基于线粒体CO Ⅰ基因序列对中国海蛾螺科的分子系统学分析
2017-02-14张树乾张素萍
张树乾,张素萍*
(1. 中国科学院 海洋研究所,山东 青岛266071)
基于线粒体CO Ⅰ基因序列对中国海蛾螺科的分子系统学分析
张树乾1,张素萍1*
(1. 中国科学院 海洋研究所,山东 青岛266071)
本研究利用18种蛾螺总科(Buccinoidea)动物的CO Ⅰ基因序列对中国海蛾螺科进行了科上和科内水平的分子系统学分析。在建立的系统发育树中,蛾螺科种类聚成了4个明显的分枝:第一枝包括侧平肩螺(Japelionlatus)、朝鲜蛾螺(Buccinumkoreana)、黄海蛾螺(Buccinumyokomaruae)、皮氏蛾螺(Voluthapaperryi)、香螺(Neptuneacumingi)、小鼠脊蛾螺(Lirabuccinummusculus)以及褐线蛾螺(Japeuthriacingulata);第二枝包括莫氏角亮螺(Antillophosmonsecourorum)和尖鱼篮螺(Nassariaacuminata);第三枝包括黑口甲虫螺(Cantharusmelanostoma)和矛唇齿螺属(Enginalanceolata);第四枝包括褐管蛾螺(Siphonaliaspadicea)和长吻真螺(Euthrialubrica)。该结果与先前基于形态分析进行的亚科划分基本一致。蛾螺科的单系性没有得到支持,织纹螺科种类(Nassariusthachi、N.sinarus、N.euglyptus)插到蛾螺科系统树中并与角亮螺属(Antillophos)及鱼篮螺属(Nassaria)聚成一枝;东风螺科种类(Babyloniaareolata、B.spirata)和榧螺科种类(Olivamustelina)与褐线蛾螺(Japeuthriacingulata)聚成一枝。东风螺属起初隶属于蛾螺科,现已被移出并作为单独的一科——东风螺科Babyloniidae,本研究确认了这一结论。鱼篮螺属(Nassaria)和角亮螺属(Antillophos)与织纹螺属(Nassarius)种类聚在一起,这与最近的分子研究得到的结果一致。
腹足纲;蛾螺科;系统发育;CO Ⅰ基因
1 引言
蛾螺科(Buccinidae)隶属于软体动物门(Mollusca)、腹足纲(Gastropoda)、新腹足目(Neogastropoda)。其种类繁多、形态多变,且分布广泛,从热带海域到两极地区、从海水到淡水、从潮间带至数千米的海底深渊,都有它们的踪迹。最早的蛾螺科化石种发现于下白垩纪时期(late Cretaceous),因此被看作为新腹足目中最古老的类群。经过漫长的适应辐射,蛾螺科已经进化成为新腹足目中最成功的类群之一[1]。到目前为止,全世界共发现和报道蛾螺科种类1 500余种,隶属于150属[2]。
尽管关于蛾螺科种类的研究和报道可以追溯至林奈时期,但到目前为止,蛾螺科仍然缺乏稳定、统一的分类系统,甚至在每个分类阶元上都存在着争议和混乱。一些学者认为,织纹螺科(Nassariidae)、细带螺科(Fasciolariidae)及盔螺科(Melongenidae)与蛾螺科(Buccinidaes.s.)在解剖结构上非常相似,并没有充分可靠的形态特征可以把它们分到科的水平上,因此将这几个科看作蛾螺科(Buccinidaes.l.)的4个亚科[3—4]。而在最新的分类系统中,这4个亚科被看作4个独立的科[2]。在科内水平上,蛾螺科种类各属、种间的系统演化关系仍然不够明确。Harasewych[5]认为,由于蛾螺科形态特征的复杂性,有接近200个种在分类上是不明确的。一些学者对蛾螺科进行了亚科水平上的划分,如Vaught[6]根据贝壳的形态特征将蛾螺科分为4个亚科:Buccininae、Volutopsiinae、Photinae以及Pisaniinae;而Higo等[7]则认为蛾螺科应该分为10个亚科:Beringiinae、Volutopsiinae、Parancistrolepisinae、Ancistrolepisinae、Neptuneinae、Photinae、Siphonaliinae、Pisaniinae、Babyloniinae以及Buccininae。这些亚科的划分主要依靠贝壳外部形态和齿舌结构,而贝壳表面的颜色、雕刻以及厣的形态,容易受到环境的影响,具有较大的不稳定性;齿舌在不同的年龄、性别中存在着差异[8]。Cernohorsky[9]认为,齿舌在科内水平上并不能作为有效的分类依据。张树乾和张素萍[10]通过观察对比蛾螺科的齿舌后发现,蛾螺科种类的齿舌在各属间存在着差异,可以作为区分属和种的有效依据。
Kantor[11]认为鱼篮螺属Nassaria种类胃的外部形态与织纹螺种类相似,最近的分子研究表明,鱼篮螺属(Nassaria)和角亮螺属(Antillophos)与织纹螺科种类聚在一起,而与其他蛾螺科种类具有相对较远的亲缘关系[12]。传统上,东风螺属(Babylonia)依据其贝壳特征被归入在蛾螺科内,但相关的解剖和分子研究表明,东风螺种类与榧螺有更近的亲缘关系,而与蛾螺科种类具有较远的亲缘关系[13—15]。
尽管分子生物学被越来越多地应用到腹足类的系统发育分析上,但关于蛾螺科的系统发育学研究仍然很少,且大多为一些区域性的研究[14—18]。本研究将利用18种蛾螺总科(Buccinoidea)动物的CO Ⅰ基因序列对中国海域分布的蛾螺科种类进行科上和科内水平的分子系统学分析。
2 材料和方法
本研究所用标本包括采自于中国海域的蛾螺科种类12属13种、织纹螺科1属3种、东风螺科1属1种,共计22个个体,经70%酒精固定,-20℃冰箱保存备用。其余的基因序列来自GenBank(表1)。
2.1 DNA提取
切取蛾螺腹足肌肉约30 mg,使用TIANGEN海洋生物基因组DNA抽提试剂盒(北京,天根生化科技)提取基因组DNA,获得的总DNA置于-20℃保存备用。
2.2 PCR及测序
实验所用的引物为Folmer等[19]所设计引物:LCO1490(F):5’-GGTCAACAAATCATAAAGATATTGG-3’, HCO2198(R): 5’-TTAACTTCAGGGTGACCAAAAAATCA-3’。由华大基因公司合成。引物稀释浓度为10 μmol/L,-20℃保存备用。
PCR反应总体积为25 μL,其中模板DNA为50~80 ng,12.5 μL Taq Mix (北京康为世纪生物科技有限公司),引物(10 μmol/L)各0.5 μL,双蒸水加至总体积为25 μL。PCR反应程序为:95℃预变性3 min,95℃变性30 s,42~47℃退火30 s,72℃延伸1 min,共运行35个循环,最后70℃延伸10 min。PCR产物直接送交华大基因公司完成纯化、测序。
2.3 序列对比和对齐
利用MEGA 6.06软件中的Clustal X对所选取的CO Ⅰ基因序列进行序列对比,辅以人工校对。利用MEGA计算各序列的碱基组成和基于Kimura 2-parameter法的各属间、种间的遗传距离[20],并利用邻接法(Neighbor-joining,NJ)和最大似然法(Maximum Likelihood method,ML)构建系统发育树[21],1 000次重复获得自展检验值。
3 结果
3.1 碱基组成和遗传距离
利用CO Ⅰ引物均扩增出一条亮带,且带型一致,大小均在 600 bp 左右,符合预定结果。测序后,共获得CO Ⅰ基因片段22条。对研究所得的22条蛾螺总科种类线粒体CO Ⅰ基因片段与从GenBank下载所得的3条CO Ⅰ基因片段进行同源序列对齐和对比。保留序列长度为552~604 bp,序列无碱基插入和缺失。17条蛾螺科种类CO Ⅰ基因序列的平均碱基组成为:T:37.1%,C:17.7%,A:25.1%,G:20.1,A+T含量明显高于G+C含量,这一结果符合其他腹足类线粒体CO Ⅰ基因序列碱基的组成特点[22—23]。3个朝鲜蛾螺(Buccinumkoreana)个体间的G+C含量无差别,2个黄海蛾螺(Buccinumyokomaruae)个体间的G+C含量分别为37.9%和38.1%;其他蛾螺种类的G+C含量为36.2%~44.1%,其中尖鱼篮螺(Nassariaacuminata)的G+C含量最低,长吻蛾螺(Euthrialubrica)的G+C含量最高。在同源序列中,共有369个保守位点;235个变异位点,其中单突变子48个,简约信息位点187个。变异为转换和颠换,无插入和缺失。本研究中的碱基平均转换/颠换值(Ts/Tv)为1.78,说明尚未达到突变饱和[20]。
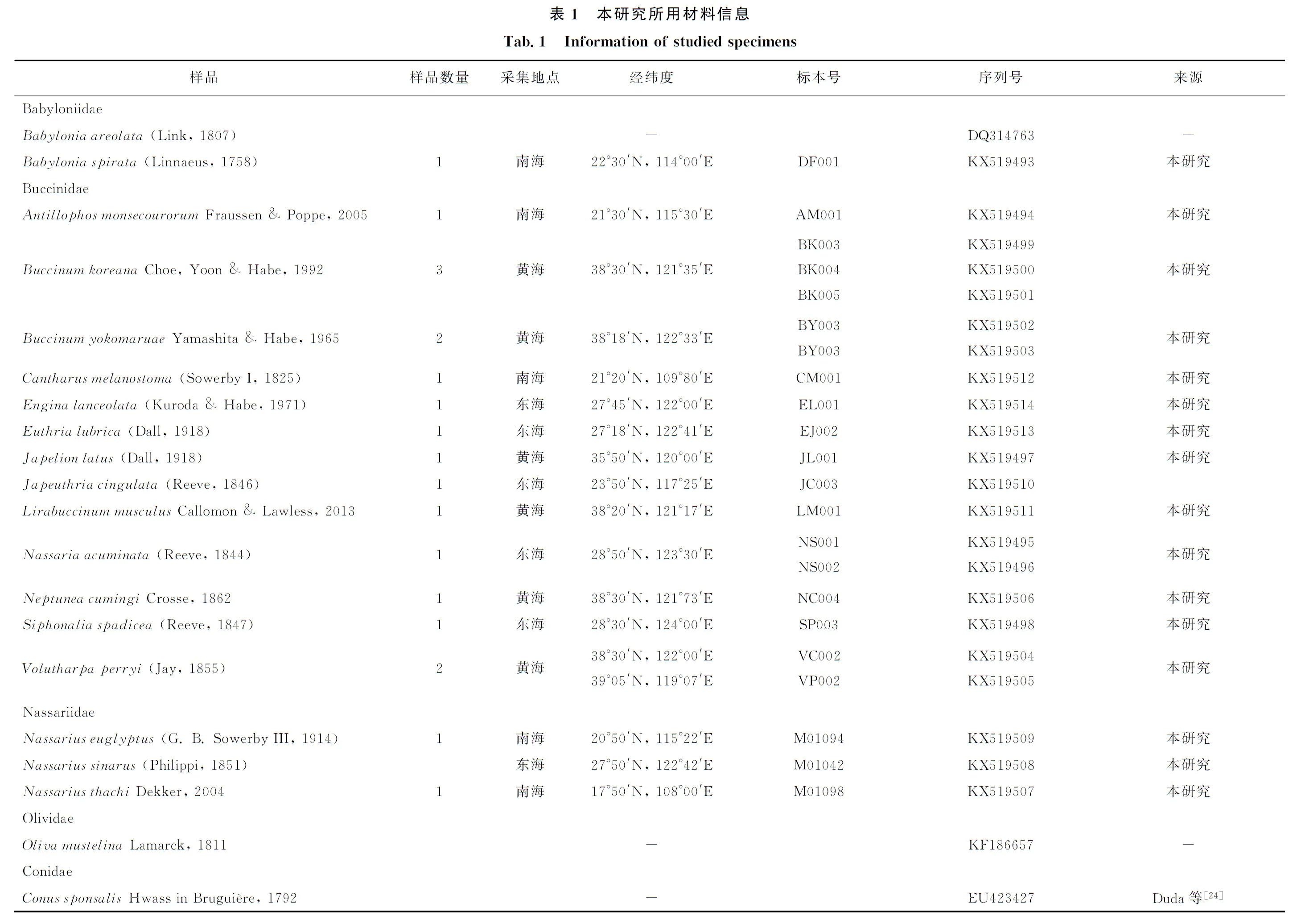
通过对比遗传距离表明,尖鱼篮螺(Nassariaacuminata)的种内遗传距离为0.022,朝鲜蛾螺(Buccinumkorena)的种内遗传距离为0~0.011,黄海蛾螺(Buccinumyokomaruae)的种内遗传距离为0.002;蛾螺科种类种间遗传距离为0.009~0.256,其中莫氏角亮螺(Antillophosmonsecourorum)与尖鱼篮螺(Nassariaacminata)之间的遗传距离最小,而尖鱼篮螺(Nassariaacuminata)与长吻真螺(Euthrialubrica)之间的遗传距离最大。蛾螺科种类与织纹螺科种类之间的遗传距离为0.162~0.268;蛾螺科种类与东风螺种类之间的遗传距离为0.190~0.285;东风螺种类与榧螺种类之间的遗传距离为0.197~0.227;东风螺种类与织纹螺种类之间的遗传距离为0.185~0.216(表2)。
3.2 序列分析
以芋螺科种类Conussponsalis为外群,采用邻接法(Neighbor-joining,NJ)和最大似然法(Maximum Likelihood method,ML)构建系统树,两种方法得到的结果基本一致(图1,图2):蛾螺科的单系性没有得到支持,织纹螺种类、东风螺种类和榧螺种类穿插到蛾螺科形成的枝序中,其中,3种织纹螺先与尖鱼篮螺和角亮螺聚在一起,再与其他蛾螺科种类相聚合;东风螺先与伶鼬榧螺聚成一枝,再与其他蛾螺科种类相聚合;蛾螺科聚成4枝:第一枝包括朝鲜蛾螺(Buccinumkoreana)、黄海蛾螺(Buccinumyokomaruae)、皮氏蛾螺(Volutharpaperryi)、侧平肩螺(Japelionlatus)、香螺(Neptuneacumingi)、小鼠脊蛾螺(Lirabuccinummuculus)和褐线蛾螺(Japeuthriacingulata);第二枝包括莫氏角亮螺(Antillophosmonsecourorum)和尖鱼篮螺(Nassariaacuminata);第三枝包括黑口甲虫螺(Cantharusmelanostoma)和矛唇齿螺(Enginalanceolata);第四枝包括褐管蛾螺(Siphonaliaspadicea)和长吻蛾螺(Euthrialubrica)。
4 讨论
4.1 (蛾螺亚科)蛾螺属+涡螺属+平肩螺属+香螺属+脊蛾螺属+线蛾螺属
本实验中,香螺属(Neptunea)种类和蛾螺属(Buccinum)种类聚成一枝,尽管置信度较低(小于50%)。Powell[25]认为香螺属(Neptunea)和蛾螺属(Buccinum)属于不同的亚科,香螺属属于香螺亚科(Neptuneinae)而蛾螺属属于蛾螺亚科(Buccininae),因为两者在厣和齿舌特别是厣的形态特征方面存在明显的区别,香螺属种类的厣为叶状,核在顶端,而蛾螺属种类的厣为椭圆形,核在中央或亚中央;Higo等[7]同样认为蛾螺属和香螺属应分别归入蛾螺亚科(Buccininae)和香螺亚科(Neptuneinae)中;Harasewych和Kantor[13]则认为,这两者除了在食道盲囊(oesophageal caecum)和莱布林腺(gland of Leiblein)的发达程度略有不同外,其他方面均很相似,两者应同属于蛾螺亚科。Hayashi[14]基于线粒体16S rRNA基因片段构建系统发育树后认为,这两属之间存在较近的亲缘关系。本实验支持了这一结论。本实验中,皮氏蛾螺(Volutharpaperryi)和蛾螺属种类聚在一起,且置信度较高(96%)。Smith[26]指出,皮氏蛾螺(Volutharpaperryi)软体部分头、触角以及眼点的位置都与蛾螺属种类非常地相似,说明两者之间存在较近的亲缘关系,本实验支持了这一观点。Vaught[6]将侧平肩螺归入蛾螺亚科中,张树乾和张素萍[10]通过观察侧平肩螺的齿舌发现,其侧齿和中央齿形态与蛾螺亚科其他种类很相似,两者应该有较近的亲缘关系,这一结论在本实验中得到了验证。小鼠脊蛾螺(Lirabuccinummuculus)的齿舌形态显示[10],其结构特征符合蛾螺亚科的齿舌形态,因而应归入蛾螺亚科中。线蛾螺属(Japeuthria)先与东风螺和榧螺聚在一起,再与其他的蛾螺亚科种类聚在一枝上。事实上,在去除东风螺和榧螺的情况下,线蛾螺属会直接与香螺属(Neptunea)聚在一起。褐线蛾螺(Japeuthriacingulata)的齿舌中央基板呈方形,后缘具6个齿尖,侧齿具3个齿尖[10],这些特征与其他蛾螺亚科种类特别是蛾螺属(Buccinum)和香螺属(Neptunea)的齿舌结构相似[27]。
4.2 (波特亚科)角亮螺属+鱼篮螺属
在本实验中,尖鱼篮螺(Nassariaacuminata)先与莫氏角亮螺(Antillophosmonsecourorum)聚在一起,再跟3个织纹螺聚成一枝。Cernohorsky[27]指出波特亚科(Photinae)种类的中央齿基板呈拱形,后缘具3个齿尖,而侧齿具2个齿尖。Cernohorsky[28]将鱼篮螺属(Nassaria)归入波特亚科中,尽管鱼篮螺属种类的齿舌中央齿基板后缘具5~10个齿尖。Galindo等[12]甚至认为,角亮螺属和鱼篮螺属应属于织纹螺科(Nasariidae)而非蛾螺科Buccinidae。事实上,角亮螺属最初归在织纹螺科[29—30],它们的齿舌形态与织纹螺更为接近。解剖学方面,Adams[31]指出,角亮螺属种类足部后端呈细丝状,与织纹螺种类的足部形态近似;Kantor[11]通过观察胃部形态后认为,鱼篮螺属种类胃的外部形态更接近织纹螺科而非蛾螺科。

4.3 (土产螺亚科)甲虫螺属+唇齿螺属
本实验中,黑口甲虫螺与矛唇齿螺聚在一起,且置信度较高(63%)。Cernorsky[27]指出唇齿螺属(Engina)与甲虫螺属(Cantharus)具有非常相似的外部形态结构,Vermeij[32]根据贝壳特别是壳口的形态将土产螺亚科划分为3个类群:唇齿螺群(Engina-group),土产螺群(Pisania-group)和甲虫螺群(Cantharus-group)。本实验得到的结果与Vermeij[32]得到的结果一致。
4.4 (管蛾螺亚科)管蛾螺属+真螺属
本实验中,管蛾螺属与真螺属聚在一枝上,尽管置信度较低(小于50%)。 Powell[25]根据贝壳和齿舌的形态把管蛾螺属(Siphonalia)归入到Siphonaliinae中;Higo等[7]接受了这一观点,但Vaught[6]将管蛾螺属归入到蛾螺亚科(Buccininae)中。Hayashi[14]利用16S rRNA构建的系统发育树显示,管蛾螺属没有与蛾螺亚科种类聚在一枝上,且置信度较高,说明其与蛾螺亚科种类之间存在着较远的亲缘关系。本实验支持了这一观点,因此管蛾螺属更倾向归入于管蛾螺亚科中。
Vaught[6]将真螺属(Euthria)归到蛾螺亚科(Buccininae)中,而Higo等[7]将真螺属作为土产螺亚科(Pisaniinae)下的一个属。本实验中,真螺属种类既没有与蛾螺亚科种类也没有与土产螺亚科种类聚在一起,而是与管蛾螺亚科中的褐管蛾螺聚在一枝上。形态学方面,真螺属与蛾螺亚科种类的贝壳形态和齿舌结构方面均存在明显的区别,真螺属种类的前水管沟很长,齿舌中央齿基板为三角铲形,而蛾螺亚科种类的前水管沟宽短,齿舌中央齿基板为方形;真螺属与土产螺亚科(Pisaniinae)亚科种类虽然在壳形上较为相似,但在齿舌结构上存在明显的区别[10];真螺属种类与管蛾螺属种类在贝类外形上较为相似(图1),且齿舌中央齿和侧齿上均具3个齿尖。因此,建议将真螺属归入管蛾螺亚科中。
4.5 关于东风螺属(Babylonia)和皮氏蛾螺的讨论
Kuroda等[33]为东风螺属(Babylonia)建立了东风螺亚科(Babyloniinae),隶属于蛾螺科(Buccinidae)。Harasewych和Kantor[13]通过研究发现,东风螺属种类与蛾螺科其他种类存在明显区别:东风螺属种类具有不成对的附属唾液腺(accessory salivary gland),一些种类中还具有直肠腺(rectal gland);齿舌中央齿基板外侧边缘具齿尖,且基板后缘的3个大齿尖聚集在中央位置。在根据分子数据建成的系统发育树上,Harasewych和Kantor[13]发现东风螺属首先与榧螺科种类聚在一起,然后才与蛾螺科相聚。根据这些证据,他们认为东风螺种类与蛾螺科种类具有较远的亲缘关系,因此将东风螺亚科提升到了科的水平上。本实验中,两个东风螺种类先与伶鼬榧螺聚在一起,其次才与蛾螺科种类相聚,支持了Harasewych和Kantor[13]的结论。
普通的皮氏蛾螺(见图1E-H)软体上通常具黑色花纹,作者于辽宁大连采得两个标本(见图1A-D),与普通皮氏蛾螺螺在壳形上非常相似,但在其软体上并不具黑色花纹或斑点、足宽大,前端为截形而非尖形。本研究中,其与普通的皮氏蛾螺聚在一起,置信度为100,且遗传距离为0.013,处于种内水平。证明两者为同一个种。
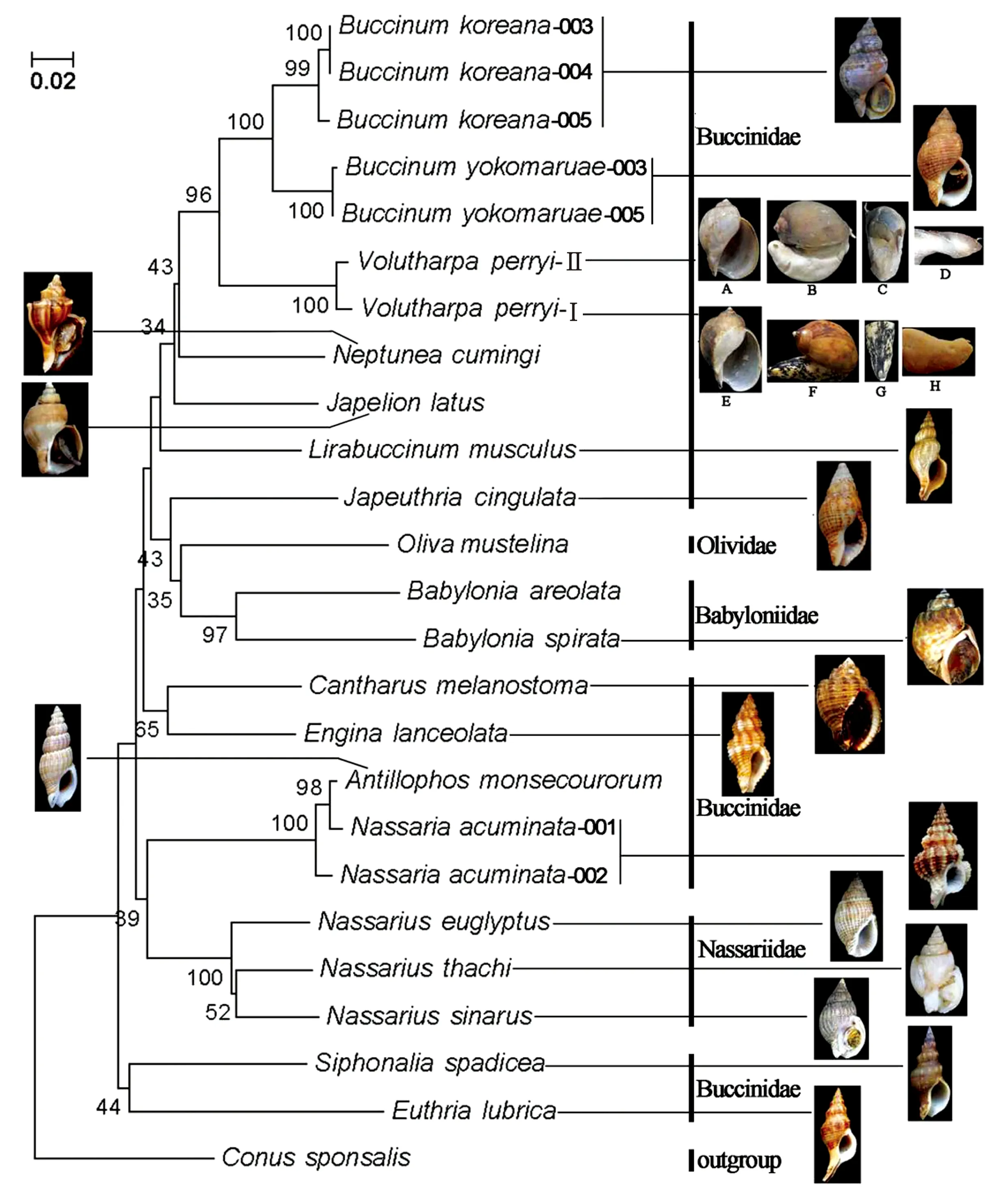
图1 基于CO Ⅰ基因序列片段的系统树(邻接法). A-D. 皮氏蛾螺(不带花纹);E-H. 皮氏蛾螺(带花纹)Fig.1 Phylogenetic tree based on CO Ⅰ gene sequences (neighbor-joining method). A-D. Volutharpa perryi (without black streak); E-H. Volutharpa perryi (with black streak)A, E. 外壳; B, F. 外壳和软体; C, G. 足; D, H. 生殖器远末端A, E. shell; B, F. shell with soft part; C, G. foot; D, H. distal end of penis
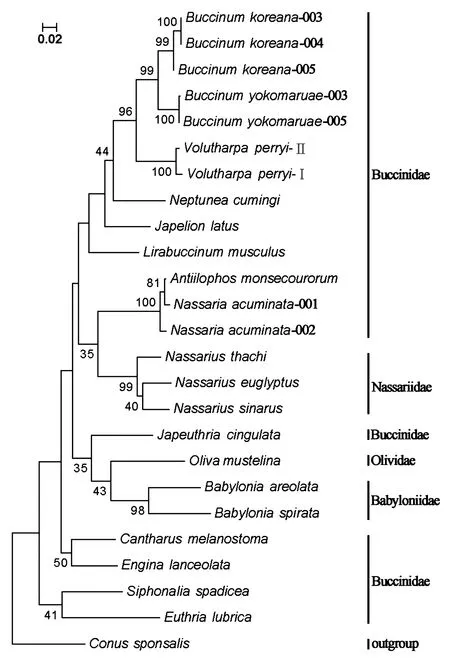
图2 基于CO Ⅰ基因序列片段的系统树(最大似然法)Fig.2 Phylogenetic tree based on CO Ⅰ gene sequences (maximum likelihood method)
[1] Taylor J D, Morris N J, Taylor C N. Food specialization and the evolution of predatory prosobranch gastropods[J]. Palaeontology, 1980, 23(2): 375-409.
[2] Bouchet P, Rocroi J P. Classification and nomenclator of gastropod families[J]. Malacologia, 2005, 47(1/2): 1-397.
[3] Ponder W F. The origin and evolution of the Neogastropoda[J]. Malacologia, 1974, 12(2): 295-338.
[4] Ponder W F, Warén A. Classification of the Caenogastropoda and Heterostropha-a list of the family-group names and higher taxa[M]//Ponder W F. Prosobranch Phylogeny. Malacological Review Supplement 4, 1988: 288-326.
[5] Harasewych M G. Studies on bathyal and abyssal Buccinidae (Gastropoda: Neogastropoda): 1.MetulafusiformisClench & Aguayo, 1941[J]. Nautilus, 1990, 104(4): 120-129.
[6] Vaught K C. A Classification of the Living Mollusca[M]. Florida: American Malacologists, 1989.
[7] Higo S I, Callomon P, Goto Y. Catalogue and Bibliography of the Marine Shell-bearing Mollusca of Japan[M]. Osaka: Elle Scientific Publications, 1999.
[8] Matthews-Cascon H, Alencar H A P, Rabay S G, et al. Sexual dimorphism in the radula of “Pisaniapusio” (Linnaeus, 1758) (Mollusca, Gastropoda, Buccinidae)[J]. Thalassas, 2005, 21(1): 29-33.
[9] Cernohorsky W O. Supplementary notes on the taxonomy of buccinid species of the subfamily Pisaniinae (Mollusca: Gastropoda)[J]. Records of the Auckland Institute and Museum, 1975, 12: 175-211.
[10] 张树乾, 张素萍. 中国近海蛾螺科(Gastropoda: Buccinidae)种类齿舌的扫描电镜观察[J]. 海洋与湖沼, 2016, 47(2): 414-424.
Zhang Shuqian, Zhang Suping. SEM study on radulae of Chinese marine Buccinids (Gastropoda: Buccinidae)[J]. Oceanologia et Limnologia Sinica, 2016, 47(2): 414-424.
[11] Kantor Y I. Comparative anatomy of the stomach of Buccinoidea (Neogastropoda)[J]. Journal of Molluscan Studies, 2003, 69(3): 203-220.
[12] Galino L A, Puillandre N, Utge J, et al. The phylogeny and systematics of the Nassariidae revisited (Gastropoda, Buccinoidea)[J]. Molecular Phylogenetics and Evolution, 2016, 99: 337-353.
[13] Harasewych M G, Kantor Y I. On the morphology and taxonomic position ofBabylonia(Neogastropoda: Babyloniidae)[J]. Bollettino Malacologico Supplemento, 2002, 38(S4): 19-36.
[14] Hayashi S. The molecular phylogeny of the Buccinidae (Caenogastropoda: Neogastropoda) as inferred from the complete mitochondrial 16S rRNA gene sequences of selected representatives[J]. Molluscan Research, 2005, 25(2): 85-98.
[15] Hou L, Dahms H U, Dong C Y, et al. Phylogenetic positions of some genera and species of the family Buccinidae (Gastropoda: Mollusca) from China based on ribosomal RNA and COI sequences[J]. Chinese Science Bulletin, 2013, 58(19): 2315-2322.
[16] Endo M, Ozawa T. Phylogenetic analysis ofBuccinumandNeptuneainferred from mtDNA sequences[C]//Abstract with Programs of the 150th Regular Meeting of the Palaeontological Society of Japan. 2001: 45.
[17] Iguchi A, Ito H, Ueno M, et al. Molecular phylogeny of the deep-seaBuccinumspecies (Gastropoda: Buccinidae) around Japan: Inter- and intraspecific relationships inferred from mitochondrial 16S rRNA sequences[J]. Molecular Phylogenetics and Evolution, 2007, 44(3): 1342-1345.
[18] Nakano T, Kurihara Y, Miyoshi H, et al. Molecular phylogeny ofNeptunea(Gastropoda: Buccinidae) inferred from mitochondrial DNA Sequences, with description of a new species[J]. Venus, 2010, 68(3/4): 121-137.
[19] Folmer O, Black M, Hoeh W, et al. DNA primers for amplification of mitochondrial cytochromecoxidase subunit I from diverse metazoan invertebrates[J]. Molecular Marine Biology and Biotechnology, 1994, 3(5): 294-299.
[20] Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences[J]. Journal of Molecular Evolution, 1980, 16(2): 111-120.
[21] Tamura K, Stecher G, Peterson D, et al. MEGA6: molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution, 2013, 30(12): 2725-2729.
[22] 张爱菊, 尤仲杰. 基于16S rDNA序列片段探讨织纹螺的系统发生关系[J]. 动物分类学报, 2008, 33(3): 549-552.
Zhang Aiju, You Zhongjie. Molecular phylogenetic relationships of some species of nassarius (gastropoda, prosobranchia, neogasrtopoda, nassariidae) based on 16srdna fragment sequence[J]. Acta Zootaxonomica Sinica, 2008, 33(3): 549-552.
[23] 陈志云, 张素萍. 基于16S rRNA基因序列对织纹螺属的分子系统学分析[J]. 动物分类学报, 2012, 37(3): 467-472.
Chen Zhiyun, Zhang Suping. Molecular phylogeny ofNassarius(gastropoda, nassariidae) based on 16s rRNA gene sequences[J]. Acta Zootaxonomica Sinica, 2012, 37(3): 467-472.
[24] Duda T F Jr, Bolin M B, Meyer C P, et al. Hidden diversity in a hyperdiverse gastropod genus: discovery of previously unidentified members of aConusspecies complex[J]. Molecular Phylogenetics and Evolution, 2008, 49(3): 867-876.
[25] Powell A W B. The recent and Tertiary species of the genusBuccinulumin New Zealand, with a review of related genera[J]. Transactions and Proceedings of the Royal Society of New Zealand, 1929, 60: 57-101.
[26] Smith E A. Notes onVolutharpaperryi[J]. Annals and Magazine of Natural History, 1887, 20(119): 347-348.
[27] Cernohorsky W O. Indo-Pacific Pisaniinae (Mollusca: Gastropoda) and related buccinid genera[J]. Records of the Auckland Institute and Museum, 1971, 8: 137-167.
[28] Cernohorsky W O. The family Buccinidae. Part 1: the generaNassaria,Trajana, andNeoteron[M]//Abbott R T. Monographs of Marine Mollusca. Florida: American Malacologists, Inc, 1981: 1-52.
[29] Woodring W P. Miocene mollusks from Bowden, Jamaica. Part Ⅱ. Gastropods and discussion of results. Contributions to the Geology and Paleontology of the West Indies[M]. Washington, DC: Carnegie Institution of Washington, 1928.
[30] Vokes E H. The genusTrajana(Mollusca: Gastropoda) in the New World[J]. Tulane Studies in Geology and Paleontology, 1969, 7(2): 75-83.
[31] Adams A. A monograph ofPhos, a new genus of gasteropodous Mollusca[J]. Proceedings of the Zoological Society of London, 1851, 18: 152-155.
[32] Vermeij G J. Taxonomy, distribution, and characters of Pre-Oligocene members of theCantharusgroup of Pisaniinae (Neogastropoda: Buccinoidea)[J]. Journal of Paleontology, 2001, 75(2): 295-309.
[33] Kuroda T, Habe T, Oyama K. The Sea Shells of Sagami Bay[M]. Tokyo: Maruzen, 1971.
Molecular phylogeny of Buccinidae (Gastropoda, Neogastropoda) from China seas based on CO Ⅰ gene sequences
Zhang Shuqian1, Zhang Suping1
(1.InstituteofOceanology,ChineseAcademyofSciences,Qingdao266071,China)
The partial mitochondrial cytochrome oxidase Ⅰ gene (CO Ⅰ) sequences of 18 Buccinoidea species were employed to evaluate the phylogenetic relationships of the family Buccinidae at the intra- and suprafamilial level. Within the family, the buccinids formed four clades: Clade Ⅰ includesJapelionlatus,Buccinumkoreana,Buccinumyokomaruae,Voluthapaperryi,Neptuneacumingi,LirabuccinummusculusandJapeuthriacingulata;Clade Ⅱ includesAntillophosmonsecourorumandNassariaacuminata; Clade Ⅲ includesCantharusmelanostomaandEnginalanceolata; Clade Ⅳ includesSiphonaliaspadiceaandEuthrialubrica. This result generally agrees with the previous subfamilial assignments based on morphological data. The monophyly of Buccinidae was interrupted by insertion of Nassariids, Olivids and Babyloniids. Babyloniidae, previously considered as a subfamily of Buccinidae, is now treated as an independent family. The present study confirmed this assignment. The generaNassariaandAntillophosclustered together with nassariids closer than with other buccinids, indicating that they have very close relationship with Nassariidae, this result is consistent with that of recently published molecular study.
Gastropoda; Buccinidae; phylogeny; cytochrome oxidase Ⅰ gene
10.3969/j.issn.0253-4193.2017.02.008
2016-07-14;
2016-11-11。
国家自然科学基金项目(41376167)。
张树乾(1984—),男,河北省石家庄市人,助理研究员, 主要从事海洋贝类分类与系统演化研究。E-mail: zsqtaxon@qdio.ac.cn
*通信作者:张素萍,研究员,主要从事海洋贝类分类与系统演化研究。E-mail:museum@qdio.ac.cn
S917.4
A
0253-4193(2017)02-0086-10
张树乾,张素萍. 基于线粒体CO Ⅰ基因序列对中国海蛾螺科的分子系统学分析[J].海洋学报,2017,39(2):86—95,
Zhang Shuqian, Zhang Suping. Molecular phylogeny of Buccinidae (Gastropoda, Neogastropoda) from China seas based on CO Ⅰ gene sequences[J]. Haiyang Xuebao,2017,39(2):86—95, doi:10.3969/j.issn.0253-4193.2017.02.008
