拉萨河谷植物物种丰富度空间分布格局及其环境解释
2016-11-12拉多张燕杰刘杰崔玲玲庞有智
拉多,张燕杰,刘杰,崔玲玲,庞有智
(1.西藏大学理学院,西藏 拉萨 850000;2.内江师范学院地理与资源科学学院,四川 内江 641100)
拉萨河谷植物物种丰富度空间分布格局及其环境解释
拉多1*,张燕杰1,刘杰1,崔玲玲1,庞有智2
(1.西藏大学理学院,西藏 拉萨 850000;2.内江师范学院地理与资源科学学院,四川 内江 641100)
为研究拉萨河谷植物物种丰富度的空间分布格局,按不同植被类型设置47个样地,每个样地随机设置3个样方,共141个样方。记录维管植物247种,隶属47科、134属。运用基于距离的Moran特征向量图(MEM)和方差分解, 分析环境因子和空间变量对植物物种丰富度空间分布的影响;运用广义可加模型(GAM)对各环境因子与转换物种丰富度(TSR)进行回归分析,同时对各环境因子之间进行相关分析;运用除趋势对应分析(DCA)对样地和物种进行非约束排序,并将环境因子与排序轴之间进行相关分析。结果表明空间结构对拉萨河谷植物物种丰富度分布具有重要作用,而环境因子的空间格局是影响物种丰富度空间分布格局的重要因素;各环境因子与TSR的GAM拟合结果发现气候因子、经纬度和海拔对TSR存在显著的影响;DCA排序结果表明干扰可能是影响物种丰富度空间格局的未测环境因子,DCA第二轴反映了湿度梯度。干扰和湿度可能是拉萨河谷植物物种丰富度空间分布格局的主要影响因素。
物种丰富度;Moran特征向量图;干扰;气候因子;拉萨河谷
生物多样性(biodiversity)对于维持生态系统稳定至关重要[1-2]。物种多样性(species diversity)则是生物多样性的基本内容,是最简单有效的生物多样性测度。因此物种多样性的分布格局及其影响因素是生态学领域研究的核心问题之一[3-4],而物种多样性的空间分布格局研究则多以物种丰富度(species richness)作为测度指标[3]。不同空间尺度和环境梯度上的相关研究发现的物种丰富度空间分布格局也不尽相同[5-9]。影响物种丰富度分布格局的主要环境因素包括海拔、纬度、经度、气候、地形、土壤等。由于海拔和纬度梯度包含了水分、能量和光照等重要环境因子的变化而成为物种丰富度梯度格局研究的重要方面;气候被认为是影响物种丰富度大尺度格局的主要因素[10],而地形和土壤因子的作用则是物种丰富度小尺度格局研究的重要内容[11-12]。另外,植物群落的空间结构也可用于探讨物种多样性空间分布格局[12]。基于距离的Moran特征向量图(Moran’s eigenvector maps, MEM),之前被称为邻体矩阵主坐标分析(principal coordinates of neighbor matrices, PCNM)[13-15],近年来逐渐被引入到物种多样性空间格局的研究中[12,16-18],而这一方法在青藏高原地区的相关研究中则鲜有采用。
青藏高原独特的地理位置和自然条件使这一地区物种多样性的相关研究备受关注。杨元合等[11]、王长庭等[19]、段敏杰等[20]、武建双等[21]、拉琼等[8]、罗黎明等[22]、刘哲等[23]均对青藏高原不同区域的物种丰富度分布格局进行了相关研究,然而拉萨河谷整体植物物种丰富度空间分布格局的相关研究则鲜有报道。虽然拉萨河谷经纬度跨度不大,但由于地理位置特殊、地形复杂,由东到西、由南到北气候差异比较明显,植被类型的分布也比较复杂,拉萨河谷平原也是整个西藏人类活动最强的区域。因此本研究以拉萨河谷作为研究区域,探讨拉萨河谷植物物种丰富度空间分布格局的影响因素,有助于更全面地了解该区域植物物种丰富度空间分布格局的成因,为该区域的生物多样性及生态系统保护提供科学依据。
1 材料与方法
1.1研究区域
拉萨河谷位于西藏自治区中南部,是雅鲁藏布江最大的支流拉萨河流域河谷,位于东经90°05′-93°20′和北纬29°20′-31°15′之间,流域面积约32471 km2[24]。气候以温暖半干旱为主,由区域内和附近4个气象监测站点的平均数据可知,拉萨河谷内年均气温5.9 ℃,年降雨量440 mm。降雨量的90%集中在5月至9月,多为夜雨,蒸发量大。拉萨河谷地处雅鲁藏布江中游谷地亚高山灌丛草原区[25],是藏东南部湿润的亚热带森林、灌丛、草甸与藏西北部寒冷干旱的高山草甸,高寒草原、荒漠之间的过渡带,以高山草甸、高寒草原和亚高山灌丛草原为主,而河谷底部主要以荒漠植被占优势。
1.2样方调查
于2014年7月和2015年7月对研究区域植被进行样方调查,按植被类型不同共设置47个样地(图1),每个样地随机设置3个样方,林地与灌丛样方面积为5 m×5 m,稀疏灌丛区样方面积为2 m×2 m,草原及草甸区样方面积为1 m×1 m,共设置样方141个。对样方进行GPS定位,记录其经纬度和海拔,并记录植物物种组成、覆盖度等信息。每个样地取3个样方中间位置的GPS定位信息作为样地的位置信息、物种总数为样地的物种数、平均覆盖度为样地的覆盖度。大部分植物在野外鉴定到种水平,未能鉴定的植物采集标本并且拍照,记录其所在样方编号,带回实验室进一步鉴定。由于没有花或果实,或者因花或果实不完整而无法鉴定到种水平的,以“属名 sp.”或“科名+数字”记录,如紫菀属一种记为“Astersp.”、禾本科一种记为“Gramineae 1”。
图1 拉萨河谷47个样地分布图Fig.1 Distribution of 47 plots in Lhasa Valley
1.3环境因子
协同克里金(Cokriging)插值法可以引入海拔等地形因子作为协同变量,对降雨量和气温都可以取得较好的插值效果[26-27]。以本研究区域内及周围16个气象监测站平均30年(最少17年,最多40年)的气候数据为基础,以海拔作为协同变量,运用ArcGIS10.1进行空间插值计算,得到47个样地的年降水量(AP)和年平均气温(MAT),并计算实际蒸散量(AET)、潜在蒸散量(PET)和湿润指数(MI),计算过程请见参考文献[6,28-29]。湿润指数的数值越大表示生境越干燥[6]。
1.4数据分析
由于不同植被类型设置了不同面积的样方,因此每个样地3个样方的总面积不同,采用转换物种丰富度(transformed species richness, TSR)[30]作为各样地的物种丰富度测度可用于消除面积对物种丰富度的影响[31-32],其计算公式为:
TSR=N/lgS
其中,N为样地中植物的种数,S为取样面积。
基于每个样地的空间坐标,运用基于距离的Moran’s特征向量图(MEM)构建空间变量,运用双终止准则(two-step procedure)[28]对空间变量和环境因子进行前向选择(forward selection)后与物种丰富度数据进行冗余分析(RDA)和方差分解(variation partitioning),来评估空间变量与环境因子对物种丰富度的解释程度。MEM的详细步骤请见参考文献[13],MEM和方差分解运用R3.2.2的PCNM软件包完成。
采用广义可加模型(generalized additive model,GAM)对单个环境因子与物种丰富度进行回归分析。用“可解释偏差(deviance explained,de)”来评估模型的拟合效果[29]。环境因子之间可能存在的较强相关性会影响回归结果的分析,因此对各环境因子进行相关分析,用以结合回归分析来探讨环境因子对物种丰富度的影响。GAM和相关分析运用R3.2.2的mgcv软件包完成。
对样地和物种数据进行非约束排序(direct ordination)可用于探讨样地和物种的分布格局所反映出的环境梯度,在非约束排序基础上将环境因子变量与排序轴进行相关分析,可以探讨已测环境因子与物种分布的关系[14]。因此本研究采用DCA排序分析,再将环境因子与排序轴进行相关分析,探讨影响物种分布格局的环境因子。DCA和相关分析运用R3.2.2的vegan软件包完成。
2 结果与分析
2.1物种组成
本研究47个样地141个样方共记录维管植物47科,134属,247种(植物的分类鉴定以《Flora of China》为依据)。其中出现频度最高的科为菊科(308次),其次为莎草科、豆科、蔷薇科和禾本科;出现频度最高的属为嵩草属,其次为委陵菜属和蒿属;频度最高的种为高山及亚高山草甸的主要优势种矮生嵩草(Kobresiahumilis),其次为高山嵩草(Kobresiapygmaea),另外,蔷薇科的钉柱委陵菜(Potentillasaundersiana)、豆科的笔直黄耆(Astragalusstrictus)、菊科的毛莲蒿(Artemisiavestita)、白花丹科的小蓝雪花(Ceratostigmaminus)、紫草科的毛果草(Lasiocaryumdensiflorum)、玄参科的肉果草(Lanceatabetic)等也均以较高的频度出现。
47个样地中,转换物种丰富度(TSR)最高为52.4,分布在海拔4239 m的嵩草草甸,以矮生嵩草和高山嵩草为优势种,伴生种多且覆盖度较均匀,并有狼毒(Stellerachamaejasme)、笔直黄耆等干扰指示种出现;TSR最低为2.1,分布在海拔3604 m的砂生槐灌丛。以灌木砂生槐(Sophoramoorcroftiana)为优势种,且其冠盖度约为50%,伴生种很少,仅见藏沙蒿(Artemisiawellby)、菊叶香藜(Dysphaniaschraderiana)等。
2.2MEM和方差分解

图2 方差分解图Fig.2 Variation partitioning results图中数字为校正R2。the Arabic numbers are adjusted R-squares.
将47个样地的地理坐标转换为笛卡尔坐标后,运用MEM和前向选择得到3个显著(P<0.001)的特征向量PCNM2、PCNM3和PCNM6作为空间变量。将环境因子进行前向选择后与空间变量一起进行冗余分析(RDA)和方差分解,方差分解结果如图2所示。环境因子与空间变量对物种分异的总解释量为44.3%(a+b+c),其中环境因子的总解释量为21.9%(a+c),空间变量的总解释量为37.6%(b+c)。环境因子单独解释的部分为6.7%(a),而环境因子与空间变量共同解释部分为15.2%(c),环境因子与空间变量共同的解释部分即诱导性空间依赖,即环境因子的空间结构引起物种丰富度差异[14]。空间变量单独解释部分为22.4%(b),这部分与已测环境因子无关,说明空间结构对物种丰富度存在重要影响。
2.3环境因子及覆盖度对物种丰富度的影响
环境因子及覆盖度与转换物种丰富度(TSR)的GAM拟合结果显示(图3),除覆盖度对TSR的影响不显著(P>0.05)外,环境因子对TSR均有较显著的影响(P<0.05)。
TSR随海拔升高的变化较为复杂,控制其自由度(edf<3)时,拟合线的可解释偏差为14.5%,呈现先升高后降低后再稍有升高的格局。海拔4000 m左右时最高,3600 m左右时最低。TSR随纬度升高逐渐升高后趋于平缓,随经度升高呈现先升高后降低的单峰格局。
年降水量和实际蒸散量与TSR的GAM拟合模型呈较对称的单峰型,即年降水量和实际蒸散量最低和最高时,TSR最低,处于中间值时TSR最高。TSR随年平均气温和潜在蒸散量升高,呈现先升高后降低的单峰格局,即年平均气温和潜在蒸散量最大时,TSR最低。TSR随湿润指数的变化较为复杂,控制自由度(edf<3)后,拟合线较为平滑,TSR随湿润指数增大先降低后升高,之后在湿润指数最高时TSR降至最低,即环境条件最干燥时TSR最低(表1)。

表1 环境因子及覆盖度之间的相关系数(r)矩阵
AP: Annual precipitation; MAT: Mean annual temperature; AET: Actual evapotranspiration; PET: Potential evapotranspiration; MI: Moisture index.**表示P<0.01;*表示P<0.05。**represents correlation is significant at the 0.01 level, * represents correlation is significant at the 0.05 level.
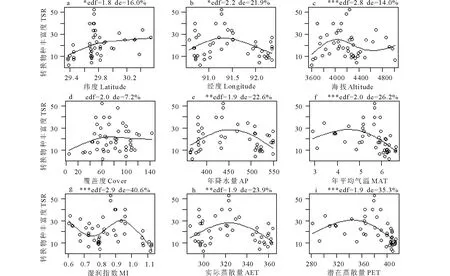
图3 环境因子及覆盖度与转换物种丰富度(TSR)的GAM拟合图Fig.3 Generalized additive model of each environmental factors and TSR AP: Annual precipitation; MAT: Mean annual temperature; AET: Actual evapotranspiration; PET: Potential evapotranspiration; MI:Moisture index. edf代表估计自由度,de代表可解释偏差。“edf” is estimated degree of freedom, “de” is deviance.*代表P<0.05,**代表P<0.01,***代表P<0.001. * represents P<0.05, **represents P<0.01, ***represents P<0.001.
2.4样地和物种的DCA排序及环境解释
对47个样地131个物种的覆盖度矩阵(已剔除频度≤5且覆盖度≤5%的偶见种)进行DCA排序。DCA排序第一轴的解释量为10%(特征值为0.66),第二轴的解释量为5%(特征值为0.36)。在环境因子相关分析的基础上,选择6个环境因子与排序轴进行相关分析(图5B)。值得关注的是,样地沿两个轴的排列基本上能够对应样地的地理位置(少数几个样地除外,如样地35、36)(图4,图5A,表2),即样地沿第一轴的排列基本上对应了地理位置上由西到东的分布,沿第二轴的排列基本上对应了地理位置由南到北的分布,表明拉萨河谷植物物种分布与其所处地理位置关系密切。
物种排序图显示,第一轴左侧靠近排序轴的位置排列了狼毒、笔直黄耆等草场退化及干扰指示种,以及绢毛蔷薇(Rosasericea)、川藏香茶菜(Isodonpharicus)等灌木,右端则主要排列了三脉梅花草(Parnassiatrinervis)、球花马先蒿(Pedicularisglobifera)、云生毛茛(Ranunculusnephelogenes)、独一味(Lamiophlomisrotate)等生长在较茂盛草甸的物种,可能反映了干扰梯度。样地排序图显示,第一轴从左到右基本排列了研究区域从西到东的样地顺序,可能反映了经度梯度。加入环境因子后显示,经度和海拔与第一轴显著正相关(r=0.898和0.790,P<0.001),表明DCA排序第一轴可能反映了干扰、经度和海拔的共同梯度。

图5 样地和物种DCA排序图Fig.5 DCA ordination of plots and species A为样地排序图, 圆圈由大到小代表每个样地的转换物种丰富度(TSR)由高到低;B为物种排序图,物种编号对应的物种信息详见表2,为使图更清晰,一些物种未在图中显示,以灰色空心点表示。 A: DCA ordination of plots, size of the circles represents difference of TSR. B: DCA ordinations of species, species information please refer to Table 2, for clarity, some species were indicated with gray dots.
样地排序图显示,第二轴上端排列了大果圆柏(Juniperustibetica)群落以及川西锦鸡儿(Caraganaerinacea)、毛叶绣线菊(Spiraeamollifolia)和小叶栒子(Cotoneastermicrophyllus)等较喜湿的多种灌木混生的灌丛群落,而生长在干旱生境的砂生槐群落和毛莲蒿群落等则排列在第二轴下端。加入环境因子后发现,第二轴与年降水量和纬度显著正相关(r=0.832和0.997,P<0.001),与年平均气温和湿润指数显著负相关(r=-0.947和-0.986,P<0.001),即第二轴可能反映了水分和能量共同作用的湿度梯度。

表2 DCA排序图中显示的主要物种表
3 讨论
DCA样地排序结果与样地的地理位置基本对应(图4,图5A),表明拉萨河谷植物物种分布与其所处地理位置存在较密切的关系,也可以解释MEM基于样地的地理位置构建的空间变量总解释量很高(37.6%)的原因,而GAM回归也发现经度和纬度对TSR分布均有较显著的影响(图3),由此可知拉萨河谷物种丰富度的空间分布格局与地理位置有关。
方差分解发现环境因子与空间变量共同解释量很高(15.2%),表明已测环境因子的空间结构是影响物种丰富度空间分布的重要原因。海拔与DCA第一轴显著相关(r=0.790,P<0.001,图5),并且对TSR的分布存在较显著的影响(图3)。海拔梯度实际上是环境水分和能量的综合反映,水分和能量都对物种的分布产生影响,而且在不同的环境中两者对物种丰富度的影响程度可能不同[7]。以往青藏高原的相关研究中,物种丰富度沿海拔梯度的分布格局不尽相同[8,11,19-23]。本研究区域内海拔跨度较大(约1400 m),低海拔区域处于研究区域西南部干暖河谷,水分的限制作用使植被主要为禾草草原或荒漠,TSR最低;随着海拔升高,水分条件好转,植被发展为草甸和灌丛草甸,物种组成逐渐丰富,TSR逐渐升高;然而随着海拔再升高,气温降低,能量的限制作用开始凸显,TSR降低;海拔继续升高至4700 m左右时植被发展为灌丛到草甸的过渡带,TSR再次稍上升,当然也不排除因海拔4700 m以上样地较少而影响GAM拟合模型的可能。总体上在研究区域的中海拔区域TSR最高,与段敏杰等[20]、罗黎明等[22]和刘哲等[23]在西藏地区的研究结果相似。
在青藏高原,一般情况下降雨量对植物物种丰富度存在显著影响,随着降雨量增加物种丰富度呈升高趋势[8,11,21],然而本研究中年降雨量和实际蒸散量与TSR的GAM拟合模型均呈单峰格局(图3)。以往的一些研究中年平均气温和潜在蒸散量与物种丰富度之间普遍存在单峰曲线关系[9],新疆地区的相关研究也发现相似的单峰格局[7],本研究也发现TSR与年平均气温和潜在蒸散量的GAM拟合模型呈单峰格局(图3)。拉萨河谷从东北到西南气温逐渐上升,降雨量逐渐减少,经度与年降雨量显著相关(表1)。东北部属于年平均气温和潜在蒸散量较低的区域、年降雨量和实际蒸散量较大,能量的限制使TSR较低;从东北到西南随着年平均气温和潜在蒸散量增加、年降雨量和实际蒸散量减少,TSR开始受到水分限制,能量增加会加剧水分亏缺,从而使TSR随能量增加而降低。另外,TSR这种格局也可能反映的是水分、能量、经纬度和干扰的共同作用。
方差分解还发现空间变量单独解释量很高(22.4%),表明空间结构对物种丰富度的分布格局也发挥着重要作用,这种空间结构可能来自物种分布的空间自相关性,也可能有未测的环境因子或历史生态事件的影响。物种丰富度的空间分布格局呈现一定的聚集性,距离近的样地之间TSR也相似(图4)。野外调查过程中也发现拉萨河谷植被存在斑块状分布的特征,这种斑块状分布的空间结构可能来源于扩散限制,区域内植物群落的草本优势种多为多年生根茎草本(如嵩草属),克隆繁殖较为普遍;灌木和乔木优势种的种子存活率也容易受到环境限制(如大果圆柏、砂生槐),可能是物种丰富度空间结构的一个成因。另外,干扰也可能使生境趋于破碎化,进而影响物种分布,也可能是物种丰富度具有空间结构的另一个原因。本研究区域中部样地处于拉萨市周围,人类活动可能使其受到一定程度的干扰,中度干扰假说认为中等强度的干扰可以使物种丰富度达到最高[34],在藏北草原也有相关研究认为中等强度的放牧干扰可以使高寒草地具有较高的物种丰富度[20],因此,干扰也可能是研究区域中部TSR较高的一个原因。
植被覆盖度可以用于表征生产力来探讨生产力与物种丰富度的关系[35]。西藏其他地区的相关研究中发现了物种丰富度随覆盖度增加而增加的格局[8],然而,本研究中覆盖度对物种丰富度的影响并不显著(图3)。覆盖度在50%到100%之间时,TSR在10到40之间均有分布。虽然植被覆盖度可以认为是环境因子综合作用的外在表现,然而如果优势种的分盖度占绝对优势,覆盖度高不一定代表生境质量好,竞争作用的影响可能使物种丰富度更低。
4 结论
运用Moran’s特征向量图(MEM)、方差分解、回归分析和DCA排序方法探讨了拉萨河谷植物物种丰富度空间分布格局及其与环境因子的关系,拉萨河谷植物物种丰富度的空间分布格局受经纬度、海拔、气候因子和未测环境因子干扰的共同影响,空间结构对物种丰富度的分布格局也发挥着重要作用。干扰和湿度可能是影响拉萨河谷植物物种丰富度空间分布格局的主要因素。今后可以进一步将干扰进行半定量化,并引入土壤等因素对拉萨河谷植物物种丰富度格局进行研究,更全面地揭示拉萨河谷植物物种丰富度分布格局及其与环境的关系。
致谢:感谢许敏在植物分类鉴定工作中给予的帮助。
[1]Naeem S, Li S. Biodiversity enhances ecosystem reliability. Nature, 1997, 390: 507-509.
[2]Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade long grassland experiment. Nature, 2006, 441: 629-632.
[3]Gaston K J. Global patterns in biodiversity. Nature, 2000, 405: 220-227.
[4]Chi X L, Tang Z Y, Fang J Y. Patterns of phylogenetic beta diversity in China’s grasslands in relation to geographical and environmental distance. Basic and Applied Ecology, 2014, 15: 416-425.
[5]Qian H, Ricklefs R E.A latitudinal gradient in large-scale beta diversity for vascular plants in North America. Ecology Letters, 2007, 10: 737-744.
[6]LiuY, Zhang Y P, He D M,etal. Elevational patterns of species richness and climate interpretation of Longitudinal Range-Gorge Region. Chinese Science Bulletin, 2007, 52(II): 43-50.
[7]Li L P, Nurbay A, Wang S P,etal. Distribution patterns and climatic explanations of species richness of vascular plants in Xinjiang, China. Arid Zone Research, 2011, 28(1): 25-30.
[8]La Q, Zhaxi C, Zhu W D,etal. Plant species-richness and association with environmental factors in the riparian zone of the Yarlung Zangbo River of Tibet, China. Biodiversity Science, 2014, 22(3): 337-347.
[9]Liu Q F, Liu Y, Sun X L,etal. The explanation of climatic hypotheses to community species diversity patterns in Inner Mongolia grasslands. Biodiversity Science, 2015, 23(4): 463-470.
[10]Francis A P, Currie D J.A globally consistent richness-climate relationship for angiosperms. The American Naturalist, 2003, 161(4): 523-536.
[11]Yang Y H, Rao S, Hu H F,etal. Plant species richness of alpine grasslands in relation to environmental factors and biomass on the Tibetan Plateau. Biodiversity Science, 2004, 12(1): 200-205.
[12]Zhao H B, Liu T, Lei J Q,etal. β diversity characteristic of vegetation community on south part of Gurbantunggut Desert and its interpretation. Acta Prataculturae Sinica, 2010, 19(3): 29-37.
[13]Dray S, Legendre P, Peres-Neto P R. Spatial modelling: A comprehensive framework for principal coordinate analysis of neighbour matrices (PCNM). Ecology Model, 2006, 196: 483-493.
[14]Borcard D, Gillet F, Legendre P. Numerical Ecology with R[M]. New York: Springer Science+Business Media, 2011.
[15]Gao M X, He P, Sun X,etal. Relative contributions of environmental filtering, biotic interactions and dispersal limitation in a soil collembolan community from a temperate deciduous forest in the Maoer mountains. Chinese Science Bulletin, 2014, 59: 2426-2438.
[16]Zhao A J, Hu T X, Chen X H. Multiple-scale spatial analysis of community structure in a mountainous mixed evergreen-deciduous broad-leaved forest, southwest China. Biodiversity Science, 2009, 17(1): 43-50.
[17]Yuan X, Ma K M, Wang D. Partitioning the effects of environmental and spatial heterogeneity on distribution of plant diversity in the Yellow River Estuary. Science China, 2012, 55(6): 542-550.
[18]Cai B, Huang Y, Chen Y Y,etal. Geographic patterns and ecological factors correlates of snake species richness in China. Zoological Research, 2012, 33(4): 343-353.
[19]Wang C T, Wang Q J, Long R J,etal.Changes in plant species diversity and productivity along an elevation gradient in an alpine meadow. Acta Phytoecologica Sinica, 2004, 28(2): 240-245.
[20]Duan M J, Gao Q Z, Guo Y Q,etal. Species diversity distribution pattern of alpine grassland communities along an altitudinal gradient in the Northern Tibet. Pratacultural Science, 2011, 28(10): 1845-1850.
[21]Wu J S, Li X J, Shen Z,etal. Species diversity distribution pattern of alpine grasslands communities along a precipitation gradient across Northern Tibetan Plateau. Acta Prataculturae Sinica, 2012, 21(3): 17-25.
[22]Luo L M, Miao Y J, Wu J S,etal. Variation in the biodiversity of montane shrub grassland communities along an altitudinal gradient in a Lhasa River basin valley. Acta Prataculturae Sinica, 2014, 23(6): 320-326.
[23]Liu Z, Li Q, Chen D D,etal. Patterns of plant species diversity along an altitudinal gradient and its effect on above-ground biomass in alpine meadows in Qinghai-Tibet Plateau. Biodiversity Science, 2015, 23(4): 451-462.
[24]Comprehensive Scientific Expedition Team of Chinese Academy of Sciences in Qinghai-Tibet Plateau. Tibetan Rivers and Lakes[M]. Beijing: Science Press, 1984.
[25]Comprehensive Scientific Expedition Team of Chinese Academy of Sciences in Qinghai-Tibet Plateau. Tibetan Vegetation[M]. Beijing: Science Press, 1988.[26]Goovaerts P.Geostatistical approaches for incorporating elevation into the spatial interpolation of rainfall. Journal of Hydrology, 2000, 228: 113-129.
[27]He H Y, Guo Z H, Xiao W F. Review on spatial interpolation techniques of rainfall. Chinese Journal of Ecology, 2005, 24(10): 1187-1191.
[28]Blanchet F G, Legendre P, Borcard D. Forward selection of explanatory variables. Ecology, 2008, 89(9): 2623-32.
[29]Liu X Z, Wang G A, Li J Z,etal. Relationship between temperature and δ13C values of C3herbaceous plants and its implications of WUE in farming-pastoral zone in North China. Acta Ecologica Sinica, 2011, 31(1): 123-136.
[30]He F, Legendre P, Lafrankie J V. Spatial pattern of diversity in a tropical rain forest in Malaysia. Journal of Biogeography, 1996, 23: 57-74.
[31]Qian H. Large scale biogeographic patterns of vascular plant richness in North America: An analysis at the genera level. Journal of Biogeography, 1998, 25: 829-836.
[32]Wang Z H, Chen A P, Piao S L,etal. Pattern of species richness along an altitudinal gradient on Gaoligong Mountains, Southwest China. Biodiversity Science, 2004, 12(1): 82-88.
[33]Wang S P, Zhang Z Q, Zhang J J,etal. Modeling the effects of topography on the spatial distribution of secondary species in a small watershed in the Gully region on the Loess Plateau, China. Acta Ecologica Sinica, 2010, 30(22): 6102-6112.
[34]Petraitis P S, Latham R E, Niesenbaum R A. The maintenance of species diversity by disturbance. The Quarterly Review of Biology, 1989, 64(4): 393-418.
[35]La Q, Grytnes J A, Birks H J B. Alpine vegetation and species richness patterns along two altitudinal gradients in the Gyama Valley, south-central Tibet, China. Plant Ecology and Diversity, 2010, 3: 235-247.
[6]刘洋, 张一平, 何大明, 等. 纵向岭谷区山地植物物种丰富度垂直分布格局及气候解释. 科学通报, 2007, 52(II): 43-50.
[7]李利平, 努尔巴依·阿布都沙力克, 王少鹏, 等. 新疆野生维管束植物物种丰富度分布格局的水热解释. 干旱区研究, 2011, 28(1): 25-30.
[8]拉琼, 扎西次仁, 朱卫东, 等. 雅鲁藏布江河岸植物物种丰富度分布格局及其环境解释. 生物多样性, 2014, 22(3): 337-347.
[9]刘庆福, 刘洋, 孙小丽, 等. 气候假说对内蒙古草原群落物种多样性格局的解释, 生物多样性, 2015, 23(4): 463-470.
[11]杨元合, 饶胜, 胡会峰, 等. 青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系. 生物多样性, 2004, 12(1): 200-205.
[12]赵怀宝, 刘彤, 雷加强, 等. 古尔班通古特沙漠南部植物群落β多样性及其解释. 草业学报, 2010, 19(3): 29-37.
[15]高梅香, 何萍, 孙新, 等. 环境筛选、扩散限制和生物间相互作用在温带落叶阔叶林土壤跳虫群落构建中的作用. 科学通报, 2014, 59: 2426-2438.
[16]赵安玖, 胡庭兴, 陈小红. 西南山地阔叶混交林群落空间结构的多尺度特征. 生物多样性, 2009, 17(1): 43-50.
[18]蔡波, 黄勇, 陈跃英, 等. 中国蛇类物种丰富度地理格局及其与生态因子的关系. 动物学研究, 2012, 33(4): 343-353.
[19]王长庭, 王启基, 龙瑞军, 等. 高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究. 植物生态学报, 2004, 28(2): 240-245.
[20]段敏杰, 高清竹, 郭亚奇, 等. 藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局. 草业科学, 2011, 28(10): 1845-1850.
[21]武建双, 李晓佳, 沈振, 等. 西藏北高寒草地样带物种多样性沿降水梯度的分布格局. 草业学报, 2012, 21(3): 17-25.
[22]罗黎明, 苗彦军, 武建双, 等. 拉萨河谷山地灌丛草地物种多样性随海拔升高的变化特征. 草业学报, 2014, 23(6): 320-326.
[23]刘哲, 李奇, 陈懂懂, 等. 青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响. 生物多样性, 2015, 23(4): 451-462.
[24]中国科学院青藏高原综合科学考察队. 西藏河流与湖泊[M]. 北京: 科学出版社, 1984.
[25]中国科学院青藏高原综合科学考察队. 西藏植被[M]. 北京: 科学出版社, 1988.
[27]何红艳, 郭志华, 肖文发. 降水空间插值技术的研究进展. 生态学杂志, 2005, 24(10): 1187-1191.
[29]刘贤赵, 王国安, 李嘉竹, 等. 中国北方农牧交错带C3草本植物δ13C 与温度的关系及其对水分利用效率的指示. 生态学报, 2011, 31(1): 0123-0136.
[32]王志恒, 陈安平, 朴世龙, 等. 高黎贡山种子植物物种丰富度沿海拔梯度的变化. 生物多样性, 2004, 12(1): 82-88.
[33]王盛萍, 张志强, 张建军, 等. 黄土残塬沟壑区流域次生植被物种分布的地形响应. 生态学报, 2010, 30(22): 6102-6112.
Spatial distribution patterns and environmental interpretation of plant species richness in the Lhasa Valley, Tibet
La-Duo1*, ZHANG Yan-Jie1, LIU Jie1, CUI Ling-Ling1, PANG You-Zhi2
1.College of Science, Tibet University, Lhasa 850000, China; 2.College of Geography and Resources Sciences, Neijiang Normal University, Neijiang 641100, China
Spatial distribution patterns of species richness and potential influencing factors are important aspects of species diversity research and biodiversity conservation. However, little biodiversity research has been carried out in the Lhasa valley. Based on vegetation types, 47 plots with 141 quadrats were established to investigate the spatial distribution of species richness and its relationship with environmental factors. The study showed that a total of 247 species of vascular plant species, belonging to 47 Families and 134 different Genus, were recorded in this study. Climatic data for each plot was calculated using the Cokriging method. Moran's eigenvector maps (MEM) and variation partitioning were generated to explore the influence of spatial structures and environmental factors on the distribution of species richness, and generalized additive model analyses (GAM) were performed to examine the influence of environmental factors and vegetation cover on species richness. De-trended correspondence analysis (DCA) was utilised to detect and analyze the distribution of species richness and its relationships with environment. The results of MEM and variation partitioning indicated that spatial structure was important to the distribution of species richness but environmental factors also played an important role in spatial structure. GAM of environmental factors and transformed species richness (TSR) suggested that distribution patterns of plant species richness are significantly related to longitude, latitude, altitude, AP (annual precipitation), AET (actual evapotranspiration), MAT (mean annual temperature), PET (potential evapotranspiration), MI (moisture index). DCA also supported this result, indicating that these environmental factors influenced the distribution patterns of species richness. Other important factors influencing biodiversity include disturbance and humidity.
species richness; Moran’s eigenvector map; disturbance; climate factors; Lhasa Valley
10.11686/cyxb2015566
2015-12-15;改回日期:2016-01-26
国家自然科学基金-地区科学基金项目(41362001)资助。
拉多(1969-),男,西藏日喀则人,副教授。E-mail: Lhaduo@hotmail.com
Corresponding author. E-mail: Lhaduo@hotmail.com
http://cyxb.lzu.edu.cn
拉多,张燕杰,刘杰,崔玲玲,庞有智. 拉萨河谷植物物种丰富度空间分布格局及其环境解释. 草业学报, 2016, 25(10): 202-211.
La-Duo, ZHANG Yan-Jie, LIU Jie, CUI Ling-Ling, PANG You-Zhi. Spatial distribution patterns and environmental interpretation of plant species richness in the Lhasa Valley, Tibet. Acta Prataculturae Sinica, 2016, 25(10): 202-211.
