木聚糖酶的研究进展
2016-12-14陈洪洋蔡俊林建国王常高杜馨
陈洪洋,蔡俊,林建国,王常高,杜馨
(湖北工业大学发酵工程教育部重点实验室,工业发酵湖北省协同创新中心,湖北武汉430068)
木聚糖酶的研究进展
陈洪洋,蔡俊*,林建国,王常高,杜馨
(湖北工业大学发酵工程教育部重点实验室,工业发酵湖北省协同创新中心,湖北武汉430068)
木聚糖是仅次于纤维素的第二丰富的可再生资源,通过筛选出产木聚糖酶的菌株,利用基因工程技术,将木聚糖酶基因进行异源表达,提高木聚糖酶产量。该文综述了菌株产生的木聚糖酶酶学性质,并对酶学性质进行改善使木聚糖酶更符合应用的条件,同时研究木聚糖酶分离纯化的步骤,来提高木聚糖酶的酶活。文章重点介绍了木聚糖酶的产生菌和木聚糖酶在基因工程技术等方面研究进展,并对木聚糖酶的分离纯化方法做了简要地介绍。
木聚糖酶;基因工程;酶学性质;分离纯化
木聚糖是植物半纤维素的主要组成部分,广泛存在于玉米芯、甘蔗渣、麦麸、秸秆等农作物废弃物中,由于木聚糖酶能够将木聚糖分解成不同长度的低聚木糖和木糖,该产物具有重要的经济价值,通过木聚糖酶将这些可利用资源充分利用,发挥其潜在的应用价值,木聚糖酶的研究也受到充分的重视。现在木聚糖酶已广泛应用于造纸、食品、饲料和能源等领域,带来了很大的经济效益。目前木聚糖酶的生产主要来源于微生物发酵,对发酵得到的木聚糖酶的酶学性质进行研究,以便了解该酶的应用范围,使木聚糖酶在工业应用中发挥其巨大的潜能,增加更多的经济效益,为各领域的发展做出更大的贡献。本文主要从微生物产木聚糖酶的种类和木聚糖酶的酶学性质,以及基因工程在木聚糖酶生产中的应用做了综述,对木聚糖酶的分离纯化进行了简要的介绍,旨在提高木聚糖酶的活力及扩展其应用范围。
1 木聚糖酶简介
木聚糖(xylan)是一种存在于植物细胞壁中的异质多糖,约占植物细胞干质量的15%~35%,是植物半纤维素(hemicellose)的主要成分,其含量非常丰富,是自然界中仅次于纤维素的多糖[1]。主要有O-乙酰基、4-O-甲基-D-葡糖醛酸残基、L-阿拉伯糖残基等[2]。
由于木聚糖组成的复杂性,木聚糖的生物降解也因此需要一个复杂的酶系统,通过其中各种组分的相互协同作用来降解木聚糖,因此木聚糖酶(xylanase,Xyl)是个复杂的酶系,主要包括β-1,4-内切木聚糖酶、β-木糖苷酶、α-L-阿拉伯糖苷酶、α-D-葡糖苷酸酶、乙酰基木聚糖酶和酚酸酯酶[3],可降解自然界中大量存在的木聚糖类半纤维素,在木聚糖水解酶系中,β-1,4-内切木聚糖酶是最关键的水解酶,它通过水解木聚糖分子的β-1,4-糖苷键,将木聚糖水解为小寡糖和木二糖等低聚木糖,以及少量的木糖和阿拉伯糖。β-木糖苷酶通过水解低聚木糖的末端来催化释放木糖残基。另外,参与彻底降解木聚糖的还有α-L-阿拉伯呋喃糖苷酶、α-葡萄糖醛酸苷酶、乙酰木聚糖酯酶,以及能降解木聚糖中阿拉伯糖侧链残基与酚酸(如阿魏酸或香豆酸)形成的酯键的酚酸酯酶等侧链水解酶,它们作用于木糖与侧链取代基之问的糖苷键,协同主链水解酶的作用,最终将木聚糖转化为它的组成单糖,各木聚糖酶的作用位点如表1所示[4]。

表1 木聚糖酶系组成及作用位点Table 1 Composition and action site of xylanase system
2 木聚糖酶的微生物来源及酶学性质
产木聚糖酶的微生物来源非常广泛,而且种类众多,用来生产木聚糖酶的微生物主要分为真菌和细菌(见表1)。真菌产木聚糖酶主要有曲霉菌(Aspergillus)、青霉菌(Penicillium)、木霉菌(Trichoderma)、酵母菌(Saccharomyces)等,细菌产木聚糖酶主要包括芽孢杆菌属(Bacillus)、链霉菌(Streptomyces)、大肠杆菌(Escherichia coli)等。酵母和大肠杆菌则常用作基因克隆的表达菌株。
在真菌产木聚糖酶方面,HE H等[5]采用米曲霉(Aspergillus oryzae)HML366利用甘蔗渣生产木聚糖酶XynH1,由于该酶有较好的热稳定性和pH耐受性,在造纸和生物能源领域有重要的应用。ANG S K等[6]发现烟曲霉菌SK1直接利用未经处理的油棕的树干,通过固态发酵产生纤维素酶、β-葡萄糖苷酶、外切葡聚糖酶和木聚糖酶,其中木聚糖酶活最高为418.70 U/g,通过这些酶的协同作用可以水解木质纤维素原料。DORRA S等[7]利用丝状真菌青霉occitanis Pol6产胞外的内切-β-1,4-木聚糖酶,该酶在酸性条件下的稳定性,可以用在饲料和食品领域。ZHANGH等[8]发现黄青霉固态发酵产木聚糖酶和β-甘露聚糖酶,使用粗酶液水解氨水溶液预处理过的玉米芯得到木糖最高产量为236.63 mg/g,还原糖的含量为553.94 mg/g,可以用该酶处理玉米芯得到木糖和还原糖,使玉米芯这样的农作物残渣充分利用。

表2 微生物来源的木聚糖酶及其酶学性质Table 2 Xylanase from microorganism and its enzyme properties
除了上述的真菌产木聚糖酶,还有细菌中的链霉菌和芽孢杆菌等也能产木聚糖酶。LUO L等[9]发现异硫链霉菌(Streptomyces althioticus)LMZM利用玉米芯采用液体深层发酵,该内切木聚糖酶,在pH 6.0~11.0和40~80℃范围内仍具有活性,在纸浆漂白过程中添加该酶后,提高了纸张亮度。MANDER P等[10]发现链霉菌属CS624使用农业残渣麦麸作为培养基底物,得到木聚糖酶EX624,该酶能将含有木质纤维素的农作物残渣水解成低聚木糖。DEESUKONW等[11]发现从烂稻草中筛选得到的链霉菌属SWU10,产生两种形式的内切木聚糖酶XynSW2A和XynSW2B,由于这两种酶的pH和热稳定性,可以利用稻杆生产生物能源,可以在食品、纺织、废水处理有广泛的应用。BAJAJ B K等[12]利用短小芽孢杆菌SS1以麦麸为唯一碳源,得到的木聚糖酶纯化后的比酶活为12.7 U/mg,该酶能利用麦麸作为底物产生相当大的木聚糖酶效价,表明麦麸可以使用作为一个廉价的底物生产木聚糖酶。GAUR R等[13]利用花域芽孢杆菌RSPP-15产生一种耐热的木聚糖酶,由于该酶的耐热性和耐碱稳定性,可以用在纸浆漂白、皮革制造、清洁剂等领域。WALIAA等[14]通过纤维化纤维微细菌产生内切-β-1,4-木聚糖酶,在50~60℃和pH 5~9范围内能保持稳定,在制浆造纸领域有很广泛的应用。
通过这些微生物产木聚糖酶的研究,菌株利用的底物偏向廉价的、易获得的原料来生产木聚糖酶,节省了生产成本,通过木聚糖酶水解农作物残渣来生产木糖或低聚木糖等,解决了资源浪费和农作物残渣造成的环境污染等问题,同时使资源充分利用。对于木聚糖酶活较高的菌株可以进一步的通过放大实验,用玉米芯、麦麸等原料来得到较理想的酶活和酶产量,为工业化大量生产提供了理论依据。而那些能产生多种酶的菌株,则可以通过融合蛋白的方法,来提高酶催化木质纤维素的效率。
3 木聚糖酶基因克隆及酶活力
随着基因工程技术的不断发展,将木聚糖酶基因克隆到大肠杆菌、酵母和其他菌株中进行高效表达也是研究的热点,木聚糖酶基因工程表达菌株和载体的选择情况见表3。
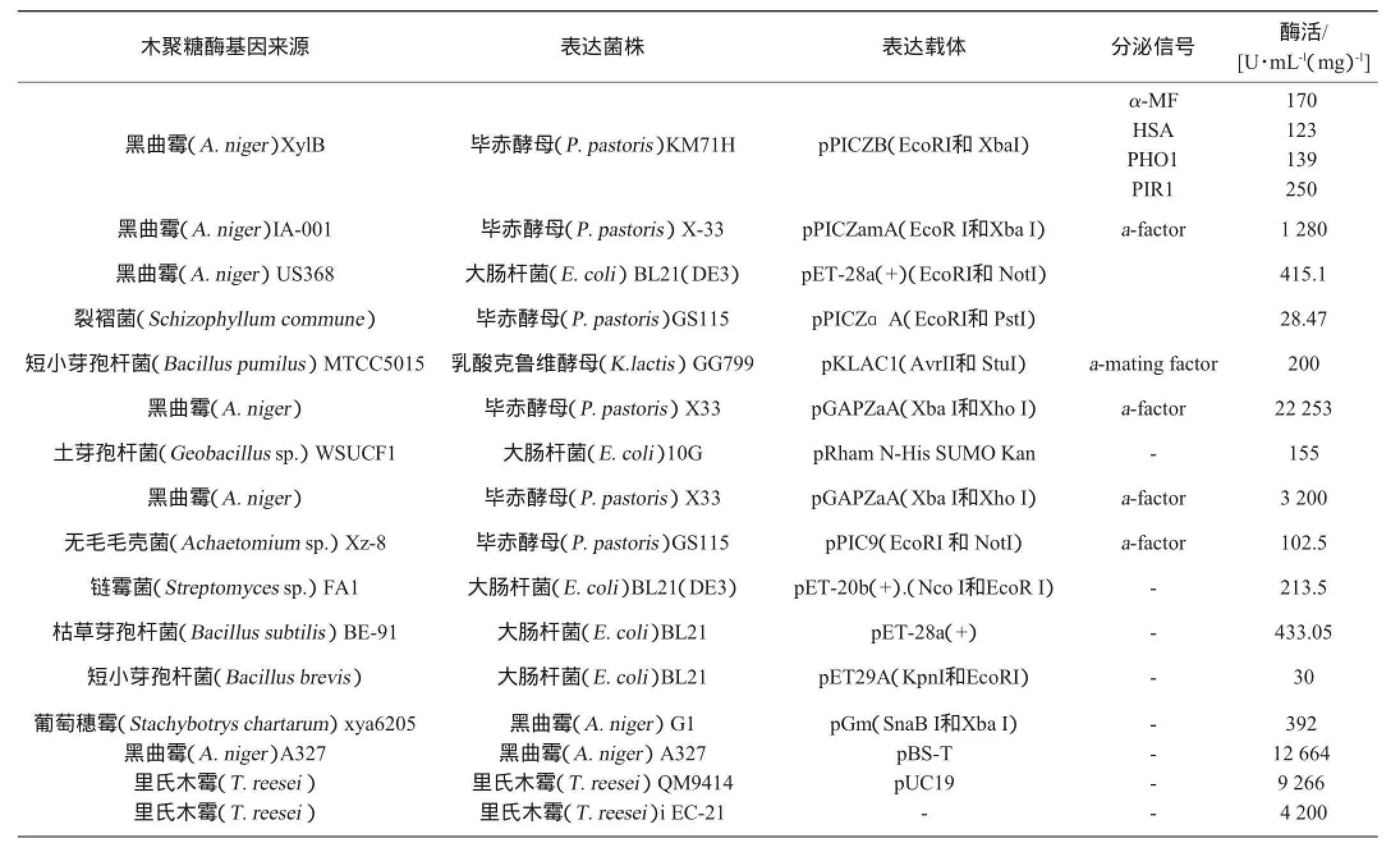
表3 木聚糖酶基因工程表达菌株和载体的选择Table 3 Xylanase gene engineering strains and choice of carrier
ELGHARBI F等[15]将黑曲霉US368的内切-β-1,4-木聚糖酶基因克隆到毕赤酵母(Pichia pastoris)SMD1168H中在组成型的GAP启动子下表达,得到木聚糖酶比原始菌株的酶活提高了3倍多。在面包制作过程中添加该酶后,降低了面团的吸水性,增加了生面团的延展性和体积。VERMA D等[16]将嗜热地芽孢杆菌的的木聚糖酶基因在大肠杆菌BL21(DE3)中表达,得到木聚糖酶的效价比野生型菌株高出了27倍。THOMAS L等[17]通过单因素试验和响应面分析,得到木聚糖酶的最高产量为200 IU/mL,酶活也有很大程度的提高。MIYAUCHI S等[18]将嗜热网球菌的木聚糖酶基因xynB克隆到里氏木霉RUT-C30,在egl2和cbh2糖基水解酶基因启动子下进行表达,结果分析木聚糖酶有不同程度的N-聚糖和O-聚糖修饰。木聚糖酶基因在大肠杆菌和毕赤酵母中表达后,酶活都有明显的提高,但是当木聚糖酶在进行异源表达时,会遇到木聚糖酶的表达后修饰等问题。
为了让木聚糖酶的酶学性质更加符合我们的期望,满足应用需求,在改善木聚糖酶的酶学性质方面,SOOZANIPOUR A等[19]将木聚糖酶共价连接在功能性的磁性纳米颗粒表面,酶活力、可重复利用性、热稳定性、pH稳定性、储存稳定性都优于原始的木聚糖酶。RAKOTOARIVONINAH等[20]从玉米秸秆中分离出的木聚糖酶(Tx-Xyn11),将该酶表面的疏水残基突变成非疏水性的,该酶的热稳定性和非特异性吸附率都大大的降低了,从侧面说明木聚糖酶表面的疏水残基和酶的热稳定性是密切相关的,为改善木聚糖的酶学性质提供了另一条途径。LI F等[21]对来自黑曲霉的木聚糖基因进行定点突变向碱性pH方向改造,与初始酶相比有0.5 pH单位的改变。HE L等[22]研究高温高压条件下的嗜热网球菌木聚糖酶二硫键突变后的酶学性质,发现该酶在500 MPa、80℃相当稳定。陈亚文等[23]在黑曲霉XZ-3S木聚糖酶的N-端区域引入半胱氨酸,定点突变后在大肠杆菌中表达,发现突变酶的最适温度比原始提高了5℃,在40℃和45℃时的半衰期都有延长了。RIBEIRO L F等[24]发现将木聚糖酶插入到木糖结合蛋白中,在有木糖存在时,酶活性提高了20%,原始的酶活性则被抑制了。李同彪等[25]通过在黑曲霉木聚糖酶XynZF-2α-螺旋催化活性中心位点处引入疏水性氨基酸,该酶的热稳定性得到大大提高。所以对木聚糖酶基因进行定点突变和固定化等方法,来改善木聚糖酶学性质,使木聚糖酶的酶学性质满足应用需求。
通过上述研究报道,可以通过酶的固定化、定点突变引入疏水性残基、二硫键和中性氨基酸等方法,来改善木聚糖酶的性质是有效的和可行性的,不同程度的改善了酶学性质,为以后的研究提供了理论依据和方法。而且将木聚糖酶基因转化到益生菌细胞中,作为食品和饲料添加剂,也会有较大的利用潜能。
4 木聚糖酶分离纯化方法及酶学性质
通过菌株发酵得到木聚糖酶后,由于木聚糖降解酶系比较复杂,有的菌株还同时产生纤维素酶等其他酶,这些酶的性质相近,给木聚糖酶的分离纯化带来较多的困难。随着分离技术的发展,木聚糖酶的分离纯化方法由非特异性方法发展为特异性。非特异性分离纯化方法包括硫酸铵分级沉淀,脱盐后经过离子交换、凝胶过滤层析和疏水层析等的方法,有的还要经过超滤浓缩等步骤。特异性分离纯化方法,主要是根据木聚糖酶与底物的特异性结合而发展的亲和层析方法(见表4)。

表4 木聚糖酶分离纯化方法及其酶学性质Table 4 Methods of xylanase separation and purification and its enzyme properties

续表
硫酸铵分级沉淀常用做分离纯化木聚糖酶第一步,但是高浓度的硫酸铵对木聚糖酶活力的测定存在干扰,测定酶活时可以通过透析或稀释的方法来降低盐的浓度,排除干扰。经过硫酸铵分级沉淀后得到的木聚糖酶沉淀,有的先要经过缓冲液溶解、透析、脱盐,然后进行凝胶过滤层析和离子交换层析。当使用凝胶过滤分离纯化木聚糖酶时,要考虑酶与凝胶介质(琼脂糖或葡聚糖)的相互作用,木聚糖酶结构中可能含有纤维素结合区,而葡聚糖是由α-葡聚糖残基构成的,两者可以相互结合,使凝胶过滤后的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)测定分子质量可能出现异常[26]。超滤在生物大分子的脱盐、脱水和浓缩时,能较好地保持生物大分子活性的优点,但是KOCABAS D S等[27]在分离纯化嗜热色串孢(Scytalidium thermophilum)ATCC No.16454产生的木聚糖酶时采用超滤浓缩,但是发现纤维素超滤膜有部分的降解,经测定发现粗酶液中含有较低的纤维素酶活为0.42 IU/mL,使超滤浓缩在木聚糖酶的分离纯化中的使用受到影响。双水相萃取体系条件温和、没有有机溶剂的残留问题、可按比例放大和连续操作等优点,KOCABAS D S等[27]分离纯化嗜热色串孢ATCC No.16454产生的木聚糖酶时,采用的是TX-114/水体系。FU G等[28]在分离纯化黑曲霉XZ-3S的木聚糖酶基因在大肠杆菌BL21(DE3)表达的木聚糖酶,采用的就是Ni2+-螯合剂亲和层析,经过该步骤的纯化后的酶液在SDS-PAGE显示的是单一的条带,说明木聚糖酶已高度纯化,经过亲和层析后即达到了精制纯化的目的,说明该方法具有高效性。木聚糖酶除了采用上述常用的分离纯化方法外,还有电洗脱、三相萃取、反微团萃取和置换层析[29]等不常用的分离纯化方法。
由于木聚糖酶是一个复杂的酶系组成的,而且菌株在发酵过程中产生的酶不是单一的,所以在分离纯化过程中,会将一些可能有协同作用的酶去除掉,反而使酶的回收率和酶活降低。KOKCHANG L等[30]在分离纯化青霉rolfsii c3-2(1)IBRL产生的木聚糖酶时,采用了四步分离纯化方法(硫酸铵分级沉淀、阴离子交换层析、凝胶过滤、疏水层析)经过阴离子交换层析时显示的有多个活性的峰,但是经过凝胶过滤层析只出现单一的木聚糖酶活性峰,可能使与木聚糖酶有协同作用的酶被去除掉,使酶的回收率降低了[31],这对完全水解木聚糖是不利的。在分离纯化木聚糖酶时,纯化步骤最好不要超过三步,而且要尽可能选择合适的分离纯化方法,使木聚糖酶的回收率和酶活保持较高的水平。
5 展望
由于现在生产的木聚糖酶多来自于菌株发酵,所得的木聚糖酶力较低,酶活性范围窄,限制了木聚糖酶的广泛应用。可以通过筛选高产木聚糖酶菌株和优化培养基提高木聚糖酶的生产水平,在此基础上选择成本较低的原料,减少生产成本。也可以通过基因工程技术提高木聚糖酶的表达水平,结合定点突变技术改善木聚糖酶的酶学性质,使木聚糖酶更适合不同领域的应用要求。除此之外,研究分离纯化对木聚糖酶的活性和回收率的影响,选择合适的分离纯化方法,对提高木聚糖酶活有很大帮助。
[1]BIELY P.Microbial xylanolytic systems[J].Trends Biotechnol,1985,3 (11):286-290.
[2]张红莲,姚斌,范云六.木聚糖酶的分子生物学及其应用[J].生物技术通报,2002(3):23-26.
[3]SUNNA A,ANTRANIKIAN G.Xylanolytic enzymes from fungi andbacteria[J].Crit Rev Biotechnol,2008,17(1):39-67.
[4]RENATO C,PAULINA B,JAIME E.The xylanolytic enzyme system from the genusPenicillium[J].J Biotechnol,2006,123(4):413-33.
[5]HE H,QIN Y,LI N,et al.Purification and characterization of a thermostable hypothetical xylanase fromAspergillus oryzaeHML366[J]. Appl Biochem Biotechnol,2015,175(6):3148-3161.
[6]ANG S K,SHAZA E M,ADIBAH Y,et al.Production of cellulases and xylanase byAspergillus fumigatusSK1 using untreated oil palm trunk through solid state fermentation[J].Process Biochem,2013,48(9):1293-1302.
[7]DORRA S,FATMA B,MARIEM S,et al.Purification and properties of a thermostable xylanase GH 11 fromPenicillium occitanisPol6[J].Appl Biochem Biotechnol,2012,168(4):851-863.
[8]ZHANG H,SANG Q.Production and extraction optimization of xylanase and β-mannanase byPenicillium chrysogenumQML-2 and primary application in saccharification of corn cob[J].Biocheml Eng J,2015,97: 101-110.
[9]LUO L,CAI J,WANG C,et al.Purification and characterization of an alkaliphilic endo-xylanase fromStreptomyces althioticusLMZM and utilization in pulp industry[J].J Cheml Tech Biotechnol,2015,91(4):165-171.
[10]MANDER P,YUN H C,PRADEEP G C,et al.Biochemical characterization of xylanase produced fromStreptomycessp.CS624 using an agro residue substrate[J].Process Biochem,2014,49(3):451-456.
[11]DEESUKON W,NISHIMURA Y,HARADA N,et al.Purification, characterization and gene cloning of two forms of a thermostable endo-xylanase fromStreptomycessp.SWU10[J].Process Biochem, 2011,46(12):2255-2262.
[12]BAJAJ B K,KHAJURIA Y P,SINGH V P.Agricultural residues as potential substrates for production of xylanase from alkali-thermotolerant bacterial isolate[J].Biocatal Agr Biotechnol,2012,1(4):314-320.
[13]GAUR R,TIWARI S,RAI P,et al.Isolation,production,and characterization of thermotolerant xylanase from solvent tolerantBacillus vallismortisRSPP-15[J].Int J Polym Sci,2015(14):655-661.
[14]WALIA A,MEHTA P,CHAUHAN A,et al.Purification and characterization of cellulase-free low molecular weight endo β-1,4 xylanase from an alkalophilicCellulosimicrobium cellulansCKMX1 isolated from mushroom compost[J].World J Microb Biot,2014,30(10):2597-2608.
[15]ELGHARBI F,HMIDA-SAYARI A,ZAAFOURI Y,et al.Expression of anAspergillus nigerxylanase in yeast:Application in breadmaking andin vitrodigestion[J].Int J Biol Macromol,2015,79:103-109.
[16]VERMA D,SATYANARAYANA T.Cloning,expression and applicability of thermo-alkali-stable xylanase ofGeobacillus thermoleovorans in generating xylooligosaccharides from agro-residues[J].Bioresource Technol,2011,107(2):333-338.
[17]THOMAS L,SINDHU R,BINOD P,et al.Production of an alkaline xylanase from recombinantKluveromyces lactis(KY1)by submerged fermentation anditsapplicationinbio-bleaching[J].Biochem Eng J,2015, 102:24-30.
[18]MIYAUCHI S,TE'O V S,BERGQUIST P L,et al.Expression of a bacterial xylanase inTrichoderma reeseiunder theegl2andcbh2glycosyl hydrolase gene promoters[J].New Biotechnol,2013,30(5):523-530.
[19]SOOZANIPOUR A,TAHERI-KAFRANI A,ISFAHANI A L.Covalent attachment of xylanase on functionalized magnetic nanoparticles and determination of its activity and stability[J].Chem Eng J,2015,270: 235-243.
[20]RAKOTOARIVONINA H,HERMANT B,AUBRY N,et al.Engineering the hydrophobic residues of a GH11 xylanase impacts its adsorption onto lignin and its thermostability[J].Enzyme Microb Tech,2015,81: 47-55.
[21]LI F,XIE J,ZHANG X,et al.Improvement of the optimum pH ofAspergillus nigerxylanase towards an alkaline pH by site-directed mutagenesis[J].J Microb Biotech,2014,25(1):11-17.
[22]HE L,VOUTILAINEN S,OJAMO H,et al.Stability and activity of Dictyoglomus thermophilumGH11 xylanase and its disulphide mutant at high pressure and temperature[J].Enzyme Microb Tech,2015,70: 66-71.
[23]陈亚文,周晨妍,谢晶晶,等.引入中性氨基酸对木聚糖酶XynZF-2热稳定性的影响[J].中国酿造,2015,34(10):27-31.
[24]RIBEIRO L F,NICHOLES N,TULLMAN J,et al.Insertion of a xylanase in xylose binding protein results in a xylose-stimulated xylanase [J].Biotechnol Biofuel,2015,8(1):1-15.
[25]李同彪,周晨妍,朱新术,等.引入疏水性氨基酸对GH11家族木聚糖酶热稳定性的影响[J].食品工业科技,2016,37(1):145-152.
[26]NINAWE S,KAPOOR M,KUHAD R C.Purification and characterization of extracellular xylanase fromStreptomyces cyaneusSN32[J].Bioresource Technol,2008,99(5):1252-1258.
[27]KOCABAS D S,GUDER S,OZBEN N.Purification strategies and propertiesofalow-molecularweightxylanaseanditsapplication in agricultural waste biomass hydrolysis[J].J Moleculr Catal B-Enzym, 2015,115:66-75.
[28]FU G,WANG Y,WANG D,et al.Cloning,expression,and characterization of an GHF 11 xylanase fromAspergillus nigerXZ-3S[J].Indian J Microb,2012,52(4):682-688.
[29]张小勇.一种内切木聚糖酶的亲和纯化及性质研究[D].上海:上海交通大学,2008.
[30]KOKCHANG L,ARAI T,IBRAHIM D,et al.Purification and characterization of a xylanase from the newly isolatedPenicillium rolfsiic3-2 (1)IBRL[J].Bioresources,2015,10(1):1627-1643.
[31]LUCENA-NETO S D A,FERREIRA-FILHO E X.Purification and characterizationofanewxylanasefromHumicolagriseavar.thermoidea [J].Braz J Microbiol,2004,35(1-2):86-90.
Research progress of xylanase
CHEN Hongyang,CAI Jun*,LIN Jianguo,WANG Changgao,DU Xin
(Hubei Provincial Cooperative Innovation Center of Industrial Fermentation,Key Laboratory of Fermentation Engineering, Ministry of Education,Hubei University of Technology,Wuhan 430068,China)
Abstarct:Xylan is the second most abundant renewable resources after cellulose.By screening of xylanase-producing strains,using the genetic engineering technology and the heterologous expression of xylanase gene,the production of xylanase was improved.The enzymatic properties of xylanase produced by the strain were researched and improved to make xylanase more suitable for application conditions.In the meantime,the steps of separation and purification of xylanase were researched to increase the xylanase activity.The xylanase-producing bacteria and the research progress of xylanase in genetic engineering technology were introduced emphatically,and the separation and purification of xylanase were introduced briefly.
xylanase;genetic engineering;enzymatic property;separation and purification
Q814.9
0254-5071(2016)11-0001-06
10.11882/j.issn.0254-5071.2016.11.001
2016-05-18
国家自然科学基金(31401807)
陈洪洋(1991-),男,硕士研究生,研究方向为微生物发酵。
*通讯作者:蔡俊(1968-),男,博士,教授,研究方向为发酵工程。
