial0.000.000.010.490.000.000.02Berger-ParkerP偏相关Ppartial0.350.780.390.030.680.550.81
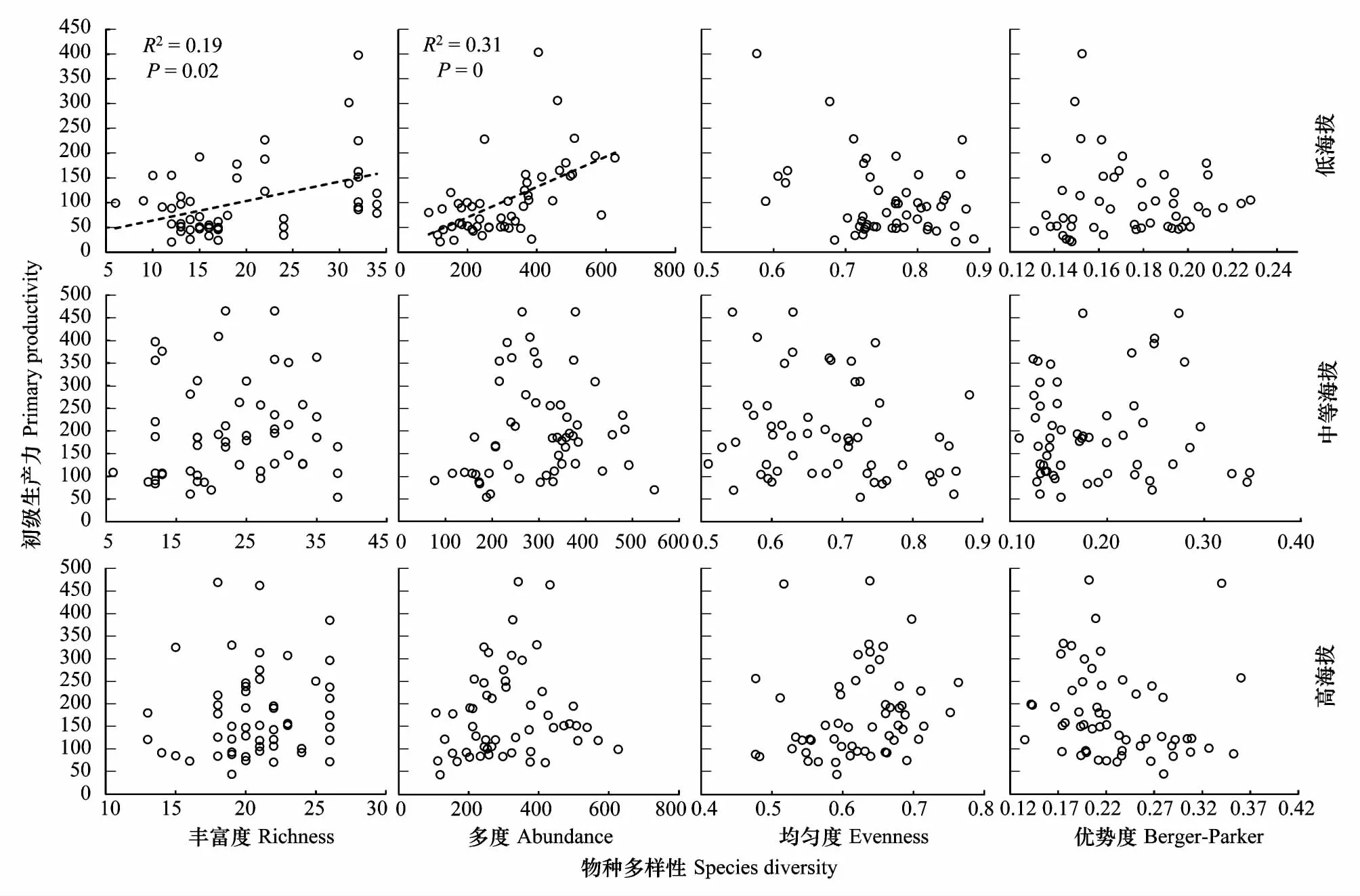
图6 不同海拔物种多样性与初级生产力的关系Fig.6 Relationship between species diversity and primary productivity in different elevation gradients
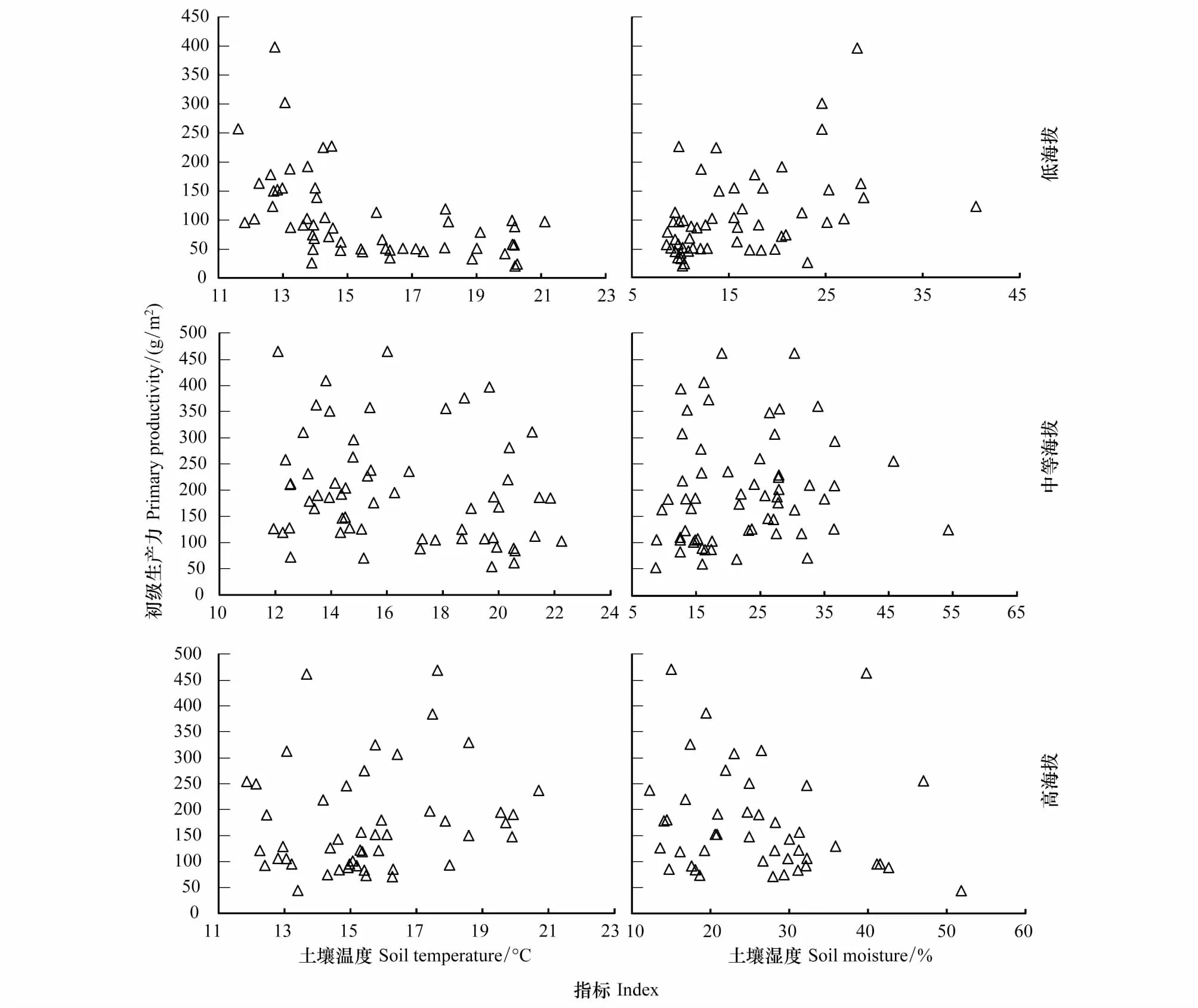
图7 不同海拔段土壤温湿度与初级生产力的关系Fig.7 Relationship between soil temperature, soil moisture and primary productivity in different elevation gradients
3.2 海拔和坡向层次对初级生产力的影响因素各异
3.2.1 坡向对土壤温湿度和物种多样性与初级生产力关系的影响较海拔更大
在自然生态系统中,物种多样性和生产力均极大地受到非生物因素的影响[18],而土壤温湿度是最主要的非生物因素。李新娥[16]发现坡向导致的土壤含水量差异性分布是限制植物生长和分布的主要因子。本研究也表明,在不同坡向,土壤温湿度和物种多样性均能不同程度对生产力产生积极作用。但对于海拔梯度而言,只有低海拔区初级生产力随物种多度和丰富度的增加而增加,其余海拔段初级生产力与土壤温湿度和物种多样性无关。本研究中海拔的梯度跨度只有170 m,土壤温湿度随坡向发生较大改变,植物群落组成也因坡向不同而显著变化。但随海拔升高气温的直减率只有0.5—0.7℃/100 m[25],海拔对土壤温湿度和群落结构造成的影响相对较小。在小尺度山体,坡向造成了土壤水热供给和物种多样性分布的较大差异。因此,在实际山体尺度高寒草甸生态系统生产和生态恢复过程中要重点考虑坡向不同造成的土壤温湿度和植物多样性对生产力的影响,进而对生态系统生产力的发挥提供积极帮助。
3.2.2 不同坡向对初级生产力的影响因素各异
土壤水热因子和物种多样性是影响高寒草甸初级生产力变化的关键因素。以往聚焦于物种多样性与初级生产力关系的研究,常以物种丰富度作为多样性的代表性测度指标[26],发现其关系主要有线性[10]、单峰[15]和无关系[17]3种情况。而多样性组分的不同反映了生物群落在组成、结构、功能和动态方面的差异。土壤水热资源供给也被认为是导致多样性与生产力关系变化的主要原因。马文静等[10]发现多样性和生产力关系的发挥受到不同水热资源供给程度的影响。赵路强和王刚[27]研究认为在高寒草甸不同区域生境的局域种群内生产力受不同多样性指数的影响程度不同。而在同一山体不同坡向和海拔下,植物群落组成和土壤温湿度已发生极大变化。本研究中,阴坡初级生产力与物种丰富度正线性相关,不同物种在环境资源利用上存在差异,且阴坡水分供应充足,物种数的增加有利于物种在不同时间、空间上,以不同的方式对资源进行利用,使资源获得最大的利用效率,从而提高生态系统生产力[10]。山脊初级生产力主要随土壤湿度的增加而增加,也受物种丰富度的显著影响。山脊由于其特殊的地理位置,水分难以保持,土壤湿度的变异系数较大,生产力主要受土壤湿度的影响。阳坡初级生产力与物种多度正线性相关,阳坡土壤温度高,湿度低,在有限资源的基础上很难维持较高数量物种的共存,但物种数的降低就必须通过增加现有种的多度来补偿。阶地初级生产力随均匀度的增加而显著增加,随优势度的增加而显著降低。阶地作为土壤养分供给最充足和异质性最小的区域,不同的物种可以在此共存,优势物种的优势降低,物种分布的越为均匀,则群落的整体生产力越高,这与部分学者在可控均质草地的实验结果一致[11]。因此,在不同水热资源供给下,多样性与生产力关系的研究中,应重点考虑具体生境类型的差异和空间尺度变化所带来的影响。
[1] 鲁春霞, 谢高地, 肖玉, 于云江. 青藏高原生态系统服务功能的价值评估. 生态学报, 2004, 24(12): 2749- 2755.
[2] 马克明, 张洁瑜, 郭旭东, 傅伯杰. 农业景观中山体的植物多样性分布: 地形和土地利用的综合影响. 植物生态学报, 2002, 26(5): 575- 588.
[3] Bai Y F, Wu J G, Pan Q M, Huang J H, Wang Q B, Li F S, Buyantuyev A, Han X G. Positive linear relationship between productivity and diversity: evidence from the Eurasian Steppe. Journal of Applied Ecology, 2007, 44(5): 1023- 1034.
[4] 董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威. 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 2017, 37(5): 1472- 1483.
[5] Cardinale B J, Wright J P, Cadotte M W, Carroll I T, Hector A, Srivastava D S, Loreau M, Weis J J. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(46): 18123- 18128.
[6] McIntosh P P. An index of diversity and the relation of certain concepts to diversity. Ecology, 1967, 48(3): 392- 404.
[7] Tokeshi M. Species abundance patterns and community structure. Advances in Ecological Research, 1993, 24: 111- 186.
[8] Magurran A E. Ecological Diversity and Its Measurement. Netherlands: Springer, 2013.
[9] Smith B, Wilson J B. A consumer′s guide to evenness indices. Oikos, 1996, 76(1): 70- 82.
[10] 马文静, 张庆, 牛建明, 康萨如拉, 刘朋涛, 何欣, 杨艳, 张艳楠, 邬建国. 物种多样性和功能群多样性与生态系统生产力的关系——以内蒙古短花针茅草原为例. 植物生态学报, 2013, 37(7): 620- 630.
[11] 贺金生, 方精云, 马克平, 黄建辉. 生物多样性与生态系统生产力: 为什么野外观测和受控实验结果不一致? 植物生态学报, 2003, 27(6): 835- 843.
[12] 李凯辉, 胡玉昆, 王鑫, 范永刚, 吾买尔·吾守. 不同海拔梯度高寒草地地上生物量与环境因子关系. 应用生态学报, 2007, 18(9): 2019- 2024.
[13] 黄瑞灵, 周华坤, 刘泽华, 雷占兰, 姚步青, 汪新川, 王文颖, 赵新全. 坡向与海拔对青海省拉鸡山不同植被土壤种子库的影响. 生态学杂志, 2013, 32(10): 2679- 2686.
[14] Burt T P, Butcher D P. Topographic controls of soil moisture distributions. Journal of Soil Science, 1985, 36(3): 469- 486.
[15] 陈超, 朱志红, 李英年, 姚天华, 潘石玉, 卫欣华, 孔彬彬, 杜家丽. 高寒草甸种间性状差异和物种均匀度对物种多样性与功能多样性关系的影响. 生态学报, 2016, 36(3): 661- 674.
[16] 李新娥. 亚高寒草甸阳坡—阴坡梯度上植物功能性状及群落构建机制研究[D]. 兰州: 兰州大学, 2011.
[17] Mittelbach G G, Steiner C F, Scheiner S M, Gross K L, Reynolds H L, Waide R B, Willig M R, Dodson S I, Gough L. What is the observed relationship between species richness and productivity? Ecology, 2001, 82(9): 2381- 2396.
[18] 刘旻霞, 马建祖. 甘南高寒草甸植物功能性状和土壤因子对坡向的响应. 应用生态学报, 2012, 23(12): 3295- 3300.
[19] Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science, 2001, 294(5543): 804- 808.
[20] 陈生云, 赵林, 秦大河, 岳广阳, 任贾文, 李元寿, 赵拥华. 青藏高原多年冻土区高寒草地生物量与环境因子关系的初步分析. 冰川冻土, 2010, 32(2): 405- 413.
[21] 杨元合, 饶胜, 胡会峰, 陈安平, 吉成均, 朱彪, 左闻韵, 李轩然, 沈海花, 王志恒, 唐艳鸿, 方精云. 青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系. 生物多样性, 2004, 12(1): 200- 205.
[22] 袁自强, 魏盼盼, 高本强, 张荣. 取样尺度对亚高寒草甸物种多样性与生产力关系的影响. 植物生态学报, 2012, 36(12): 1248- 1255.
[23] 马克明. 物种多度格局研究进展. 植物生态学报, 2003, 27(3): 412- 426.
[24] 韩大勇, 杨允菲. 松嫩草地植物群落物种多度——分布关系及其解释. 生物多样性, 2014, 22(3): 348- 357.
[25] 方精云, 沈泽昊, 崔海亭. 试论山地的生态特征及山地生态学的研究内容. 生物多样性, 2004, 12(1): 10- 19.
[26] Gotelli N J, Colwell R K. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters, 2001, 4(4): 379- 391.
[27] 赵陆强, 王刚. 甘南高寒草甸物种多样性与生产力的关系. 兰州大学学报: 自科科学版, 2009, 45(6): 82- 86, 93- 93.
