入侵藤本薇甘菊对酸性土壤的适应性
2016-12-19胡亮邓太阳张庆生李鸣光
胡亮,邓太阳,张庆生,李鸣光*
1. 中山大学地理科学与规划学院,广东 广州 510275;2. 中山大学有害生物控制与资源利用国家重点实验室,广东 广州 510275;3. 深圳市宝安区绿化委员会,广东 深圳 518101;4. 深圳市野生动植物保护管理处,广东 深圳 518048
入侵藤本薇甘菊对酸性土壤的适应性
胡亮1,2,邓太阳3,4,张庆生3,李鸣光2*
1. 中山大学地理科学与规划学院,广东 广州 510275;2. 中山大学有害生物控制与资源利用国家重点实验室,广东 广州 510275;3. 深圳市宝安区绿化委员会,广东 深圳 518101;4. 深圳市野生动植物保护管理处,广东 深圳 518048
有害藤本植物薇甘菊(Mikania micrantha)原产中南美洲,现已对我国华南地区的植被和生态造成了严重危害。了解薇甘菊对非生物环境因子的适应性是对其进行科学治理的基本前提之一。华南地区以酸性土壤为主,而铝毒害是酸性土壤环境中植物生长的重要限制因子。为揭示薇甘菊对土壤pH条件及铝毒的适应性,本研究对比了薇甘菊在不同土壤pH环境下的萌发和生长情况;比较分析了薇甘菊3个不同种群和其他5种植物种子在不同浓度铝离子胁迫(0、0.5、1和2 mmol∙L-1)下的种子萌发和幼苗生长;测定了深圳内伶仃岛薇甘菊及其他19种本地不同生活型植物茎干中3种微量元素(铝、锰和铁)的含量。结果表明,(1)薇甘菊种子在pH值为3.6~7.9的土壤环境中都能正常萌发,种子萌发率随土壤pH值的升高而逐渐降低;薇甘菊幼苗在pH值为4.3的条件下生长最佳,幼苗苗高、叶数、根数和根长都显著高于其他处理梯度。(2)当Al3+浓度不高于2 mmol∙L-1时,薇甘菊种子的萌发率无显著组间差异,与对照植物相比也没有显著差异;但薇甘菊不同种群之间的差异明显,尤其是在幼苗存活率上。(3)薇甘菊茎干中铝(655.3 mg∙kg-1)和铁(309.9 mg∙kg-1)的含量均为20种受试植物中的最高值,分别是其余19种对照植物平均值的11.5倍和3.3倍;锰含量(469.4 mg∙kg-1)仅次于鸭脚木,是其余19种植物平均值的2.7倍。以上结果表明薇甘菊对土壤酸碱度有较广的生态幅,对土壤铝离子毒害具有一定的耐受和富集能力,且铝离子毒害未影响薇甘菊对其他离子的吸收,因此对酸性土壤有较强的适应性。薇甘菊对酸性土壤的适应性有利于其成功入侵华南地区,而该区域土壤酸化的加剧可能进一步加重薇甘菊的潜在危害。
薇甘菊;铝毒;土壤pH;种子萌发;幼苗生长;微量元素
生物入侵是指一种生物扩散到其原产地以外的地区并成功定居、繁衍、扩散和造成危害,是除生境破坏之外造成生物多样性丧失的第二主导因素(Wilcove et al.,1998)。入侵生物往往对被入侵地区的生物多样性、生态系统和经济造成了巨大的危害和长期的负面影响(Holm et al.,1977;徐汝梅等,2004)。薇甘菊(Mikania micrantha H.B.K.)是受人类活动显著影响而造成危害的入侵植物之一。该种原产中、南美洲,现已在亚太热带、亚热带地区广泛分布并造成严重的生态危害(Holm et al.,1977;徐汝梅等,2004)。薇甘菊对生态环境的适应性强(徐高峰等,2014),有性生殖和无性繁殖能力都很强,植株生长迅速并具有缠绕攀援的习性。通过在其他植物体的茎干、枝叶上缠绕蔓延,薇甘菊能迅速扩张呈大面积覆盖,从而抑制其他植物的生长,降低群落物种多样性和生物量并阻碍群落恢复(Waterhouse,1994;周先叶等,2005;毛子龙等,2011)。
从宏观尺度上对入侵生物进行预测和防控,必须立足于对其生理和生态适应性的研究。因此,了解入侵植物对光、温、水、土等非生物环境因子的适应性是对其进行科学预测和防控的基本前提(Zalba et al.,2000;胡亮,2009)。为此,众多学者已先后从光照(胡玉佳等,1994;张炜银等,2002;廖飞勇等,2003;张玲玲等,2009)、温度(李鸣光等,2002;Yang et al.,2005)和水分(杨期和等,2003;徐高峰等,2013;胡亮等,2014)等非生物环境因子对薇甘菊生长的影响进行深入研究。薇甘菊与土壤环境的生态学研究目前主要集中于薇甘菊对土壤理化性质及微生物环境的改变(李伟华等,2008;吴双桃,2011;杨琼等,2015)等方面,但尚未有研究论及薇甘菊对土壤条件的适应性。野外调查表明薇甘菊在土壤pH值为4.15~8.25的生境中均有分布(曹洪麟等,2003)。那么,在不同土壤pH条件下薇甘菊的种子萌发和植株生长是否存在差异?我国华南地区森林土壤主要为酸性土和强酸性土(刘飞鹏,2007),土壤酸化的主要后果之一是使土壤中的含铝矿物释放出大量可溶性的铝离子,进而影响其他阳离子的吸收,并抑制植物细胞分裂和根的生长(Minocha et al.,2005)。因此铝毒害是酸性土壤环境中植物生长的重要限制因子(刘强等,2004;吴道铭等,2013)。与其他植物相比,薇甘菊对铝毒害的耐受能力如何?薇甘菊在酸性土壤中采取的是铝富集还是铝排斥策略?其策略是否会影响薇甘菊对其他离子的吸收?上述问题的解答可揭示薇甘菊对土壤环境的适应性,有利于对薇甘菊的分布和危害进行科学预测和评估。
本研究设置一系列实验以揭示薇甘菊对酸性土壤及铝毒的适应性。通过对比薇甘菊在不同pH土壤环境下的种子萌发和幼苗生长状况,揭示其对土壤环境的偏好;通过对比薇甘菊和其他常见本地、外来草本植物在不同程度铝离子胁迫下的种子萌发和幼苗生长情况,揭示薇甘菊对铝毒的耐受性;通过测定薇甘菊及其他19种本地不同生活型植物茎干中微量元素铝、锰和铁的含量,揭示薇甘菊对铝毒的耐受模式及其对其他阳离子吸收的影响。
1 材料与方法
本研究中,铝离子效应实验所采用的薇甘菊种子包括 2007—2008年间分别采自深圳内伶仃岛(MMN)、珠海淇澳岛(MMQ)和深圳百果园(MMB)的3个种群批次,并选取珠江三角洲5种常见草本植物为对照。分别是菊科草本植物假臭草 Praxelis clematidea(PC)、藿香蓟Ageratum conyzoides(AC)、鬼针草Bidens pilosa(BP)、翅果菊Lactuca indica(LI)及旋花科草质藤本三裂叶薯 Ipomoea triloba(IT)。其中假臭草、藿香菊、鬼针草和三裂叶薯为外来植物,翅果菊为珠三角地区的本地种。所有对照植物的种子均采集于2008年6—11月。实验时间为2009年2—3月,选取饱满、无破损的种子进行实验。
1.1 土壤酸碱度对薇甘菊种子萌发和幼苗生长的影响
采集广州市龙眼洞森林公园次生林下土壤(pH 3.3~3.8),去除杂物,搅拌均匀后,称取100.0 g土壤30份,设置5个梯度组,每组6份。其中一组不加添加物(CK),余下 4组在每份土壤中分别加入0.1、0.5、0.75和1 g分析纯氧化钙(CaO,灼烧后含量不低于98.0%),混合均匀后,将每份土壤置于小塑料盆中(口径7.5 cm,底径5 cm,深6.5 cm,底部铺1层定性滤纸),加水浸润并静置24 h后,每个梯度取1份测量其pH值。余下5份在每份中放置MMN种子20粒,每个梯度组总计100粒种子。每24小时补充水分,每2天记录1次萌芽情况。第30天统计萌发率,并在每个梯度组中各选2份土壤测量pH值和所有幼苗苗高、叶数、根数和根长;其余3份于第45天时测量相同指标。以幼苗苗高、叶数、根数和根系长作为组间比较指标。
1.2 土壤铝离子效应对薇甘菊种子萌发和幼苗生长的影响
为揭示薇甘菊对铝毒的耐受能力及与其他植物之间的差异,选取饱满、无破损的MMN、MMB、MMQ、PC、AC、BP、LI和IT种子各1批。为保持实验过程中稳定的胁迫条件,在放有两层定性滤纸的9 cm培养皿中分别加入5.0 g蒸馏水(CK)或 Al3+浓度分别为 0.5、1.0、2.0 mmol∙L-1的Al2(SO4)3溶液(因BP和IT生长迅速,采用12 cm培养皿培养并加入10.0 g溶液),每皿放置30粒受试种子,每个浓度梯度设置3个重复。称取各皿的总重作为补充损失水分的依据。将培养皿放置于RXZ-300B型人工气候箱中(光处理12 h,30 ℃,光照2500~3500 lux;暗处理12 h,28 ℃)。以胚根突破种皮作为萌发标准,每24小时记录1次萌发数并用称重法补充损失水分。实验第14天测量叶片已突破种皮的全部幼苗之根长。以相对萌发率、幼苗死亡率和相对根长作为种间比较指标。
1.3 薇甘菊和19种常见植物茎中的微量元素测定
2007年6月于深圳内伶仃岛采集薇甘菊的茎以测定其铝、锰和铁含量,并以 19种当地常见植物作为对照:其中藤本植物11种(两面针Zanthoxylum nitidum、龙须藤 Bauhinia championii、首冠藤Bauhinia corymbosa、山牵牛Thunbergia grandiflora、飞龙掌血 Toddalia asiatica、小果葡萄 Vitis balansana、秤钩风Diploclisia affinis、耳叶马兜铃Aristolochia tagala、玉叶金花Mussaenda pubescens、水忍冬 Lonicera confusa和锡叶藤 Tetracera sarmentosa);乔灌木8种(血桐Macaranga tanarius var. tomentosa、亮叶猴耳环Archidendron lucidum、破布叶 Microcos paniculata、鹅掌柴 Schefflera heptaphylla、白楸 Mallotus paniculatus、簕欓花椒Zanthoxylum avicennae、紫玉盘Uvaria macrophylla和鹰爪花Artabotrys hexapetalus)。每种植物的材料都采自同一种群中3个不同的植株。将采回的枝茎剪碎、烘干、粉碎并全部过 100目筛。用干灰化-稀盐酸溶解-ICP法测定铝元素含量;用干灰化-1∶1盐酸溶解-原子吸收分光光度法测定锰元素含量;用干灰化-1∶1盐酸溶解-原子吸收分光光度法测定铁元素含量。
实验结果在软件SPSS 15.0(SPSS Inc. 2006)中进行分析,并采用ANOVA检验各处理组间差异。
2 结果
2.1 土壤酸碱度对薇甘菊种子萌发和幼苗生长的影响
薇甘菊种子在pH值为3.6~7.9的土壤环境中都能正常萌发。种子萌发率在极强酸性土(pH=3.6)环境中最高,达到最大萌发率的时间也最短。随土壤pH值的升高,种子萌发率逐渐降低(表1)。与萌发率最高的组(pH=3.6)相比,在pH值为6.5、7.6和7.9的土壤环境中薇甘菊种子的萌发率分别降低了约11%、20%和26%。
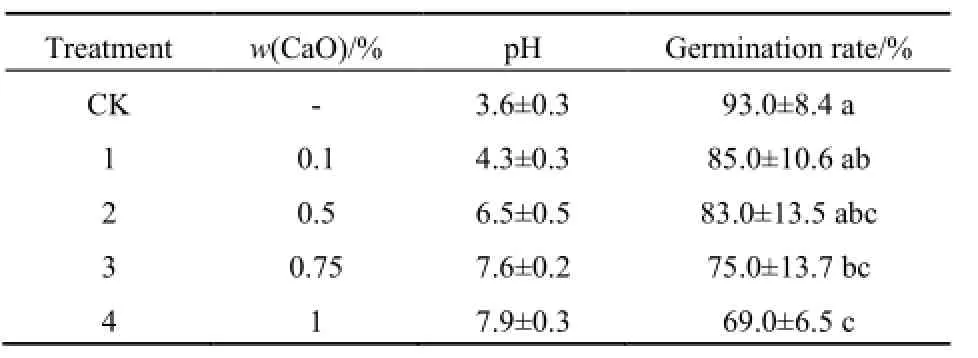
表1 薇甘菊种子在不同pH值土壤中的萌发率Table 1 Germination rate of Mikania micrantha seeds in soil with different pH
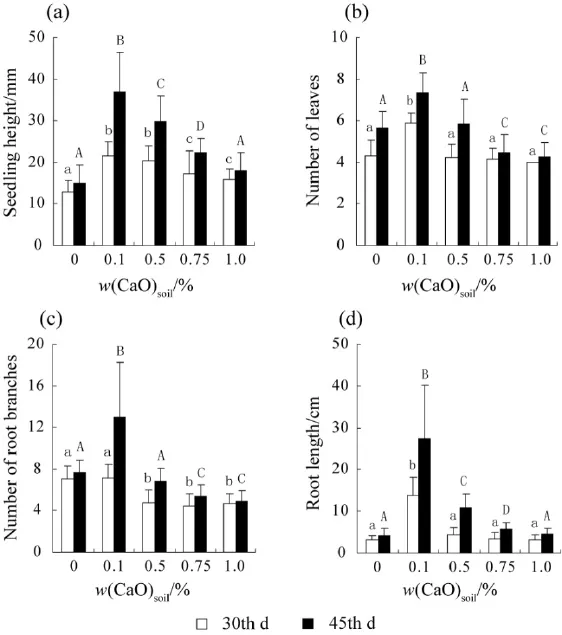
图1 薇甘菊在加入不同量氧化钙的土壤环境中生长30 d和45 d后的生长指标Fig. 1 Growth indices of Mikania micrantha grown in soils with different amounts of CaO for 30 days and 45 days
薇甘菊的幼苗生长也显示其对酸性土壤的适应性强。土壤pH值为4.3时薇甘菊幼苗苗高、叶数、根数和根长均显著高于对照组和其他处理梯度(图1);当土壤pH值继续升高时,薇甘菊幼苗的苗高、叶数、根长和根数均逐渐递减,其中根长缩短的趋势最为明显。且随着生长时间的延长,这种缩短趋势更为明显。第 45天,与根长最长的组(pH=4.3)相比,在pH值为6.5、7.6和7.9的土壤环境中薇甘菊根长分别缩短约48%、59%和63%。在pH值为3.6的极强酸性土壤环境中,薇甘菊的苗高和根长受抑制程度与在pH值为7.9的微碱性土壤环境中相当(图1a、图1d),但叶数和根数的受抑制程度相对较低(图1b、图1c)。
2.2 土壤铝离子效应对薇甘菊种子萌发和幼苗生长的影响
实验最高Al3+浓度为2 mmol∙L-1,该浓度下因Al3+导致的水分胁迫低于质量分数仅为 0.035%的NaCl盐度,可忽略水势胁迫的影响。经测定,Al3+浓度为0.5、1.0和2.0 mmol∙L-1的溶液pH值依次为4.0、3.5和3.0。
2.2.1 种子萌发
总体上,未见铝毒胁迫明显抑制薇甘菊和其他受试植物种子的萌发。各Al3+浓度胁迫条件下,薇甘菊3个种群和其余5种对照植物的种子萌发率均与各自对照组无显著差异(表2)。所有受试种群的种子初始萌发时间均未出现明显滞后,种子累计萌发情况与对照组均较为一致。
2.2.2 幼苗存活率
薇甘菊3个受试种群的幼苗对Al3+毒害的耐受能力均高于假臭草和藿香蓟,低于鬼针草、翅果菊和三裂叶薯。铝毒显著影响了MMQ、MMB、假臭草和藿香蓟已萌发幼苗的存活率,而对 MMN、鬼针草、翅果菊和三裂叶薯则没有显著影响。随Al3+浓度增加,薇甘菊、假臭草和藿香蓟已萌发幼苗死亡率显著上升。若以Al3+浓度为2 mmol∙L-1胁迫下的死亡率情况作为耐受性标准,则8个受试种群幼苗对铝毒的耐受性高低顺序为:MMN=LI=BP=IT>MMQ>MMB=AC>PC。
薇甘菊不同种群对土壤铝毒的耐受性具有较大差异,耐受能力从高到低依次为内伶仃岛种群、淇澳岛种群和百果园种群。本研究考虑到薇甘菊的藤本习性,既加入了外来植物,又加入了藤本植物作为对比;但结果表明植物对铝毒的耐受能力偏向于种间差异,而外来种和本地种在对铝毒耐受性上不存在明显差异,直立种和缠绕藤本之间亦是如此。相比之下,薇甘菊3个种群之间的差异反而更为明显,尤其是在幼苗存活率上(表3)。
2.2.3 幼苗生长
总体上,薇甘菊因铝离子效应导致的根长受抑制程度高于假臭草和鬼针草,低于藿香蓟和翅果菊。铝离子浓度对7个受试种群植物的主根根长都有显著影响(因三裂叶薯幼苗生长迅速,实验末期其生长明显受培养皿空间抑制,故此处不加以讨论)。随 Al3+浓度增加,主根根长受抑制程度也单调增强。藿香蓟和翅果菊幼苗主根根长受抑制程度相对较高;鬼针草受抑制程度最低。MMN根长受抑制程度较高,MMB和MMQ相对较低且两种群间无显著差异(表4)。以50%相对根长为耐受性标准,7个受试种群对铝毒的耐受性高低顺序为:PC>BP=MMB=MMQ>MMN=AC=LI。
2.3 薇甘菊和19种常见植物茎中的微量元素
薇甘菊茎干中微量元素铝和铁的含量显著高于其他植物,锰的含量也较高。薇甘菊茎中铝的质量分数高达655.3 mg∙kg-1,是其余19种植物平均值的11.5倍,比次高的玉叶金花(193.6 mg∙kg-1)高出2.4倍。薇甘菊茎中铁质量分数(309.9 mg∙kg-1)在20种受试植物中也是最高;是其余19种本地植物平均值的 3.3倍。薇甘菊茎中锰质量分数(469.4 mg∙kg-1)为藤本植物中最高,仅次于鸭脚木(597.6 mg∙kg-1),是 19种本地植物平均值的2.7倍。除薇甘菊外,其余由藤本组成的藤本种组与乔灌种组之间不存在因生活型而体现出的明显的组间差异(图2)。
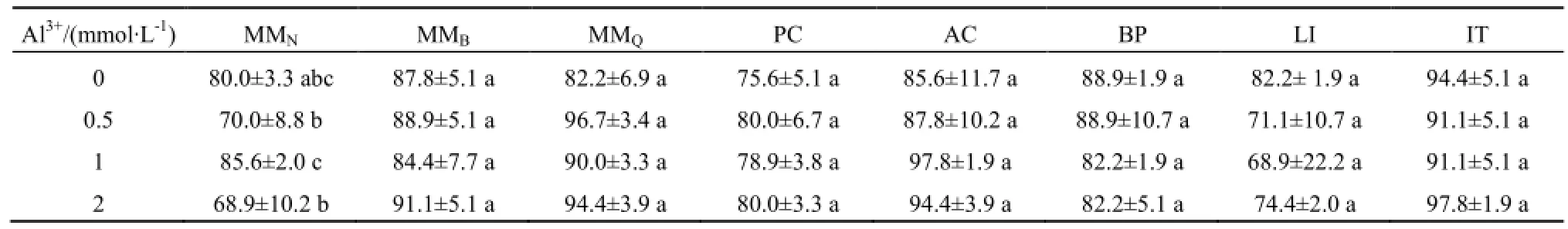
表2 薇甘菊等6种植物种子在不同浓度Al3+胁迫下的萌发率Table 2 Seed germination rates of six species under different concentration of Al3+stresses %

表3 薇甘菊等6种植物在不同浓度Al3+胁迫下的幼苗死亡率Table 3 Seedling mortality of six species grown under different concentration of Al3+stresses %

表4 薇甘菊等5种植物在不同浓度Al3+胁迫下生长14 d的根长Table 4 Average root length of five species grown under different concentration of Al3+stresses for 14 days mm
3 讨论
薇甘菊为酸性土植物。当其他条件适宜时,在pH为 3.6~7.9的土壤环境中薇甘菊都能萌发和生长;且在pH值为4.3的极强酸性土壤中幼苗生长最佳。广东省是我国受薇甘菊危害最为严重的省份,其森林土壤主要为酸性土和强酸性土,其中强酸性土壤占30%(刘飞鹏,2007;王登峰等,2004);且酸化程度和铝毒害日趋严重(吴道铭等,2013)。依据土壤pH梯度实验的结果,推测薇甘菊在强酸性土壤上的生长情况极可能比在pH值为6.5及以上的中性或微碱性土壤环境中生长要好。因此,土壤pH不会成为薇甘菊在广东地区分布和蔓延的限制因素。吴双桃(2011)比较了野外薇甘菊入侵地及其相邻无薇甘菊分布对照样地土壤,发现薇甘菊入侵地土壤pH值显著高于后者;由于被入侵地在薇甘菊入侵前的土壤理化性质不详,无法判断这是薇甘菊入侵带来的影响还是原土壤环境的特殊性。而杨琼等(2015)的盆栽实验结果显示薇甘菊生长并未导致土壤pH值发生显著变化。因此,薇甘菊在酸性土壤环境生长得更好可能是薇甘菊本身就适应酸性土壤,而不是改变了其pH值。在薇甘菊的潜在危害区域中,广西的石灰岩面积较大。石灰岩地区生态环境脆弱,土层浅薄,干旱缺水,植被生长缓慢;一旦遭受破坏会迅速恶化,难于恢复甚至不可逆转(陈作雄,1995)。秦新生等(2008)的研究表明,藿香蓟和假臭草在海南岛石灰岩中分布较广,因此石灰岩地区仍可能是薇甘菊的潜在分布范围。但与酸性土壤条件相比,石灰岩地区薇甘菊的生长将会受显著抑制,至于其受抑制程度与其他石灰岩喜钙耐旱植物相比如何,其无性繁殖和蔓延速度是否会威胁石灰岩地区的植物生长和群落演替,这些问题有待进一步研究。
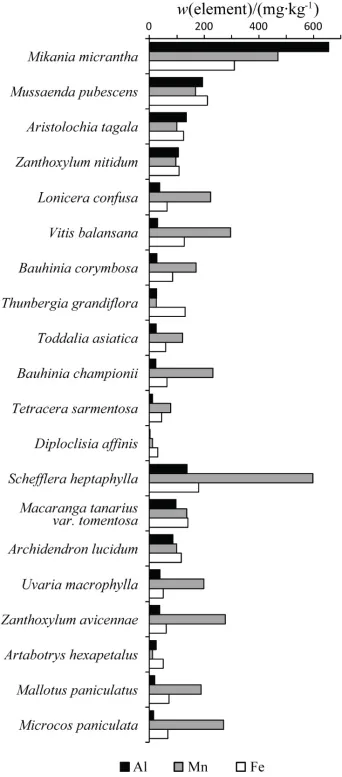
图2 深圳内伶仃岛20种植物茎干中铝、锰和铁元素含量Fig. 2 Contents of Al, Mn and Fe elements in the stems of 20 species in Neilingding island, Shenzhen
薇甘菊不同种群对土壤铝毒的耐受性具有较大差异。尽管薇甘菊的种子萌发率在3个受试种群间没有显著差异,但幼苗期的存活率表现为内伶仃岛种群显著较高于其他两个种群,淇澳岛种群次之,百果园种群最低。铝毒实验中所设Al3+梯度没有调节 pH,但 pH梯度实验结果显示在铝毒实验pH的波动范围内薇甘菊都能生长,因此pH的差异不致死薇甘菊,幼苗的死亡主要是Al3+毒害。种群之间耐受能力的差异可能源于薇甘菊对异质环境的适应性,但需进一步实验验证。
薇甘菊茎干中铝、铁、锰的含量高表明薇甘菊对酸性土壤的适应性高。耐铝毒是植物适应酸性土壤所必须的能力,较高的耐铝毒能力是植物适应酸性土壤环境的关键因素(Marschner,1995)。在内伶仃岛,薇甘菊茎干中铝含量比相同生境中其他藤本植物和木本植物显著要高,也比其他地带性植物如假苹婆 Sterculia lanceolata、朱砂根 Ardisia crenata、台湾相思 Acacia confusa、红楠 Machilus thunbergii等要高,约为同地带铝超富集植物茶树Camellia sinensis茎干中所测浓度的69%~72%(谢正苗等,2002),表明薇甘菊具有一定的铝富集作用。薇甘菊茎干中微量元素铁和锰的含量较高,表明薇甘菊对其他阳离子的吸收没有受到铝离子的明显抑制;而生长迅速、对离子需求量大可能是其含量远超其他藤本植物的主要原因。推测薇甘菊在其根系和叶片等部位同样具有铝富集能力,但实际情况以及铝在其内部的转运、累积机理还有待进一步研究。
4 结论
薇甘菊对土壤酸碱度有较广的生态幅,对土壤铝毒具有一定的富集和耐受能力,且铝离子毒害未影响薇甘菊对其他离子的吸收,因此薇甘菊对酸性土壤有较强的适应性。薇甘菊对非生物环境的广适性可能是其入侵成功的原因之一,华南地区的水热条件适合薇甘菊的生长,薇甘菊对酸性土壤的偏好以及南方土壤酸化问题的不断加剧可能进一步加深其潜在的危害程度和扩大分布范围。
HOLM L G, PLUCKNETT D L, PANCHO J V, et al. 1977. The world’s worst weeds: distribution and biology [M]. Hawaii: The University Press of Hawaii: 320-327.
MARSCHNER H. 1995. Mineral Nutrition of Higher Plants (Second edition) [M]. London: Academic Press.
MINOCHA R, MINOCHA S C. 2005. Effects of soil pH and Aluminum on plant respiration[C]//Lambers H, Ribas-Carbo M. Plant Respiration. Netherlands: Springer: 159-176.
WATERHOUSE D F. 1994. Biological Control of Weeds: Southeast Asian Prospects [M]. Canberra: Australian Centre for International Agricultural Research.
WILCOVE D S, ROTHSTEIN D, DUBOW J, et al. 1998. Quantifying threats to imperiled species in the United States [J]. BioScience, 48(8): 607-615.
YANG Q H, WANHUI Y E, DENG X, et al. 2005. Seed germination eco-physiology of Mikania micrantha H.B.K [J]. Botanical Bulletin of Academia Sinica, 46(4): 293-299.
ZALBA S M, SONAGLIONI M I, COMPAGNONI C A, et al. 2000. Using a habitat model to assess the risk of invasion by an exotic plant [J]. Biological Conservation, 93(2): 203-208.
曹洪麟, 冯惠玲, 邓雄, 等. 2003. 薇甘菊在广东的分布与危害[C]//徐汝梅, 叶万辉. 生物入侵: 理论与实践. 北京: 科学出版社: 236-248.
胡亮. 2009. 薇甘菊Mikania micrantha H.B.K.对非生物环境改变的响应及其分布预测[D]. 广州: 中山大学.
胡玉佳, 毕培曦. 1994. 微甘菊生活史及其对除莠剂的反映研究[J]. 中山大学学报: 自然科学版, 33(4): 88-95.
李鸣光, 张炜银, 王伯荪, 等. 2002. 薇甘菊种子萌发特性的初步研究[J].中山大学学报: 自然科学版, 41(6): 57-59.
廖飞勇,谢瑛,何平, 等. 2003.不同光强对薇甘菊生长及光系统的影响[J]. 生命科学研究, 7(4): 355-359.
李伟华, 韩瑞宏, 高桂娟. 2008. 薇甘菊入侵对土壤微生物生物量和土壤呼吸的影响[J]. 华南师范大学学报: 自然科学版, (3): 95-102.
刘飞鹏. 2007. 广东省森林土壤酸化现状及调控措施[J]. 林业调查规划, 32(4): 69-71.
毛子龙, 赖梅东, 赵振业, 等. 2011. 薇甘菊入侵对深圳湾红树林生态系统碳储量的影响[J]. 生态环境学报, 20(12): 1813-1818.
秦新生, 张荣京, 陈红锋, 等. 2008. 海南岛石灰岩地区的外来植物[J].生态学杂志, 27(11): 1861-1868.
王登峰, 薛春泉, 刘志武. 2004. 广东省森林生态状况监测报告[M]. 北京: 中国林业出版社.
吴道铭, 傅友强, 于智卫, 等. 2013. 我国南方红壤酸化和铝毒现状及防治[J]. 土壤, 45(4): 577-584.
吴双桃. 2011. 薇甘菊对入侵地植物群落及土壤理化性质的影响[J]. 湖北农业科学, 50(18): 3711-3713.
谢正苗, 黄铭洪, 叶志鸿. 2002. 铝超积累植物和铝排斥植物吸收和累积铝的机理[J]. 生态学报, 22(10): 1653-1659.
徐高峰, 申时才, 张付斗, 等. 2013. 土壤水分对薇甘菊不同繁殖体单位存活能力和植株表型可塑性影响[J]. 中国农业科学, 46(15): 3134-3141.
徐高峰, 申时才, 张付斗. 2014. 异质环境下入侵植物薇甘菊的适应性
与繁殖特性[J]. 生态环境学报, 23(8): 1258-1264.
徐汝梅, 叶万辉. 2004. 生物入侵: 理论与实践[M]. 北京: 科学出版社.杨期和, 冯惠玲, 叶万辉, 等. 2003. 环境因素对薇甘菊开花结实影响初探[J]. 热带亚热带植物学报, 11(2): 123-126.
杨琼, 梁羽, 杨剑, 等. 2015. 入侵植物薇甘菊的根际土壤微生物特征[J].生态科学, 34(2): 148-155.
张玲玲, 孙芳芳, 温达志. 2009. 薇甘菊和飞机草抗氧化物和膜脂过氧化物对光照和土壤水分的响应[J]. 植物生态学报, 33(5): 974-983.
张炜银, 王伯荪, 李鸣光, 等. 2002. 不同光照强度对薇甘菊幼苗生长和形态的影响[J]. 中山大学学报论丛, 22(1): 222-226.
周先叶, 王伯荪, 李鸣光, 等. 2005. 薇甘菊对内伶仃岛植被危害的相关分析[J]. 应用生态学报, 16(2): 350-354.
刘强, 郑绍建, 林咸永. 2004. 植物适应铝毒胁迫的生理及分子生物学机理[J]. 应用生态学报, 15(9): 1641-1649.
陈作雄. 1995. 广西石灰岩山区生态系统受害原因探讨[J]. 广西师院学报: 自然科学版, (1): 1-4.
胡亮, 李鸣光, 韦萍萍. 2014. 入侵藤本薇甘菊的耐盐能力[J]. 生态环境学报, 23(1): 7-15.
The Adaptation of Mikania micrantha to Acid Soil
HU Liang1,2, DENG Taiyang3,4, ZHANG Qingsheng4, LI Mingguang2*
1. Geography and Planning School, Sun Yat-Sen University, Guangzhou 510275, China; 2. State Key Laboratory of Biocontrol, Sun Yat-Sen University, Guangzhou 510275, China; 3. Greening Committee of Bao'an District, Shenzhen 518101, China; 4. Wildlife Management Department of Shenzhen, Shenzhen, 518048, China;
Mikania micrantha is one of the world’s worst invasive plants. It is now widespread in the tropics and south subtropics of China. Here we try to reveal the adaptation of M. micrantha to acid soil and aluminum toxicity by a set of experiments. Seed germination rate and seedling growth of M. micrantha in soil with different pH values were tested; seed germination rates and seedling mortalities of M. micrantha and other 5 weeds under different Al3+stresses (0, 0.5, 1 and 2 mmol∙L-1) were compared; and the contents of three microelements (Al, Mn and Fe) in the stems of M. micrantha and other 19 species were compared. The results showed that: (1) M. micrantha seeds could germinate in soil with pH values of 3.6~7.9 and its seedlings grew best in soil with pH value of 4.3. Its seed germination rate declined with increasing soil pH value. (2) The germination rate of M. micrantha seeds was not significantly different among groups when the concentration of Al3+was not higher than 2 mmol∙L-1. Seedling mortality indicated that Al-tolerance of M. micrantha was not significantly higher than the other tested species. However, the diffenence among M. micrantha populations is obvious, especially in seedling survival rate. (3) The content of Al in the stem of M. micrantha was the highest (655.3 mg∙kg-1), 11.5 times of the mean value of the other 19 tested plants. The content of Fe was also the highest (309.9 mg∙kg-1), 3.3 times of the mean value of the other 19 tested palnts. The content of Mn (469.4 mg∙kg-1) was 2.7 times of the mean value of the other 19 tested plants. We concluded that M. micrantha has a wide ecological amplitude of soil pH. It is an Al-tolerant plant. The adaptability of M. micrantha to acidic soil was favorable to its successful invasion in southern China. The exacerbated soil acidification in southern China may increase the risk of this species.
Mikania micrnatha; aluminum toxicity; soil pH; seedling growth; germination; microelement
10.16258/j.cnki.1674-5906.2016.09.003
Q948.113
A
1674-5906(2016)09-1439-07
胡亮, 邓太阳, 张庆生, 李鸣光. 2016. 入侵藤本薇甘菊对酸性土壤的适应性[J]. 生态环境学报, 25(9): 1439-1445.
HU Liang, DENG Taiyang, ZHANG Qingsheng, LI Mingguang. 2016. The Adaptation of Mikania micrantha to acid soil [J]. Ecology and Environmental Sciences, 25(9): 1439-1445.
国家自然科学基金项目(30370243);宝安生态防护林防治薇甘菊试验的效益监测项目
胡亮(1982年生),男,副教授,博士,研究方向为植物生态学和植物地理学。E-mail: huliang_hy@163.com *通信作者。E-mail: lsslmg@mail.sysu.edu.cn
2016-07-20
