悬浮物对褐牙鲆肌肉抗氧化酶活性及相关基因表达的影响*
2016-12-09肖广侠徐文远郑德斌
肖广侠,徐文远,贾 磊*,郑德斌,张 博,马 超
(1.天津渤海水产研究所, 天津 300457;2.临沂出入境检验检疫局,山东 临沂 276034)
悬浮物对褐牙鲆肌肉抗氧化酶活性及相关基因表达的影响*
肖广侠1,徐文远2,贾 磊1*,郑德斌1,张 博1,马 超1
(1.天津渤海水产研究所, 天津 300457;2.临沂出入境检验检疫局,山东 临沂 276034)
为探究褐牙鲆(Paralichthysolivaceus)幼鱼在悬浮物胁迫下的耐受机制,以两种规格褐牙鲆为实验材料,在悬浮物质量浓度为5 000,10 000 mg/L胁迫下,研究了褐牙鲆肌肉中总超氧化物歧化酶(T-SOD)、谷胱甘肽转移酶(GST)、过氧化氢酶(CAT) 酶活力及SOD2、GST、CAT的mRNA转录水平的变化情况。实验结果显示:T-SOD酶活力具有一定的变化趋势,B幼鱼T-SOD酶活力呈现先升高、再降低、再升高的趋势,SOD2基因相对表达量在第48小时达到峰值(P<0.05),两种幼鱼T-SOD基因相对表达变化趋势在第72小时和第96小时相反;CAT酶活力呈现先升高、再降低的趋势,A幼鱼及5 000 mg/L实验组B幼鱼CAT基因相对表达与CAT酶活力变化趋势一致;B幼鱼GST酶活力实验组低于对照组(P<0.05),GST基因相对表达量在第24小时和第48小时实验组高于对照组(P<0.05)、在第72小时和第96小时实验组与对照组无显著差异。研究结果表明:悬浮物对褐牙鲆抗氧化酶活力及相关基因的表达具有一定的影响;不同体长规格的褐牙鲆抗氧化酶活力及相关基因的表达存在差异;T-SOD酶活力、GST酶活力与相关基因表达量变化趋势不一致,CAT酶活力与相关基因表达量变化趋势一致。本研究可为揭示褐牙鲆应对悬浮物胁迫的耐受机制及褐牙鲆耐悬浮物品种选育提供基础数据。
褐牙鲆;悬浮物;抗氧化系统酶;基因表达
褐牙鲆(Paralichthysolivaceus)隶属于鲽形目(Pleuronectiformes)、牙鲆科(Paralichthyidae)、牙鲆属(Paralichthys),属于肉食性底栖鱼类,具有变态发育过程。褐牙鲆主要分布于中国黄渤海及朝鲜、日本沿岸水域,是我国重要的海水增养殖鱼类[1]。近几年来,人工增殖放流恢复海洋中的生物资源的措施日益受到重视,仅2014年在天津渤海湾海区放流的中国对虾达到14亿尾,褐牙鲆达到147.2万尾。渤海由于宽陆岸浅海的特性及其特殊地理位置,再加上陆地众多河流汇入,使其成为中国悬浮物质量最高的海域[2];另外,随着我国海洋经济的发展,港口码头建设及填海造地等海洋工程越来越多,在建设施工过程中产生大量水体底泥悬浮,导致海域悬浮物浓度大幅度升高,改变了水体的物理、化学环境[3]。将人工培育的水产苗种直接投放到海域中,能否适应海域自然水域环境,尤其褐牙鲆机体对悬浮物胁迫的耐受机制,在这方面尚无报道。
抗氧化系统酶是生物体内应对不利环境条件的重要物质,在生物处于不适宜的生存条件下,抗氧化系统各种酶的活力会发生变化,来应对外界的环境胁迫,这对环境污染有一定的生物指示作用[4]。目前针对影响褐牙鲆抗氧化系统酶活力的研究集中于重金属及环境因子,王凡等[5-6]研究表明SOD,CAT,GSH-PX 活性变化可以反映牙鲆在铜污染下的伤害程度,3种酶活性均表现为低浓度诱导、高浓度抑制;Huang等[7]和Cao等[8]分别研究了重金属汞(Hg)及镉(Cr)对牙鲆的抗氧化系统酶活力的影响及重金属在体内的富集情况;王茂林等[9]研究揭示了过高浓度钙(Ca)、镁(Mg)对褐牙鲆幼鱼肝脏的SOD和CAT活性产生一定抑制作用;柳意樊[10]研究了不同升温模式下,褐牙鲆和杂交鲆SOD,CAT,GSH,LZM和PK相对活力的变化情况;黄伟[11]研究了Hg,Pb,Zn对褐牙鲆仔稚鱼SOD,CAT,MDA及GSH的活性的影响;Lou等[12]认为高水温对褐牙鲆幼鱼肝脏中SOD及CAT酶活性产生显著影响,褐牙鲆幼鱼肝脏中SOD及CAT酶活力与幼鱼生长相关性显著。徐冬冬等[13]研究表明褐牙鲆肝脏中SOD及CAT活力随胁迫温度升高而降低。在悬浮物对渔业生物抗氧化酶影响方面,目前也出现较多报道。张雪等[14]研究发现菲律宾蛤仔鳃丝SOD活力对悬浮物胁迫的反应更迅速;杨国军等[15]研究发现四角蛤蜊鳃丝抗氧化酶活力会对悬浮物胁迫快速更灵敏;蒋玫等[16]研究了底泥浸出液对脊尾白虾抗氧化酶相关基因mRNA 转录水平表达在一定时间内有一定的诱导作用;王广军等[17]的研究表明底泥悬浮物质量浓度对杂色鲍的生理生化存在一定影响。但是对悬浮物对褐牙鲆抗氧化系统影响方面的研究尚无报道。
针对以上情况,本实验以两种规格褐牙鲆幼鱼为实验材料,研究其在不同悬浮物质量浓度下体内总超氧化物歧化酶(T-SOD)、谷胱甘肽S转移酶(GST)、过氧化氢酶(CAT) 酶活力及SOD2,GST,CAT的mRNA转录水平表达变化情况,分析了褐牙鲆幼鱼底泥悬浮物的耐受免疫调节机制,以期为褐牙鲆的抗逆品种选育及增殖放流提供理论依据,为进一步评估海洋工程对放流生物的损害情况提供参考依据。
1 材料与方法
1.1 实验材料
1.1.1 实验鱼
实验选用的褐牙鲆幼鱼取自于天津市兴盛水产有限公司,共分两种规格:A幼鱼体长(4.21±0.41) cm,B幼鱼体长(14.53±1.58) cm。本研究挑选体质健康、大小均匀的个体用于实验,两种幼鱼分别使用60条。
1.1.2 实验用水
实验用水为高盐卤水兑淡水,经过沉淀、砂滤、次氯酸钠(NaClO)消毒、筛绢过滤等处理。水温:(22±1) ℃;盐度:25~28;pH:8.0~8.2;OD:5.8~7.5 mg/L;NH4-N:0.012~0.248 mg/L;NO2-N:0.005~0.011 mg/L;NO3-N:0.287~0.363 mg/L;PO4-P:0.012~0.025 mg/L。
1.1.3 实验底泥
实验用悬浮物泥样采自天津大神堂海域(117°55′18″E,39°09′01″N;位于汉沽浅海生态系统海洋特别保护区,在整个保护区内具有代表性)的底泥表层,实验室内通风晾干,然后放于烘箱中(80 ℃)烘干至恒重,冷却后研磨处理,用100目筛绢过筛,放置于干燥器中低温保存。实验所用沉积物中各成分含量检测参照《海洋监测规范》[18]进行,分析结果见表1。其主要成分含量均符合《海洋沉积物质量标准》[19]第一类要求,不会对受试生物产生毒害作用。
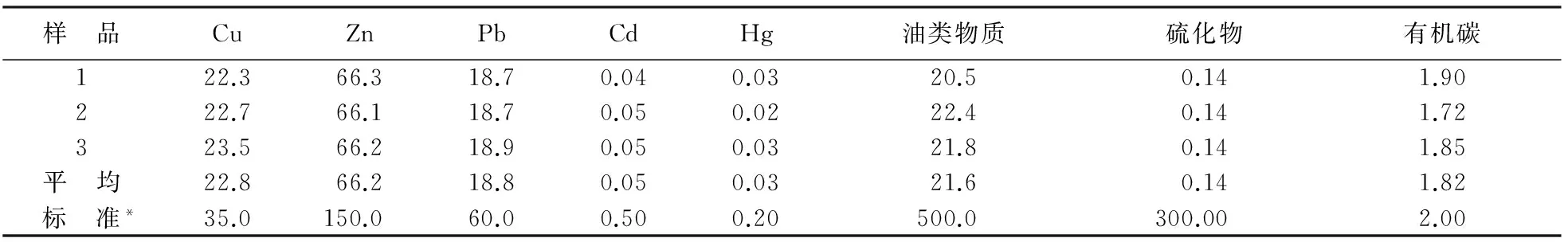
表1 实验底泥主要成分含量(mg/kg)Table 1 Contents of major components in the bottom sediment used for the experiments(mg/kg)
1.2 实验方法
1.2.1 悬浮物溶液的配置方法
将烘干泥样与海水按需浓度配置悬浮物溶液,实验前充分搅拌,使底泥充分溶解为悬浮物。
1.2.2 悬浮物粒径的测量
悬浮物粒径使用维纳2008D激光粒度仪测量,底泥粒径X80=12.70 μm,粒径X<5 μm所占的比例为29.98%,具体粒度特征见表2。
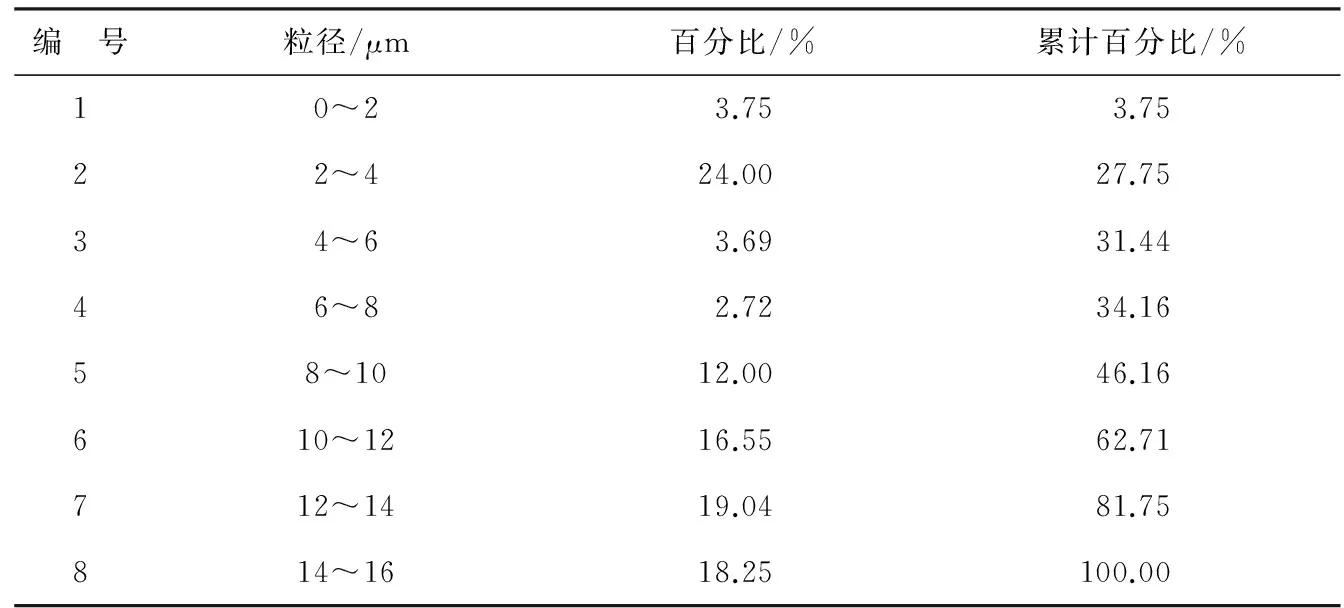
表2 实验用底泥粒径特征Table 2 Grain size characteristics of the bottom sediment used in the experiments
1.2.3 实验设计
实验参照国家标准《水质·物质对淡水鱼(斑马鱼)急性毒性测定方法》[20]进行,褐牙鲆幼鱼暂养24 h后开始实验,悬浮物溶液浓度设置为5 000和10 000 mg/L,每个质量浓度均设置5个平行组,另外设置相应的对照组(不加泥样),悬浮物浓度认为0 mg/L。实验容器为15 L透明整理箱,每个整理箱加入各质量浓度悬浮物溶液10 L,放入实验鱼4尾,整个实验过程不投饵。采用气石均匀充气及人工定时搅拌等方法,使底泥始终处于悬浮状态。实验期间不换水。悬浮物胁迫24,48,72,96 h后分别从每个实验组随机挑选3尾幼鱼取样。
1.2.4 样品处理
剪取褐牙鲆幼鱼背部肌肉,按照1∶10(W/V)加入预冷PBS缓冲溶液(pH=6.4)放置冰上以玻璃匀浆器匀浆,离心(4 ℃,800 g, 10 min)取上清液置-80 ℃冰箱保存用于3种抗氧化酶活性的测定,另-80 ℃冰箱保存肌肉样本用于抗氧化酶相关基因表达的测定。
1.2.5 抗氧化酶活性的测定
T-SOD,GST,CAT酶活力测定均根据试剂盒说明书进行(试剂盒购自南京建成生物工程研究所)。其中37 ℃时,每分钟每毫克组织蛋白使反应体系的吸光度(OD)值增加0.01 L/θ·cm时,定义为1个总抗氧化能力(T-SOD)单位;每毫克组织蛋白质,每分钟扣除非酶反应的作用,使反应体系中GSH浓度降低1 μmol/L为一个GST酶活力单位;每毫克组织蛋白每秒钟分解1 μmol的H2O2的量为1个CAT活力单位[21]。所有指标吸光值均使用UV-7504单光束紫外可见分光光度计测定。
1.3 抗氧化酶基因的表达
1.3.1 引物设计
根据褐牙鲆SOD2,CAT,GST的cDNA序列,以Primer 5.0设计Real-time PCR 扩增特异性引物,选择基因稳定表达及片段大小适合的β-actin作为内参基因,引物序列详见表3。引物合成和cDNA 序列测定委托上海生工生物工程有限公司完成。

表3 Real-time PCR 扩增的特异性引物Table 3 Specific primers used for the real-time PCR amplification
1.3.2 总RNA提取cDNA第一链的合成
将保存于-80 ℃冰箱的样品取出后于冰上融化,并按照TRizol试剂说明书提取总RNA,RNA 沉淀用DEPC水溶解,用核酸定量仪(Thermo Scientific)测定260和280 nm处的吸收值,检测RNA的产量和纯度,并使用1%琼脂糖凝胶进行RNA非变性电泳检测RNA 的完整性。取等量(2 μg)的RNA,按照All-in-OneTMFirst-Strand cDNA Synthesis Kit(GeneCopoeiaCat.No.AORT-0050)说明书反转录各组织的总RNA,合成cDNA 第一链。
1.3.3 Real-time PCR 扩增
采用Real-time PCR(SYBR Green)2-△△Ct相对定量方法,按照Platinum®SYBR®Green qPCR SuperMix-UDG试剂盒说明书进行操作,反应体系如下:SYBR®Premix Ex Super Mix (2×) 10.0 μL,Forward primer(10 μmol/L) 2.0 μL,Reverse primer(10 μmol/L) 2.0 μL,PCR板每个孔分别加入:DNA 1.0 μL,ddH2O 5.0 μL,反应总体积共20 μL。将样品在PCR 管内混匀后分装入96孔PCR板(Axygen)中,瞬时离心后放入ABI 7500 Fast荧光定量PCR仪中进行PCR 扩增,反应程序为:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火30 s,72 ℃延伸30 s,共40 个循环;熔解曲线条件为95 ℃ 15 s,60 ℃ 1 min,95 ℃ 15 s, 60 ℃ 15 s。反应完成后,用ABI 7500 system软件分析结果。
1.3.4 Comparative Delta-delta Ct相对定量
由标准曲线可知,目的基因与内参扩增效率基本一致,均在95%以上,符合Comparative Delta-delta Ct 相对定量计算要求,Comparative Delta-delta Ct相对定量参考Livak和Schmittgen的方法[22],目的基因的表达量定义为待测样品与对照组中目的基因的表达差异倍数(Fold change),计算公式为

1.4 实验数据的处理
数据统计使用软件SPSS 22.0 进行,所得数据均以平均值±标准差(x±SD)表示,使用单因素方差分析ANOVA进行显著性检验(P<0.05)。
2 结果与分析
2.1 悬浮物胁迫对褐牙鲆幼鱼T-SOD 酶及基因表达的影响
悬浮物对褐牙鲆幼鱼肌肉T-SOD酶活力的影响见图1。实验结果表明:悬浮物对褐牙鲆A幼鱼在第72和第96小时时T-SOD酶活力具有诱导作用,5 000和10 000 mg/L A幼鱼T-SOD酶活力分别在第96小时和第24小时达到峰值(P<0.05);在悬浮物胁迫下,B幼鱼T-SOD酶活力呈现先升高、再降低、再升高的趋势,并且在第96小时时达到峰值(P<0.05);悬浮物对两种幼鱼T-SOD酶活力在第72小时影响趋势相反,在第96小时诱导趋势一致。
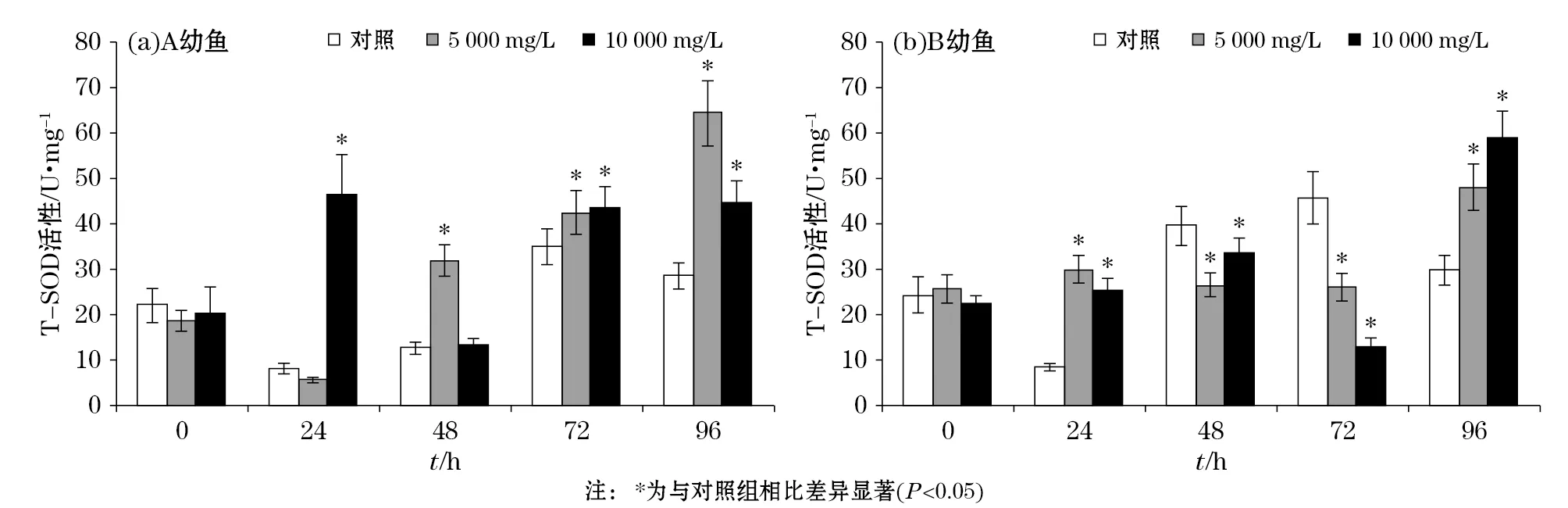
图1 褐牙鲆幼鱼体内T-SOD活性随悬浮物胁迫时间的变化Fig.1 Changes of the T-SOD activity in the body of P. olivaceus with the time of SS stress
悬浮物对褐牙鲆幼鱼肌肉内SOD2基因相对表达的影响见图2,在5 000 mg/L实验组,A幼鱼SOD2基因相对表达在24~96 h具有升高的趋势(P<0.05),B幼鱼SOD2基因相对表达在24~96 h呈现先降低、再升高、再降低的趋势;在10 000 mg/L实验组,A幼鱼SOD2基因相对表达在48~96 h呈现升高趋势(P<0.05),B幼鱼SOD2基因相对表达在48~96 h呈现先升高、再降低的趋势。两种幼鱼T-SOD基因相对表达都在第48小时达到峰值(P<0.05)。A幼鱼T-SOD酶活力与基因表达量在第72小时和第96小时实验组都呈现增高趋势(P<0.05),B幼鱼T-SOD酶活力与基因表达量在第48小时和第96小时变化趋势相反,在第72小时诱导趋势一致。

图2 褐牙鲆幼鱼体内SOD2基因相对表达量随悬浮物胁迫时间的变化Fig.2 Changes of the relative gene expression of T-SOD in the body of P. olivaceus with the time of SS stress
2.2 悬浮物胁迫对褐牙鲆CAT酶及基因表达的影响
悬浮物对褐牙鲆幼鱼肌肉内CAT酶活力的影响见图3,实验结果表明:悬浮物胁迫下,两种规格的幼鱼体内CAT酶活力均呈现先升高、后降低的趋势,在第24小时和第48小时实验组幼鱼CAT酶活力高于对照组(P<0.05),在第72小时和第96小时实验组幼鱼CAT酶活力低于对照组(P<0.05),5 000和10 000 mg/L实验组对两种规格幼鱼CAT酶活力在各时间点的诱导或者抑制趋势一致。

图3 褐牙鲆幼鱼体内CAT活性随悬浮物胁迫时间的变化Fig.3 Changes of the CAT activity in the body of P. olivaceus with the time of SS stress
悬浮物对褐牙鲆幼鱼肌肉内CAT基因相对表达的影响见图4,在悬浮物胁迫下,A幼鱼及5 000 mg/L实验组B幼鱼CAT基因表达量在24~96 h呈现先升高后降低趋势,并且第24小时和第48小时实验组高于对照组(P<0.05),第72小时和第96小时实验组低于对照组,CAT基因相对表达量在第24小时达到峰值(P<0.05)。10 000 mg/L实验组B幼鱼第72小时和第96小时CAT基因相对表达量实验组低于对照组(P<0.05),这与CAT酶活力的变化趋势相同。

图4 褐牙鲆幼鱼体内CAT基因相对表达量随悬浮物胁迫时间的变化Fig.4 Changes of the relative gene expression of CAT in the body of P. olivaceus with the time of SS stress
2.3 悬浮物胁迫对褐牙鲆GST酶及相关基因表达的影响
悬浮物对褐牙鲆幼鱼肌肉内GST酶活力的影响见图5,实验结果表明:悬浮物对A幼鱼在第24小时、B幼鱼在第24小时和第72小时肌肉内CAT酶活力呈现抑制趋势,10 000 mg/L实验组A幼鱼在第72小时和第96小时GST酶活力高于对照组(P<0.05),5 000 mg/L实验组A幼鱼在第48小时和第96小时GST酶活力与对照组无显著差异。
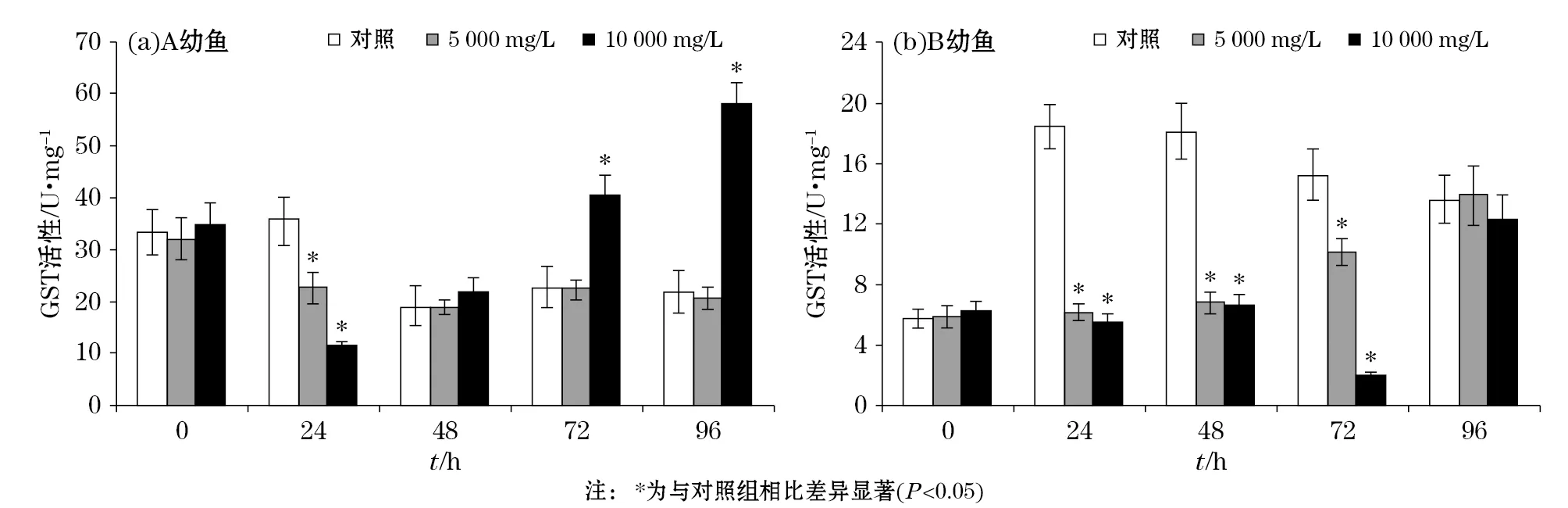
图5 褐牙鲆幼鱼体内GST活性随悬浮物胁迫时间的变化Fig.5 Changes of the GST activity in the body of P. olivaceus with the time of SS stress
悬浮物对褐牙鲆幼鱼肌肉内GST基因相对表达的影响见图6,在第24小时和第48小时悬浮物实验组对褐牙鲆A,B幼鱼GST基因相对表达诱导效果明显,实验组均高于对照组(P<0.05),并且GST基因相对表达在第24小时达到峰值(P<0.05);在第72小时和第96小时实验组与对照组差异不显著,实验组在第24小时和第48小时GST基因相对表达量明显高于第72小时和第96小时GST基因相对表达量;悬浮物对B幼鱼GST酶活力与相对基因表达量在第24小时和第48小时影响趋势相反。

图6 褐牙鲆幼鱼体内GST基因相对表达量随悬浮物胁迫时间的变化Fig.6 Changes of the relative gene expression of GST in the body of P. olivaceus with the time of SS stress
3 讨 论
在外界环境因子的胁迫下,生物体会产生一种会对细胞和基因结构造成损坏的活性氧分子,主要是超氧阴离子(O2-)、羟自由基(·OH)和H2O2[23]。低浓度的活性氧对生物体具有一定的积极作用[24],但是浓度过高的活性氧会对生物体产生损伤[25]。生物体内的抗氧化系统能清除过量的活性氧,保护体内各组织免受活性氧的损伤[26],抗氧化系统主要包括酶类抗氧化剂和非酶类抗氧化剂,目前针对生物体内的抗氧化系统主要集中于抗氧化系统酶活力方面的研究。
3.1 悬浮物对褐牙鲆总超氧化物歧化酶(T-SOD)活力及基因相对表达的影响
SOD是机体内唯一以O2为底物的酶,能催化超氧化物阴离子自由基(O·2-)发生歧化反应,及时清除生物体内的O·2-自由基,避免·OH的产生,在清除生物体内有害物质时发挥着重要的作用。已有研究表明,生物体的SOD酶活性与胁迫因子的强弱有关,大致表现为:轻度胁迫,SOD酶活力升高;重度胁迫,SOD酶活力下降[27-29]。本研究中5 000和10 000 mg/L试验组中A幼鱼在72和96 h时T-SOD酶活力变化趋势一致,原因可能是与褐牙鲆属于底栖鱼类、对悬浮物的耐受力较强有关,悬浮物浓度并未达到对褐牙鲆T-SOD酶活力产生重度胁迫的作用。在悬浮物胁迫下,B幼鱼T-SOD酶活力呈现先升高、再降低、再升高的趋势,并且在96 h达到峰值(P<0.05),推测原因是:在0~24 h,悬浮物胁迫可能刺激幼鱼体内产生较多的活性氧离子,为减少活性氧离子对机体的损害,鱼体内大量合成T-SOD酶,造成T-SOD酶活力升高,在24~72 h,鱼体短暂适应了悬浮物胁迫的环境,体内T-SOD酶合成减弱,当胁迫时间到达96 h,超过了幼鱼的耐受时间,又重新诱导了体内T-SOD酶的合成,直至T-SOD酶活力在96 h达到峰值,这与张雪等[14]对悬浮胁迫对菲律宾蛤仔对SOD影响的研究结果基本一致。
T-SOD酶活性及基因表达在受到悬浮物胁迫后,在开始的24 h或者48 h内存在一定增加的倾向,这与Stebbing[30]提出的“毒物兴奋效应”表现类似。但是胁迫时间过长,如B幼鱼T-SOD酶活力在48和72 h及SOD2的mRNA相对表达量在72和96 h实验组低于对照组,表明长时间悬浮物胁迫可能对褐牙鲆T-SOD酶活力及SOD2的mRNA 相对表达产生抑制作用。A幼鱼在悬浮物胁迫下,T-SOD酶活力与SOD2的mRNA 相对表达量在72和96 h诱导一致,这与A幼鱼T-SOD酶活力及SOD2的mRNA相对表达量变化趋势相反,原因推测是与不同体长规格的褐牙鲆对悬浮物耐受力不同,体长较小的A幼鱼为消除体内过多的活性氧分子,在72和96 h T-SOD酶活力较高,而体长较大的B幼鱼由于体内较强的悬浮物耐受能力,不需要诱导过多的T-SOD酶活力表达,便可以减少活性氧分子对机体的损害。
3.2 悬浮物对褐牙鲆过氧化氢酶(CAT)活力及基因相对表达的影响
生物体在胁迫因子的影响下,机体产生过多的H2O2,H2O2可产生活性极高的羟自由基(·OH),对生物体产生损伤[23],为减轻污染因子对机体伤害,生物体可调节抗氧化水平来清除机体中羟自由基和活性氧[31-33]。CAT是一种含铁的抗氧化酶,广泛存在于生物体各种组织[32],能清除体内过多H2O2,通过Haber-Weiss反应[31]阻断羟自由基(·OH)的产生,保持机体正常的生理功能。外源环境胁迫因子会造成细胞膜损伤,导致细胞膜透性增加,由于Haber-Weiss反应使CAT活性下降,而细胞内H2O2的水平不会产生大幅度变化[33]。
A幼鱼CAT酶活力及基因相对表达量变化趋势一致,都呈现先诱导、再抑制趋势,这表明CAT酶活力与基因相对表达存在较为明显的正相关关系。另外,悬浮物对CAT酶活力的影响可能存在一定的时间效应关系,所以在48 h以前CAT酶活力及A鱼CAT基因相对表达被诱导(P<0.05),在48~96 h时CAT酶活力及A鱼CAT基因相对表达被抑制(P<0.05),这与Stebbing[30]提出的“毒物兴奋效应”表现类似,也与蒋玫等[16]在底泥浸出液对脊尾白虾CAT酶活力及基因表达研究、杨国军等[15]在悬浮物对四角蛤蜊鳃CAT酶活力的研究结果一致。短时间胁迫会诱导褐牙鲆幼鱼体内的CAT酶活力,产生较多的CAT酶,来减轻悬浮物胁迫对机体的伤害,清除体内由于悬浮物胁迫产生的过多活性氧。当胁迫时间超过一定的限度,可能是褐牙鲆适应了胁迫环境,也可能是过长时间胁迫超过了褐牙鲆的耐受限度,造成了褐牙鲆CAT酶活力及基因相对表达量的下降。悬浮物胁迫下,两种规格的褐牙鲆CAT酶活力及基因表达相对量诱导或者抑制时间相一致,这表明,褐牙鲆肌肉内CAT的mRNA相对表达量与蛋白酶活性一致。
3.3 悬浮物对褐牙鲆谷胱甘肽转移酶(GST)活力及基因相对表达的影响
作为生物体内解毒酶系统的重要组成部分之一,GST参与生物机体的解毒代谢和抗氧化等过程,在生物体对各类来源的有毒物质的降解过程中均起着重要作用。GST的主要作用机制是催化谷胱甘肽(GSH)的巯基与一些内源性有毒物质(过氧化物)和外源性有毒物质(醌类化合物、α,β-不饱和羰基化合物)进行轭合反应,消除这些物质的毒性,保护机体内蛋白质免受有毒物质的损伤,从而达到维持机体正常生理环境的目的[34-35]。在本文研究中,悬浮物对A幼鱼24 h、B幼鱼24~72 h肌肉内CAT酶活力呈现抑制趋势,这与王凡等[5]的较高浓度Cu2+对牙鲆GSH-PX的活性产生抑制研究结果相一致,原因可能是当胁迫因子的浓度高于一定程度时,生物体内GST酶未能及时清除体内产生的有害物质,导致生物体细胞结构受到破坏,细胞中酶活性又逐渐下降[28]。A幼鱼在48 h以后GST酶活力趋势不明显,这可能与不同体长大小的鱼GST酶活力对悬浮物耐受力不同有关,一般来讲,鱼体越大,对外界胁迫因子的耐受力越强,B规格鱼体内GST酶活力下降,说明一部分GST酶可能进行降解因悬浮物胁迫机体内产生的有毒物质,保护体内一些蛋白质免受损伤,以达到保护机体维持正常的生命代谢活动。
褐牙鲆GST基因相对表达量在24~48 h内呈现明显的诱导效应,但在72~96 h与对照组无显著差异,表明在24~48 h悬浮物胁迫诱导了GSTmRNA的相对表达,但是此阶段内幼鱼GST酶活力并未呈现增加的趋势,褐牙鲆GST酶活力与mRNA相对表达量不一致,推测原因如下:第一、mRNA表达水平与蛋白水平不一定完全一致,可能有时间上的差异;第二、蛋白质检测方法的不同,在具体的酶活力测定实验中受到的干扰也有可能造成结果的差异;第三、mRNA的表达与蛋白的表达水平的线性关系只有0.4~0.5左右,影响蛋白质半衰期的因素(mRNA 转录后的调控、各种翻译后的修饰)较多,这些因素都可能造成蛋白质水平与mRNA表达水平的不一致[36]。
[1] WANG B, ZHANG C H, ZUO Y M, et al. The research introduction to biology and farming technologyof main species ofParalichthys[J].Marine Fisheries Research, 2004, 25(5): 86-72. 王波, 张朝辉, 左严明, 等. 牙鲆属主要经济鱼类的生物学及养殖研究概况[J].海洋水产研究,2004,25(5):86-72.
[2] YU W. Study on distribution variation of surface suspended solids in Bohai[D].Beijing: The University of Chinese Academy of Sciences,2011: 1-5. 于炜. 渤海表层悬浮物分布变异规律的研究[D].北京:中国科学院研究生院,2011: 1-5.
[3] ZHAO K H,translating. Chemical composition and material sources of marine suspended matter[J].Coastal Engineering,1999,15(3): 58-63. 赵奎寰译.海洋悬浮物的化学组成及物质来源[J].海岸工程,1999,15(3):58-63.
[4] LIU S, ZHOU Q X. Research advances in antioxidant enzymes for diagnosing environmental contamination[J].Chinese Journal of Ecology,2008,27(10):1791-1798.刘硕, 周启星.抗氧化酶诊断环境污染研究进展[J].生态学杂志,2008,27(10):1791-1798.
[5] WANG F, ZHAO Y F, LV J C, et al. Effect of Cu2+on CAT, SOD, GSH-PX of Paralichthys olivaceus[J].Journal of Huazhong Agricultural University,2007,26(6):836-838.王凡, 赵元凤, 吕景才,等.铜对牙鲆CAT、SOD和GSH-PX活性的影响[J].华中农业大学学报,2007, 26(6):836-838.
[6] WANG F, ZHAO Y F, LIU C F. Effect of Cu2+on the antioxidative defence in muscle ofParalichthysolivaceus[J].Freshwater Fisheries, 2007,37(2):27-29. 王凡, 赵元凤, 刘长发. Cu2+对牙鲆肌肉抗氧化防御系统的影响[J].淡水渔业,2007,37(2):27-29.
[7] HUANG W, CAO L, YE Z J, et al. Antioxidative responses and bioaccumulation in Japanese flounder larvae and juveniles under chronic mercury exposure[J].Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2010, 152(1): 99-106.
[8] CAO L, HUANG W, LIU J H, et al.Accumulation and oxidative stress biomarkers in Japanese flounder larvae and juveniles under chronic cadmium exposure[J].Comparative Biochemistry and Physiology Part C: Pharmacology, Toxicology and Endocrinology, 2010, 151(3): 386-392.
[9] WANG M L, ZHANG X M, HUANG G Q, et al. Effects of Ca2+and Mg2+concentrations on growth,SOD and CAT enzymatic activity of juvenileParalichthysolivaceus[J].Progress in Fishery Sciences, 2010, 31(3):29-36.王茂林, 张秀梅, 黄国强, 等. 不同钙、镁浓度对褐牙鲆幼鱼生长及SOD和CAT酶活力的影响[J].渔业科学进展, 2010, 31(3):29-36.
[10] LIU Y F. Study on the thermal tolerance of juvenileParalichthysolivaceusand the hybridsP.olivaceus♀×P.dentatus.♂[D].Qingdao:Ocean University of China,2014: 66-81.柳意樊. 褐牙鲆(Paralichthysolivaceus)与杂交鲆(褐牙鲆P.olivaceus♀×大西洋牙鲆P.dentatus♂)幼鱼高温耐受性研究[D].青岛:中国海洋大学, 2014:66-81.
[11] HUANG W. Toxic effects of mercury, lead and zinc on early life stages of flounder(Paralichthysolivaceus)[D].Beijing: University of Chinese Academy of Sciences,2010, 45-57.黄伟.汞、铅、锌对褐牙鲆(Paralichthysolivaceus)早期发育过程毒理作用的研究[D].北京:中国科学院研究生院, 2010: 45-57.
[12] LOU B,XU D D, XU H X, et al. Effect of high water temperature on growth, survival and antioxidant enzyme activities in the Japanese flounderParalichthysolivaceus[J].African Journal of Agricultural Research, 2011, 6(12): 2875-2882.
[13] XU D D, LOU B, ZHAN W, et al. Effect of high temperature stress on growth performance and activities of antioxidant enzymes in liver of olive flounderParalichthysolivaceus[J].Journ Aloffisheries of China, 2010, 34(7):1100-1106. 徐冬冬, 楼宝, 詹炜,等. 高温胁迫对褐牙鲆生长及肝脏抗氧化酶活性的影响[J].水产学报, 2010, 34(7):1100-1106.
[14] ZAHNG X, SONG L, YANG G J, et al. Effect of suspended solids on superoxide dismutase activity in Manila ClamRuditapesphilippinarum[J].FISHERIESS SCIENCE.2013, 32(6): 328-332.张雪, 宋伦, 杨国军,等. 悬浮物对菲律宾蛤仔超氧化物歧化酶活力的影响[J].水产科学, 2013, 32(6): 328-332.
[15] YANG G J, SONG L ,WANG N B, et al.Effects of suspended solid on the survival, growth, and antioxidant enzyme activities ofMactraveneriformis[J].Chinese Journal of Ecology, 2013, 32(8):2097-2103. 杨国军, 宋伦, 王年斌, 等. 悬浮物对四角蛤蜊生存和氧化酶活性的影响[J].生态学杂志, 2013, 32(8):2097-2103.
[16] JIANG M, LI L, SHEN X Q, et al.Effects of lixivium from ooze mud exposure on activity of antioxidant enzymesand related genes expression ofExopalaemoncarinicaudamysis larvae[J].Ecology and Environmental Sciences, 2013, 22(7):1182-1186. 蒋玫, 李磊, 沈新强, 等.底泥浸出液对脊尾白虾抗氧化解毒酶及相关基因表达的影响[J].生态环境学报, 2013, 22(7): 1182-1186.
[17] WANG G J, XIE J, YU D G, et al.Physiological responses of abaloneHaliotisdivericolorto suspended sediment stress[J].Journal of Dali an Fisheries University, 2007, 22(5):353-357. 王广军, 谢骏,余德光,等. 杂色鲍对底泥悬浮物胁迫的生理响应[J].大连水产学院学报, 2007, 22(5):353-357.
[18] Specification for marine monitorying: GB 17378-2007[S].Beijing:Standards Press of China, 2007.海洋监测规范: GB 17378-2007[S].北京:中国标准出版社,2007.
[19] Marine sediment quality: GB 18668-2002[S].Beijing:Standards Press of China,2002.海洋沉积物质量标准: GB 18668-2002[S].北京:中国标准出版社,2002.
[20] Water quality-determination of the acute toxicity of substance to a freshwater fish(BrachydaniorerioHamilion-Bachanan):GB/T 13267-91[S].Beijing:Standards Press of China,1992.水质·物质对淡水鱼(斑马鱼)急性毒性测定方法:GB/T 13267-91[S].北京:中国标准出版社,1992.
[21] WANG J Z, FAN M.Protein technical manual[M].Beijing:Science Press,2000: 42-47.汪家政, 范明. 蛋白质技术手册[M].北京: 科学出版社,2000: 42-47.
[22] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T))method[J].Methods,2001, 25(4):402-408.
[23] WEN C G, DAI G Y, XIE Y H, et al.Pb2+Effect on superoxide anion and scavenging hydroxyl free radicaland analysis soluble protein from anodonta woodioma[J].Journal of Nanchang University (Natural Science), 2009,33(4): 380-384.文春根, 代功园, 谢彦海, 等. 铅对背角无齿蚌抗超氧阴离子能力与抑制羟自由基的影响以及可溶性蛋白分析[J].南昌大学学报(理科版),2009,33(4): 380-384.
[24] ZHENG Z H, DONG S L, TIAN X L. Effects of cyclic pH with different durations on the growth ofLitopenaeusvannamei[J].Periodical of Ocean University of China,2008, 38(1):45-51.郑振华,董双林,田相利. pH不同处理时间的周期性变动对凡纳滨对虾生长的影响[J].中国海洋大学学报(自然科学版), 2008, 38(1):45-51.
[25] FRANCO R, SNCHE-OLEA R, REYES-REYES E M, et al. Environmental toxicity, oxidative stress and apoptosis: Ménageàtrois[J].Mutation Research/Fundamental & Molecular Mechanisms of Mutagenesis, 2009,674(2):3-22.
[26] CAO L. Toxic effects of cadmium and copper to early life stages of flounder (Paralichthysolivaceus)[D].Beijing:The University of Chinese Academy of Sciences, 2010:95-114. 曹亮. 铜、镉对褐牙鲆(Paralichthysolivaceus)早期发育阶段的毒理效应研究[D].北京:中国科学院研究生院, 2010:95-114.
[27] LIU C Q, LIU LI J, ZHANG Y J. et al. Effect of Ca2+/Mg2+in brine water from saltworks on growth and activities of APK and SOD in white legged shrimp[J].Fisheries Science, 2007, 26(2): 67-69. 刘存歧, 刘丽静, 张亚娟,等.基于卤水的养殖用水中Ca2+/Mg2+对凡纳滨对虾生长及体内SOD和AKP的影响[J].水产科学, 2007, 26(2): 67-69.
[28] LU S Q, LIU S J, LIU H Y. Effects of Cu2+on activities of protecting enzymes SOD,CAT and GSH-PX in liver tissue of Monopterus albus[J].Journal of Fishery Sciences of China,2002, 9(2): 138-141. 鲁双庆, 刘少军, 刘红玉. Cu2+对黄鳝肝脏保护酶 SOD,CAT,GSH-PX 活性的影响[J].中国水产科学,2002,9(2):138-141.
[29] YANG L H, FANG Z Q, ZHENG W B. Experiment with effect of cadmium on activity of superoxide dismutase in gill and liver tissue of crucian[J].Journal of Safety and Environment,2003, 3(3): 13-16. 杨丽华, 方展强, 郑文彪. 镉对鲫鱼鳃和肝脏超氧化物歧化酶活性的影响[J].安全与环境学报, 2003, 3(3): 13-16.
[30] STEBBING A R D. Hormesis the stimulation of growth by low levels of inhibition[J].Science of the Total Environment, 1982, 22(3):213-234.
[31] FENG T, ZHENG H Y, HONG W S, et al. Effect of benzo(a)pyrene on antioxidant enzyme activities inBoleophthalmuspectinirostrisliver[J].Chinese Journal of Applied Ecology, 2001,12(3):442-424. 冯涛,郑徽云,洪万树,等.苯并(a)花对大弹涂鱼肝脏抗氧化酶活性影响的初步研究[J].应用生态学报,2001, 12(3):442-424.
[32] FANG Y Z, LI W J. Basic theory of free radical and enzyme and its application in biology and medicine[M].Beijing: Science Press,1989:129-146. 方允中, 李文杰. 自由基与酶基础理论及其在生物学和医学上的应用[M].北京: 科学出版社, 1989: 129-146.
[33] SHEN A L, MA S Q, JIA Q H. Research on the physiological responses ofSparusmacrocephalusstressedby Cu2+and Cd2+[J].Marine Fisheries,2007, 29(3):23-29. 沈盎绿, 马胜伟, 贾秋红. 黑鲷受铜、镉胁迫的生理反应[J].海洋渔业,2007, 29(3):23-29.
[34] MASELLA R, DI Benedetto R D, VARI R, et al. Novel mechanisms of natural antioxidant compounds in biological systems: involvement of glutathione and glutathione-related enzymes[J].The Journal of Nutritional Biochemistry, 2005, 16(10): 577-586.
[35] SHIMADA T. Xenobiotic-metabolizing enzymes involved in activation and detoxification of carcinogenic polycyclic aromatic hydrocarbons[J].Drug Metabolism and Pharmacokinetics, 2006, 21(4):257-276.
[36] MATHEWS M B, SONENBERG N, HERSHEY J W B. Translational control in biology and medicine[M].New York:Cold Spring Harbor Laboratory Press, 2007: 534-546.
Received: December 10, 2015
Effects of Suspended Substances on Antioxidant Enzyme Activities in Muscle ofParalichthysolivaceusand Related Gene Expressions
XIAO Guang-xia1, XU Wen-yuan2, JIA Lei1, ZHENG De-bin1, ZHANG Bo1, MA Chao1
(1.BohaiFisheriesResearchInstituteofTianjin, Tianjin 300457, China; 2.LinyiEntry-ExitInspectingandQuarantineBureau, Linyi 276034, China)
In order to study the tolerance mechanism of juvenile ofParalichthysolivaceusisunder the stress of suspended substances (SS), two sizes ofP.olivaceusare chosen as the experimental material and the activities of total superoxide dismutase (T-SOD), catalase (CAT), glutathions S-transferase (GST) in the muscle ofP.olivaceusand the changes in mRNA transcription levels of SOD2, CAT and GST are studied under the conditions of suspended substance concentrations of 5 000 mg/L and 10 000 mg/L. The experimental results show that the enzyme activity of T-SOD has a certain changing tendency: in the B size of juvenile the enzyme activity of T-SOD shows a trend of first increased-then decreased and then increased again and the relative gene expression of T-SOD reaches to the peak value (P<0.05) in 48 h. The relative gene expressions of T-SOD in the two sizes of juveniles show an opposite changing tendency in 72 h and 96 h. The enzyme activity of CAT tends to increase first and then to decrease and the relative gene expressions of CAT in the A size of juvenile and the B size of juvenile tested under the condition of 5 000 mg/L are consistent in their changing tendency with the CAT enzyme activity. The enzyme activity of GST in the B size of juvenile is lower in the experimental groups than that in the control group (P<0.05). The relative gene expression of GST is higher in the 24 h and 48 h experimental groups than that in the control group (P<0.05), but shows no significant difference in the 72 h and 96 h experimental groups and the control group. All these results indicate that the SS has a certain effect on the antioxidant enzyme activities and gene expressions ofP.olivaceusand different size ofP.olivaceusvaries in the antioxidant enzyme activity and gene expression. The enzyme activities of T-SOD and GST differ in their changing trends from the relative gene expression, whereas the changing trend of the CAT enzyme activity is consistent with that of the relative gene expression. This study may provide basic data not only for studying the tolerance mechanism ofParalichthysolivaceusisunder the SS stress, but also for breeding theP.olivaceusspecies which can be tolerant of suspended substances.
Paralichthysolivaceus; suspended substances(SS); antioxidant system enzyme; gene expression
2015-12-10
天津市水产局青年科技创新项目——悬浮物胁迫对褐牙鲆存活、抗氧化系统酶活力及基因表达的研究(J2014-02);天津市农委农业科技合作项目——海水新品种的引进及示范(201304070);国家鲆鲽类产业技术体系项目——天津综合试验站建设(CARS-05-z01);天津市科技计划项目——海珍品高效养殖技术集成与应用(15zxbfnc00100);国家农业科技成果转化项目——鲆鲽类高效生态养殖技术的集成与示范(2014GB2A100531)
肖广侠(1984-),男,工程师,硕士,主要从事水产动物遗传育种及增殖放流评估方面研究. E-mail:gxxiao@163.com
*通讯作者:贾 磊(1983-),男,高级工程师,硕士,主要从事苗种培育及生态养殖方面研究. E-mail: tianjinbohaisuo@163.com
Q945.78
A
1671-6647(2016)04-0542-11
10.3969/j.issn.1671-6647.2016.04.010
