全基因组预测枯草芽孢杆菌BEST195的分泌蛋白
2016-12-15任文来韩长志夏秀芹
任文来 韩长志 刘 通 夏秀芹
(1. 廊坊市文安县农业局,河北 廊坊 065800;2. 西南林业大学云南省森林灾害预警与控制重点实验室,云南 昆明 650224)
全基因组预测枯草芽孢杆菌BEST195的分泌蛋白
任文来1韩长志2刘 通1夏秀芹1
(1. 廊坊市文安县农业局,河北 廊坊 065800;2. 西南林业大学云南省森林灾害预警与控制重点实验室,云南 昆明 650224)
枯草芽孢杆菌BEST195作为纳豆菌的一种,是纳豆生产的重要菌株之一。利用SignalP、ProtComp、TMHMM、Phobius、LipoP、TatP等预测程序对该菌中4 456条蛋白质序列进行分泌蛋白找寻,同时,对上述分泌蛋白的氨基酸分布、信号肽长度大小、信号肽切割位点等性质进行分析。结果表明:枯草芽孢杆菌含有分泌蛋白102个,其氨基酸长度、信号肽长度与植物病原菌不同;信号肽切割位点属于A-X-A类型,与其他已经报道的植物病原真菌、细菌以及卵菌中分泌蛋白信号肽切割位点一致。通过上述生物信息学分析方法有效地实现了枯草芽孢杆菌BEST195分泌蛋白的预测,分泌蛋白的信号肽切割位点具有物种保守型特点。
枯草芽孢杆菌;分泌蛋白;信号肽;预测
枯草芽孢杆菌BEST195 (Bacillussubtilissubsp.nattoBEST195) 为纳豆菌的一种,是日本三菱化学生命研究所从纳豆中分离到的重要工业菌株[1]。纳豆菌属于枯草芽孢杆菌属,与该属中其他枯草芽孢杆菌不同,其具有将大豆发酵为纳豆的作用,可以产生较多的多聚γ-谷氨酸(γPGA),这是形成粘性聚合物的主要组成成分[2]。国内外学者对纳豆菌的研究主要集中在代谢产物纳豆激酶[3]、抗菌作用[4]、微生态制剂[5]等方面。
目前,学术界关于稻瘟菌[6]、大丽轮枝菌[7]、致病疫霉[8]等植物病原菌的分泌蛋白已经明确,并逐渐成为植物病理学研究的热点之一。然而,国内对于细菌分泌蛋白的预测研究尚不够深入,涉及诸多革兰氏阴性菌:香蕉细菌性软腐病菌[9]、茄青枯病菌[10]以及丁香假单胞杆菌番茄致病变种[11]等。同时,随着枯草芽孢杆菌BEST195全基因组序列的公布[12-13],对其开展了全基因组与其他纳豆菌的比较基因组学分析,明确纳豆菌中鞭毛蛋白FliF与yPGA的产生有直接关系,但yPGA的产生与否并不能作为枯草芽孢杆菌是否具有发酵功能的预测指标[2]。作为影响枯草芽孢杆菌yPGA产量的重要功能蛋白,纳豆菌中分泌蛋白具有十分重要的作用,然而,尚未见到对重要工业生产菌株——枯草芽孢杆菌BEST195分泌蛋白的预测报道,因此,开展该菌分泌蛋白的预测具有重要的理论意义。
基于细菌分泌蛋白所具有的主要特征,本研究利用SignalP、ProtComp、TMHMM、Phobius、LipoP、TatP等在线生物信息学分析程序对枯草芽孢杆菌BEST195进行分泌蛋白预测,以期为深入解析分泌蛋白在发挥发酵作用的功能解析提供重要的理论参考。
1 材料与方法
1.1 枯草芽孢杆菌BEST-195数据来源
B.subtilissubsp.nattoBEST195序列均来源于美国国立卫生院 (http://www.ncbi.nlm.nih.gov/genome)[13],其BioProject ID分别为PRJNA183001。
1.2 分泌蛋白确定方法
1.2.1 N-端信号肽预测
利用SignalP v4.1在线分析实现[14]。
1.2.2 亚细胞定位预测
对含有N-端含有信号肽的蛋白质序列,利用ProtCompB亚细胞定位预测实现 (http://linux1.softberry.com/berry.phtml?topic=pcompb&group=programs&subgroup=proloc)。
1.2.3 跨膜结构域预测
利用TMHMM v2.0 (http://www.cbs.dtu.dk/services/TMHMM/) 和Phobius[15]。
1.2.4 信号肽酶识别位点及脂蛋白预测
利用LipoP v1.0[16]和TatP v1.0[17]分别对蛋白进行信号肽酶识别位点、脂蛋白的预测。
2 结果与分析
2.1 枯草芽孢杆菌BEST195中典型分泌蛋白的数量特征
信号肽预测结果表明,仅有205个序列在N端含有典型的信号肽序列。进一步开展细胞定位情况分析发现,87个蛋白质分泌至胞外,30个蛋白尚未预测出定位情况,88个蛋白分泌到胞内,其中,转运至细胞质膜、细胞质的蛋白数量分别为77、11个。通过对103个蛋白 (包括87个转运到胞外的蛋白和30个尚未预测出定位情况的蛋白) 进行跨膜结构域分析,结果显示,56个蛋白质不含有跨膜结构域,另外56个蛋白含有1个跨膜结构域,还有2个蛋白含有2个跨膜结构域。由于TMHMM v2.0程序并不能完全对信号肽序列和所属跨膜域区序列进行区分,故选择112个蛋白质序列 (包括不含有任何跨膜结构域的蛋白以及含有1个跨膜结构域的蛋白) 进行后续分析。通过Phobius预测,结果显示gi 674710112、gi 674709268、gi 674708909共3个蛋白定位在细胞质中,获得109个分泌蛋白。通过对上述蛋白质序列进行LipoP分析,明确gi 674709563、gi 674709457为脂蛋白,通过排除及TatP分析,明确gi 674709457、674709254、428281022、gi 674709219、gi 428279469、gi 428280358共6个蛋白具有典型的tat信号肽,并获得102个分泌蛋白;因此,通过上述生物信息学在线程序对B.subtilissubsp.nattoBEST195中4456条蛋白质序列进行预测分析,最终获得了102个具有典型特征的分泌蛋白。
对上述分泌蛋白氨基酸大小进行分析,结果显示,这些分泌蛋白集中于101~500 aa之间,所占比例为84.31%,尤以201~300 aa分泌蛋白最多,所占比例为32.35% (图1)。
2.2 枯草芽孢杆菌BEST195典型分泌蛋白的信号肽特征
对上述分泌蛋白所含的信号肽长度进行分析,发现分泌蛋白中信号肽长度以27个aa的蛋白居多,所占比例为12.75%;分泌蛋白中的信号肽长度多集中于23~32个aa,上述蛋白质数量最多,所占比例为72.55% (图2)。

图1 枯草芽孢杆菌BEST195分泌蛋白氨基酸长度分析
Fig.1 Analysis of protein sequences with different length of amino acid inB.subtilissubsp.nattoBEST195
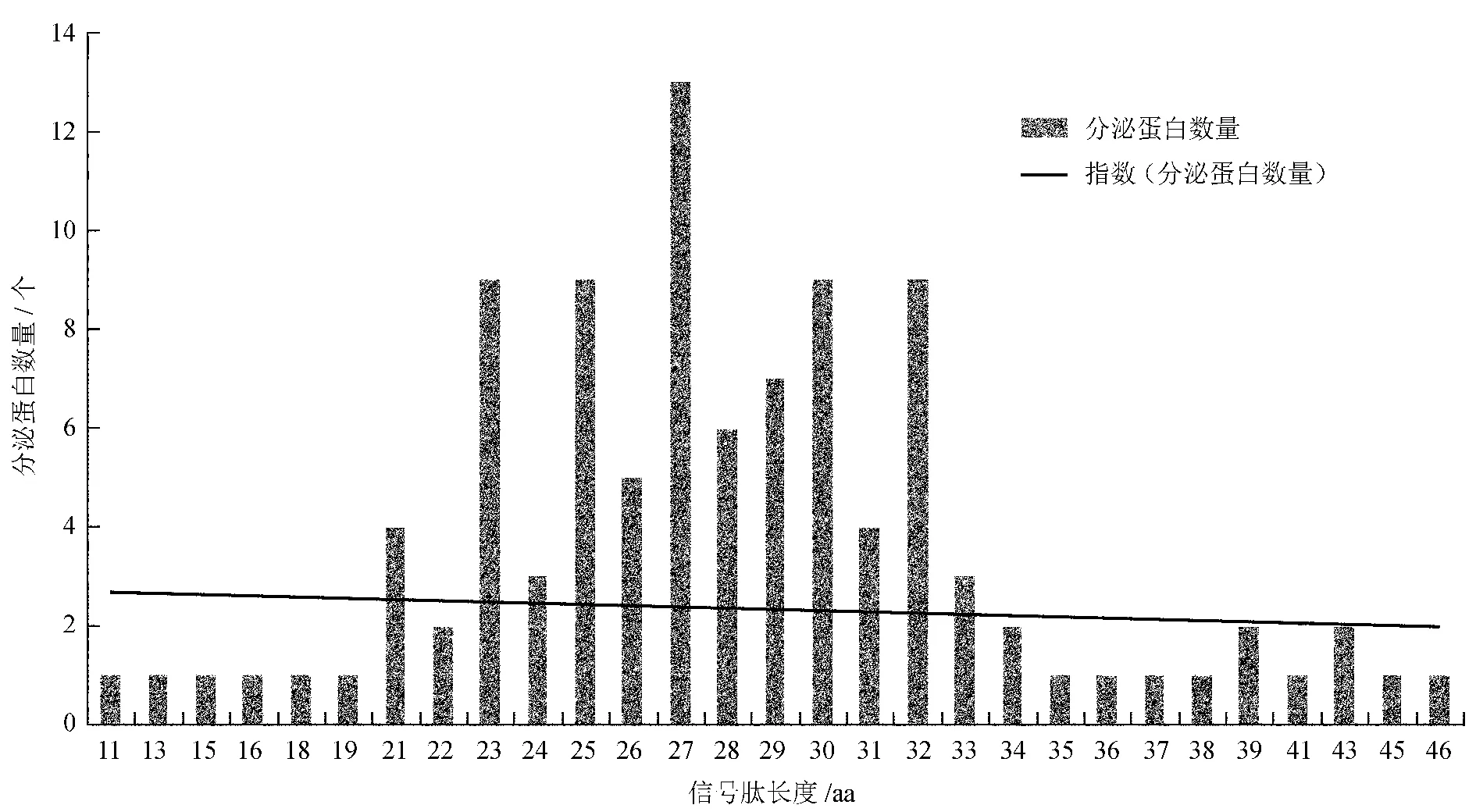
图2 枯草芽孢杆菌BEST195分泌蛋白信号肽长度分析
Fig.2 Analysis of protein sequences with different length of signal peptide inB.subtilissubsp.nattoBEST195
此外,对分泌蛋白信号肽的切割位点-3位到+3位进行统计分析,发现位于-3位、-2位、-1位、1位、2位、3位最多的氨基酸分别为A、S、A、A、E、K,所占比例分别为50.98%、26.47%、83.33%、32.35%、16.67%、13.73% (表1)。位于信号肽切割位点之前的-3位、-2位、-1位的氨基酸组成为A-S-A,属于SPⅠ型信号肽识别位点[18],与前人通过对致病疫霉 (Phytophthorainfestans)[8]、禾谷炭疽菌 (Colletotrichumgraminicola)[19]、粗糙脉孢菌 (Neurosporacrassa)[20]等分泌蛋白具有的信号肽酶切位点类型相同。上述分析结果表明,真菌、细菌、卵菌分泌蛋白进行信号肽序列识别、切割的信号肽酶类型多为Ⅰ型,具有物种保守性特点。
同时,对组成蛋白质的20种氨基酸在信号肽中的分布情况进行统计分析结果显示,A所占比例最大,为14.72%;其次为L,所占比例为14.44%;再次为K、S、M、V、F、I、G、T、C、R、N、P、Q、E、Y、W、H、D,所占比例分别为9.03%、9.03%、7.36%、7.22%、6.42%、6.35%、5.96%、5.76%、2.27%、1.95%、1.81%、1.78%、1.46%、1.26%、0.98%、0.80%、0.73%、0.66% (图3)。

表1 枯草芽孢杆菌BEST195分泌蛋白信号肽切割位点-3~+3氨基酸分布情况
图3 20种氨基酸在枯草芽孢杆菌BEST195分泌蛋白信号肽中所占比例情况
Fig.3 The frequency of 20 amino acid residues of signal peptides in secreted proteins inB.subtilissubsp.nattoBEST195
3 结论与讨论
近些年,学术界对植物病原真菌[6,7,19-21]、卵菌[8,22]等真核生物分泌蛋白的预测较多,同时,对属于革兰氏阴性细菌的植物病原细菌研究也较多[9-11],前期本研究小组也对一株生防菌——枯草芽孢杆菌XF-1的分泌蛋白进行了预测 (未发表数据)。
此外,前期研究发现,植物病原真菌、卵菌、细菌所含分泌蛋白的数量不等[19],所占比例也不同,其中,植物病原真菌分泌蛋白在全蛋白中的比例为3.65%~9.58%[19,23-24],卵菌所含分泌蛋白所占比例为2.96%~4.01%[8,22],细菌所含分泌蛋白所占比例为5.05%~12.09%[9-11,25]。除刘雅婷等[11]、陈建森等[25]对细菌分泌蛋白的预测较为宽泛,有待于进一步进行验证外,细菌分泌蛋白所占全蛋白的比例为5.05%~5.41%。与植物病原真菌、细菌、卵菌分泌蛋白所占比例不同,本研究发现的102个分泌蛋白,仅占全蛋白数量的2.29%,明显低于植物病原菌。同时,本研究小组明确褐环乳牛肝菌中的分泌蛋白所占比例也仅为1.79% (未发表数据),以及茎瘤固氮根瘤菌分泌蛋白所占比例为1.21%[26],结合本研究结果,推测与益生菌、植物外生菌根菌不同,植物病原菌在与植物长期的互作过程中,进化出较多的分泌蛋白,共同操控植物。
本研究利用诸多生物信息学预测程序总共获得102个分泌蛋白,明显低于植物病原真菌、细菌和卵菌中分泌蛋白的数量和比例,值得一提的是,上述研究所获得的分泌蛋白均为典型分泌蛋白,利用SecretomeP 2.0[27]对于4 251个不具有信号肽序列的蛋白进行非典型分泌蛋白预测,明确非典型分泌蛋白序列有1 342个 (未发表数据);同时,本研究所获得的典型分泌蛋白在氨基酸长度、信号肽长度方面也与前期所研究的植物病原菌报道不相同,而无论是植物病原真菌,还是植物病原细菌、卵菌,抑或是本研究的工业化纳豆生产菌,其信号肽酶切位点类型均为AXA,表明信号肽的酶切位点类型具有物种保守性特点。该研究为深入开展枯草芽孢杆菌发酵机制的解析提供了理论依据。
[1] Itaya M, Matsui K. Conversion ofBacillussubtilis168: Natto producingBacillussubtiliswith mosaic genomes[J]. Bioscience, Biotechnology and Biochemistry, 1999, 63(11): 2034-2037.
[2] Kamada M, Hase S, Fujii K, et al. Whole-genome sequencing and comparative genome analysis ofBacillussubtilisstrains isolated from non-salted fermented soybean foods[J]. PloS One, 2015, 10(10): e0141369.
[3] 蒋立文, 周传云, 黄香华. 纳豆菌的研究现状和应用进展[J]. 中国食物与营养, 2007(6): 24-26.
[4] 纪宁, 孔繁东, 祖国仁, 等. 纳豆菌抗菌作用的研究现状与展望[J]. 食品研究与开发, 2006, 27(1): 138-141.
[5] 尹聪, 许啸. 纳豆菌微生态制剂的研究进展[J]. 北方牧业, 2012(1): 14.
[6] 陈继圣, 郑士琴, 郑武, 等. 全基因组预测稻瘟菌的分泌蛋白[J]. 中国农业科学, 2006, 39(12): 2474- 2482.
[7] 田李, 陈捷胤, 陈相永, 等. 大丽轮枝菌 (VerticilliumdahliaeVdLs.17) 分泌组预测及分析[J]. 中国农业科学, 2011, 44(15): 3142-3153.
[8] 周晓罡, 侯思名, 陈铎文, 等. 马铃薯晚疫病菌全基因组分泌蛋白的初步分析[J]. 遗传, 2011, 33(7): 125-133.
[9] 肖文超, 范会云, 白亭亭, 等. 香蕉细菌性软腐病菌XJ8-3-3基因组中ORFs的信号肽及分泌蛋白功能预测分析[J]. 果树学报, 2014, 31(6): 1057-1064.
[10] 黄俊丽, 吴金钟, 肖崇刚, 等. 植物病原细菌RalstoniasolanacearumGMI1000中分泌蛋白信号肽分析[J]. 遗传, 2007, 29(11): 1409-1416.
[11] 刘雅婷, 李正跃, 朱有勇, 等. 植物病原细菌Pseudomonassyringaepv.tomato基因组中的信号肽分析[J]. 遗传, 2005, 27(6): 103-108.
[12] Nishito Y, Osana Y, Hachiya T, et al. Whole genome assembly of a natto production strainBacillussubtilisnatto from very short read data[J]. BMC Genomics, 2010, 11(1): 243.
[13] Kamada M,Hase S,Sato K, et al. Whole genome complete resequencing ofBacillussubtilisnatto by combining long reads with high-quality short reads[J]. PLoS One, 2014, 9(10): e109999.
[14] Petersen T N, Brunak S, von Heijne G, et al. SignalP 4.0: discriminating signal peptides from transmembrane regions[J]. Nat Methods, 2011, 8(10): 785-786.
[15] Käll L, Krogh A, Sonnhammer E L. Advantages of combined transmembrane topology and signal peptide prediction: the Phobius web server[J]. Nucleic Acids Research, 2007, 35(S2): 429-432.
[16] Juncker A S, Willenbrock H, Von Heijne G, et al. Prediction of lipoprotein signal peptides in Gram-negative bacteria[J]. Protein Science, 2003, 12(8): 1652-1662.
[17] Bendtsen J D, Nielsen H, Widdick D, et al. Prediction of twin-arginine signal peptides[J]. BMC Bioinformatics, 2005, 6(1): 167.
[18] Tjalsma H, Bolhuis A, Jongbloed J D, et al. Signal peptide-dependent protein transport inBacillussubtilis: a genome-based survey of the secretome[J]. Microbiology and Molecular Biology Reviews, 2000, 64(3): 515-547.
[19] 韩长志. 全基因组预测禾谷炭疽菌的分泌蛋白[J]. 生物技术, 2014, 24(2): 36-41.
[20] 周晓罡, 李成云, 赵之伟, 等. 粗糙脉孢菌基因组分泌蛋白的初步分析[J]. 遗传, 2006, 38(2): 200-207.
[21] 于钦亮, 马莉, 刘林, 等. 禾谷镰刀菌基因组中含寄主靶向模体分泌蛋白功能的初步分析[J]. 生物技术通报, 2008(1): 160-165, 180.
[22] 韩长志. 全基因组预测樟疫霉的候选效应分子[J]. 南京林业大学学报 (自然科学版), 2015, 39(2): 69-74.
[23] 陈琦光, 王陈骄子, 杨媚, 等. 希金斯刺盘孢全基因组候选效应分子的预测[J]. 热带作物学报, 2015, 36(6): 1105-1111.
[24] 陈相永, 陈捷胤, 肖红利, 等. 植物病原真菌寄生性与分泌蛋白组CAZymes的比较分析[J]. 植物病理学报, 2014, 44(2): 163-172.
[25] 陈建森, 佘菲菲. 全基因组预测幽门螺杆菌的分泌蛋白[J]. 中国人兽共患病学报, 2008, 24(7): 607-611.
[26] 王新新, 吴亮, 朱生凤, 等. 茎瘤固氮根瘤菌AzorhizobiumcaulinodansORS571基因组分泌蛋白的生物信息学分析[J]. 基因组学与应用生物学, 2012, 31(4): 394-400.
[27] Bendtsen J D, Kiemer L, Fausbøll A, et al. Non-classical protein secretion in bacteria[J]. BMC Microbiology, 2005, 5(1): 58.
(责任编辑 张 坤)
Prediction for Secreted Proteins fromBacillussubtilissubsp.nattoBEST195 Genome
Ren Wenlai1, Han Changzhi2, Liu Tong1, Xia Xiuqin1
(1. Agriculture Bureau of Wen′an County, Langfang Hebei 065800, China; 2. The Key Laboratory of Forest Disaster Warning and Control of Yunnan Province, College of Forestry, Southwest Forestry University, Kunming Yunnan 650224, China)
Bacillussubtilissubsp.nattoBEST195 is one of the important strains produced natto. To identify the secreted protein fromB.subtilisand clearly it′s characteristic. 4 456 protein sequences inB.subtiliswas analyzed to find the secreted protein using the program including SignalP, ProtComp, TMHMM, Phobius, LipoP and TatP. Meanwhile, the distribution of amino acids, the length of signal peptide as well as the signal peptide cleavage site of secreted protein was analyzed. 102 secrete proteins were found inB.subtilis. And the length of amino acids and the signal peptide were different the plant pathogens. The signal peptide cleavage site belongs to AXA type, which was same as other plant pathogenic fungi, bacteria and oomycete. Through the above bioinformatics analysis, the predicted secreted proteins can effectively achieve inB.subtilis, and the type of signal peptide cleavage site have consistent with other secreted proteins from different species.
Bacillussubtilissubsp.natto, secreted protein, signal peptide, prediction

10. 11929/j. issn. 2095-1914. 2016. 06. 017
2016-01-09
国家自然科学基金项目 (31560211) 资助;云南省森林灾害预警与控制重点实验室开放基金项目 (ZK150004) 资助;云南省优势特色重点学科生物学一级学科建设项目 (50097505) 资助;云南省高校林下生物资源保护及利用科技创新团队 (2014015) 资助;云南省教育厅科学研究基金项目 (2014Y330) 资助。
韩长志 (1981—),男,博士,副教授。研究方向:经济林木病害生物防治与真菌分子生物学。Email: hanchangzhi2010@163.com。
S759.81
A
2095-1914(2016)06-0106-06
第1作者:任文来 (1979—),男,学士。研究方向:农作物病虫害防治与新品种推广。Email: lfwazfd@126.com。
