基于转录组信息的艾纳香牻牛儿基牻牛儿基焦磷酸合成酶基因(BbGGPS)的克隆及序列分析
2016-11-09夏奇峰刘红昌官玲亮庞玉新
夏奇峰,赵 致,刘红昌,官玲亮,庞玉新
(1.贵州大学 生命科学学院,贵州 贵阳 550025;2.贵州大学 农学院贵州省中药材重点实验室,贵州 贵阳 550025;3.中国热带农业科学院热带作物品种资源研究所/农业部华南作物基因资源与种质创制重点开放实验室/海南省艾纳香工程技术研究中心,海南 儋州 571737)
基于转录组信息的艾纳香牻牛儿基牻牛儿基焦磷酸合成酶基因(BbGGPS)的克隆及序列分析
夏奇峰1,赵致2*,刘红昌2,官玲亮3,庞玉新3
(1.贵州大学 生命科学学院,贵州 贵阳 550025;2.贵州大学 农学院贵州省中药材重点实验室,贵州 贵阳 550025;3.中国热带农业科学院热带作物品种资源研究所/农业部华南作物基因资源与种质创制重点开放实验室/海南省艾纳香工程技术研究中心,海南 儋州 571737)
采用RT-PCR和RACE(Rapid amplification of cDNA ends)技术,从艾纳香(Blumea balsamifera L·DC)的叶片中克隆到二萜化合物合成的关键酶牻牛儿基牻牛儿基焦磷酸合成酶(BbGGPS)基因。结果显示:BbGGPS基因的cDNA全长1475 bp,包含开放阅读框(ORF)1002 bp,编码334个氨基酸;亚细胞结构定位于叶绿体,既非膜蛋白也非分泌性蛋白。疏水性分析显示,BbGGPS是亲水性蛋白。同源性比对结果显示,BbGGPS蛋白与其他植物中GGPS蛋白具有高度的相似性。系统发育分析表明,所有序列被聚为5大类,BbGGPS与菊科植物刺菜蓟聚(Cynara cardunculus var)为一类,表明与其亲缘关系最近。
艾纳香;牻牛儿基牻牛儿基焦磷酸合成酶;氨基酸序列;系统发育分析
艾纳香(Blumea balsamifera L.DC.)为菊科艾纳香属多年生木质草本植物,最早记载于公元741年(唐开元二十九年)陈藏器所编著 《本草拾遗》,此后宋代刘翰等编著《开宝本草》(公元 973 -974 年)也曾记载[1]。艾纳香以根、嫩枝、叶入药,其性微温,味辛、微苦,具有祛风消肿、活血散瘀之功效,可用于治疗感冒、风湿性关节炎、产后风痛、痛经、 外用跌打损伤、疮疖痛肿、湿疹皮炎。另外,艾纳香还具有抗氧化、抗癌和抗病毒的作用[2]。主要分布于我国海南、贵州、广西、广东、云南、台湾等省。在黎族、苗族、壮族等少数民族地区有着悠久的药用历史,是一种重要的民间药物。随着现代药学、分析化学等学科的发展,艾纳香所含有成分的分析已有较多文献报导[3-7]。研究表明,艾纳香中所含成分主要是黄酮类物质和挥发油。黄酮类化合物主要包括黄酮和黄酮苷类物质,挥发油主要包括一系列萜类化合物,从单体组成上划分,可以分为单萜、倍半萜、二萜和三萜等,如L-龙脑、β-石竹烯、γ-桉叶油醇、樟脑、芳樟醇等。
二萜类化合物药理活性广泛,现代药理学研究表面二萜类化合物具有抗肿瘤、抗菌、抗病毒、抑制超敏反应等作用[8-9]。牻牛儿基牻牛儿基焦磷酸合成酶(geranylgeranyl pyrophosphate synthase,GGPS)在二萜生物合成途径中起着关键性的作用,GGPS催化法尼基焦磷酸(FPP)与异戊烯焦磷酸(IPP)形成的牻牛儿基牻牛儿基焦磷酸(GGPP),它是二萜生物合成的共同前体[10]。在GGPP的结构基础上,经过环化异构形成各种阳离子中间产物,之后进一步环化形成一系列二萜化合物。
对GGPS基因的研究已有较多的文献报导,如:马利刚等[11]成功克隆到北葶苈子(Lepidium apetalum)的GGPS基因(LaGGPS),化文平等[12]克隆了丹参(Salvia miltiorrhiza Bunge)的GGPS基因(SmGGPS),李翠萍等[13]克隆了番茄(Solanum lycopersicon L)的GGPS 2基因(SlGGPS2),闵丹丹等[14]克隆了三七(Notoginseng Radix Et Rhizoma)的GGPS基因。目前还未见有关艾纳香GGPS基因(BbGGPS)的研究报导。本课题组基于艾纳香的转录组测序信息,以预测所得的一条可能的GGPS基因序列作为模板,设计特异引物克隆了一条BbGGPS基因,并进行了基因序列与其编码的蛋白质序列的生物信息学分析。艾纳香在分子生物学方面的研究尚未起步,对艾纳香BbGGPS基因进行研究,有望从分子水平揭示艾纳香活性成分代谢途径和调控机制,为提高艾纳香活性成分的含量奠定理论基础。
1 材料与方法
1.1材料
1.1.1植物材料
艾纳香(Blumea balsamifera L·DC)栽培于海南省儋州市,中国热带农业科学院热带作物品种资源研究所南药种质资源圃,试验材料为艾纳香长势良好的叶片。
1.1.2试剂
总RNA提取试剂盒(Plant RNA Kit)、反转录试剂盒、载体连接试剂盒、异丙基硫代半乳糖苷(IPTG)、X-gal、多功能 DNA 纯化回收试剂盒均购自广州飞扬生物工程有限公司。引物合成与测序均由上海美吉(Majorbio)生物工程技术服务有限公司完成。
1.2方法
1.2.1总RNA提取及 BbGGPS全长cDNA的克隆
按照Plant RNA kit试剂盒说明书,提取艾纳香叶片的总RNA,采用RT-PCR合成cDNA第一链。通过从本课题组前期对艾纳香花和叶片转录组的信息,挖掘艾纳香中GGPS相关基因的序列片段,设计特异引物扩增cDNA。正向引物Pf:5’-GTAATTTGCA TCTCATCTGA TATTCG-3’、反向引物Pr:5’-CAAAACGCCA AAACCCAAG-3’,以 cDNA为模板进行PCR扩增。反应体系:PCR反应总体积为50 μL,1.5 mmol/L MgCl2,4种dNTP各200 μmol/L,引物各150 ng,1.5 U Taq plus DNA polymerase(高保真),100 ng cDNA。反应条件:94℃、5 min;94℃、30 s,53℃、30 s,72℃、60 s,30个循环;72℃后延伸10 min。并进行克隆、测序及序列分析。
1.2.2BbGGPS 基因的测序与分析
BbGGPSPCR产物经1.0%的琼脂糖凝胶电泳检测,用胶回收试剂盒对目的片段进行纯化,纯化后将其连接到pEASY-Blunt cloning vector载体上,转化 E.coli Trans1-T1 感受态细胞,涂布于添加氨苄青霉素、IPTG、X-gal 的 LB 平板上,37℃培养箱中培养过夜,随机挑取阳性克隆,使用M13Pf/Pr引物,经PCR检测后扩菌培养,再送样进行测序,获得重组载体pEASY-Blunt-BbGGPS。
1.3艾纳香BbGGPS 基因的生物信息学分析
使用DNAMAN软件分析BbGGPS基因的cDNA序列,找到对应的开放阅读框(ORF)以及对应编码的氨基酸序列;利用DNAMAN软件的多序列比对功能,将BbGGPS氨基酸序列与NCBI中BLAST检索到的高同源性序列进行比对,对保守区域进行考察;使用BioEdit等软件对BbGGPS氨基酸序列的理化性质进行分析,并使用Swiss-model工具对BbGGPS的三维结构在线进行预测;最后利用Mega 6.0将BbGGPS氨基酸序列与从NCBI中进行BLAST比对搜索得到的序列进行系统进化分析。
2 结果与分析
2.1艾纳香BbGGPS基因的克隆
以艾纳香新鲜叶片的cDNA为模板,基于转录组测序预测的GGPS基因序列设计一对特异引物扩增出的PCR产物在1450 bp左右,与预期片段大小一致,电泳结果见图1。将目的条带回收,构建重组载体pEASY-Blunt-BbGGPS转化大肠杆菌,将PCR鉴定为阳性的克隆送样测序。
测序之后得到1475 bp的cDNA序列,通过在线ORF Finder 分析,发现其中包含一个1002 bp的ORF,编码333个氨基酸,碱基序列见图2。将此氨基酸序列在NCBI blastp对比nr数据库,比对结果表明该序列与菊科植物刺菜蓟(Cynara cardunculus var.scolymus)的聚戊烯合成酶序列相似性最高,为86%;其次与地黄(Rehmannia glutinosa)的GGPS序列相似性高达78%。从而可以推测,克隆所得的cDNA序列编码GGPS蛋白,将此基因命名为BbGGPS。
2.2BbGGPS氨基酸序列分析
2.2.1理化性质分析
通过BbGGPScDNA序列预测得到其编码的氨基酸序列,使用 Prot Param tool工具分析其理化性质[11-14],选择软件默认参数。结果显示BbGGPS肽链包含有334个氨基酸残基,分子量为36710.9,等电点(pI)为5.32(图3)。氨基酸的种类分布如 图4 所示,该肽链总共含有20种氨基酸,包括天冬酰胺和谷氨酰胺。含量最多的氨基酸种类为亮氨酸和缬氨酸,其次是亮氨酸和谷氨酸。而半胱氨酸、组氨酸的含量则相对较少,色氨酸仅有一个。带负电荷氨基酸残基总数(Asp+Glu)为45个,带正电荷氨基酸残基总数(Arg+Lys)为36个。总体而言,非极性氨基酸的含量相对较高。不稳定系数为37.92,属于稳定蛋白。
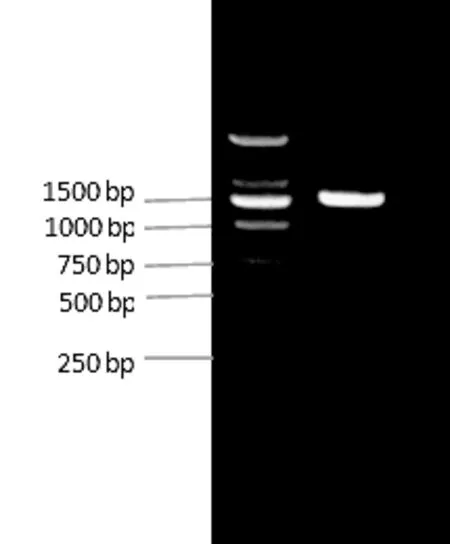
图1 从艾纳香中克隆所得的GGPS基因片段Fig.1 The GGPS gene clone fragment from Blumea balsamifera leaves

图2 艾纳香叶片中BbGGPS基因的cDNA序列(图中框内分别是起始密码子与终止密码子)Fig.2 cDNA sequence ofBbGGPS in Blumea balsamifera leaves(The shadows are initiation codon and termination codon)

图3 BbGGPS氨基酸序列Fig.3 Amino acid sequence of BbGGPS
2.2.2BbGGPS跨膜区、信号肽、亚细胞定位分析
使用TMHMM Sever v.2.0预测BbGGPS的跨膜区域[11-14],输出格式选择为默认的图形化显示(Extensive,with graphics)。分析结果表明,BbGGPS无跨膜区,属于膜外在蛋白;利用ExPASy SignalP 4.0 Serve分析BbGGPS蛋白,并没有发现信号肽,表明该蛋白为非分泌蛋白;用WoLFPSORT工具对BbGGPS进行亚细胞结构定位预测,预测结果表明,BbGGPS最可能定位于叶绿体。
2.2.3疏水性分析
使用ExPASy-ProtScale工具中的Kyte &Doolittle法(K-D法)对艾纳香GGPS蛋白质的疏水性进行在线分析[11-14],窗口参数(Window size)设定为11。分析结果表明,BbGGPS亲水性略大于疏水性,BbGGPS属于亲水蛋白。在第100~150个氨基酸残基之间,有一个强亲水区域,大约在第270个氨基酸残基处,还有一个很强的亲水区(图5)。

图4 BbGGPS的氨基酸组成Fig.4 Amino acid number of BbGGPS
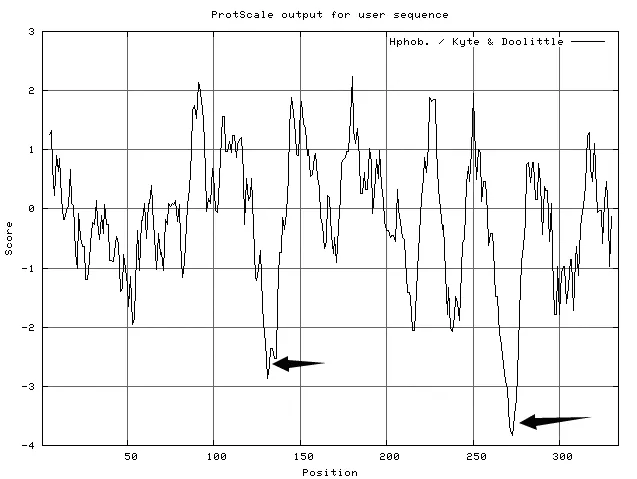
图5 ProtScale分析BbGGPS疏水性的结果(图中箭头所指位置为强亲水区)Fig.5 Hydrophobic analysis of BbGGPS(Strong hydrophilic regions are marked in the picture)
2.2.4蛋白质二级结构预测和三维建模
利用SSpro 4.0(http://download.igb.uci.edu/sspro4.html)对BbGGPS的二级结构进行在线分析[11-14]。结果表明该蛋白二级结构中α-螺旋(H)占 64.8%,β-折叠(E)占 1.7%,无规则卷曲(C)占 33.5%。
使用Swiss-model工具对BbGGPS的三维结构在线进行预测(图6),不使用指定模版,默认原始参数[11-14]。从图中可以看到,BbGGPS的三维结构主要由α-螺旋构成,其次是无规则卷曲。β-折叠只占有很少的一部分,三维结构预测的结果与蛋白质二级结构预测相吻合。分析结果表明,BbGGPS属于α-螺旋型蛋白。其三维结构的“口袋”区域,可能是BbGGPS起催化作用的活性中心所在。
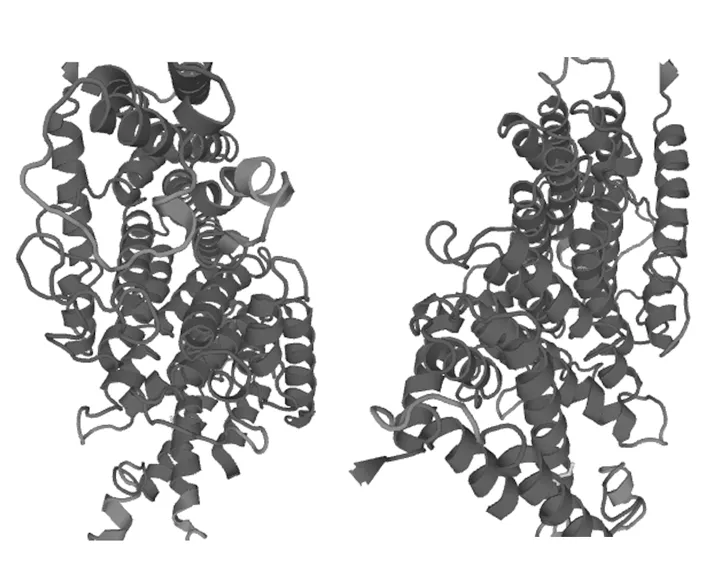
图6 BbGGPS三维结构预测Fig.6 Three dimensional structure prediction of BbGGPS
2.3多序列比对与系统进化分析
2.3.1多序列比对
使用NCBI的BLAST工具,将BbGGPS序列在蛋白质数据库中进行比对搜索。BbGGPS在蛋白质数据库中比对上不同物种的19条蛋白质序列,其中刺菜蓟(Cynara cardunculus var.scolymus)的聚戊烯合成酶序列相似性最高,为86%;其次与地黄(Rehmannia glutinosa)的GGPS序列相似性高达78%。利用 DNAMAN 将BbGGPS氨基酸序列与从NCBI 中挑选的部分同源性较高的已知序列进行多序列比对,结果表明BbGGPS蛋白与其他植物中GGPS蛋白具有高度的同源性(图7)。


图7 BbGGPS 多序列比对结果Fig.7 Multiple sequence alignment of BbGGPS
2.3.2系统进化分析
利用Mega 6.0软件将BbGGPS氨基酸序列与从NCBI中BLAST比对搜索得到的高分值序列(表1)进行系统进化分析,使用除权配对法(UPGMA)构建系统进化树,用差异位点距离(P-distance)估计进化距离。所构建进化树的置信度显著性检验使用自助法(Bootstrap method)。由结果可知位于系统发育树最下方的田七(Panax notoginseng,PnGGPS)、毛果杨(Populus trichocarpa,PtGGPS)、菜豆(Phaseolus vulgaris,PvGGPS)成为一个独立的初级分支,表明这一个分支的物种与其余物种之间亲缘关系较远;拟南芥(Arabidopsis thaliana,AtGGPS)在进化树中作为树根,表明拟南芥相对进化树中的其他物种进化时序最早,进化地位最原始。艾纳香(Blumea balsamifera,BbGGPS)、刺菜蓟(Cynara cardunculus var,CvGGPS)同属于菊科植物,位于一个分支上,表明其亲缘关系较近(图8),这与形态学植物分类结果相似,表明GGPS基因可为植物分类方面的分析或研究提供一定的佐证。
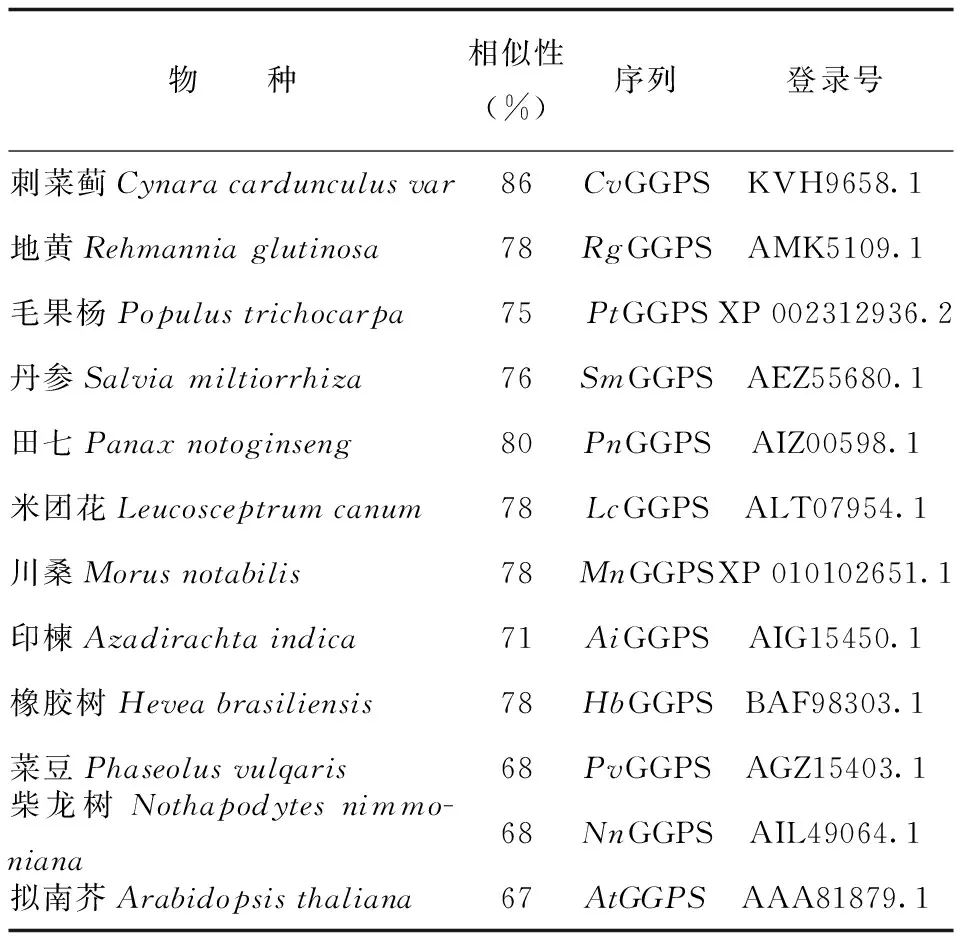
表1 BLAST比对高同源性GGPS序列
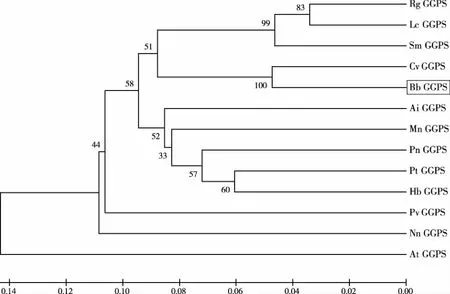
图8 Mega对BbGGPS系统进化分析结果(艾纳香BbGGPS序列用方框标出)Fig.8 system evolution analysis of BbGGPS(Gene sequence of BbGGPS is marked in frame)
3 结论与讨论
本研究基于艾纳香叶片的转录组数据信息,根据挖掘到的GGPS基因的序列片段,设计特异性引物成功从艾纳香总RNA中扩增BbGGPS的全长cDNA序列。将扩增得到的cDNA通过克隆、测序验证,扩增得到的cDNA序列正确。
萜类化合物在植物体中普遍存在,有研究表明,许多高等植物中的药用活性成分均属于萜类化合物。萜类化合物包括单萜、二萜、倍半萜等。目前关于艾纳香中二萜化合物的成分分析与合成机制等方面均未见文献报道,尚属空白。艾纳香GGPS是二萜类物质合成途径中的关键酶,在艾纳香二萜合成机制的研究中具有重要意义。GGPS属于蛋白超家族,在高等植物不同物种之间具有较高的保守性。高等植物不同物种GGPS研究已有较多的文献报导,不论是GGPS cDNA序列长度或是编码的GGPS氨基酸个数都有较大的相似性。例如:北葶苈子GGPS的cDNA全长1146 bp,编码381个氨基酸[11];三七GGPS的cDNA全长1203 bp,编码344个氨基酸[14]。GGPS基因具有的保守结构域主要是聚丙烯合成酶(polyprenylsynthetase);萜类合成酶(terpenoid synthase);聚丙烯相关合成酶(polyprenyl synthetase-related)。使用DNAMAN软件,将艾纳香BbGGPS与同源性较高的多条序列进行比对,发现BbGGPS氨基酸序列存在多个保守结构域,与前人对其他物种的GGPS氨基酸序列研究成果相一致,证明BbGGPS同属于GGPS蛋白超家族中的一员。GGPS在不同高等植物中的进化上具有保守性,因此GGPS氨基酸序列的局部差异可以作为物种进化时序的证据之一。
在本次研究中,通过不同物种间GGPS的氨基酸位点差异构建了系统进化树,结果表明艾纳香与菊科植物刺菜蓟亲缘关系最近,BbGGPS序列的系统发育分析结果与艾纳香已有的种属分类相一致,进一步说明本次分析的结果是具有较高真实性和可信度的。植物中通常含有数目不等的多个GGPS基因,不同的GGPS基因亚细胞定位可能不同,表达模式也不尽相同,在不同的代谢途径中起作用。化文平等已经从丹参中克隆到了三条GGPS基因,实时定量PCR表明丹参的GGPS基因在不同发育时期不同器官中表达差异显著[12]。本次测序得到的艾纳香叶片转录组数据中也有多个不同序列的GGPS基因,有些基因序列并不完整,有待将来通过RACE技术获取全长序列之后进一步研究其生物学功能。本实验克隆得到了艾纳香二萜化合物合成的关键酶基因 GGPS,并对所得序列进行了比较详细的生物信息学分析,以期从分子生物学的角度揭示艾纳香二萜生物合成的内在机制,为艾纳香的后续深入研究奠定了基础。
[1]袁媛,庞玉新,王文全,等.艾纳香属系统分类研究进展[J].热带农业科学,2011,31(3):81-86.
[2]官玲亮,庞玉新,王丹,等.中国民族特色药材艾纳香研究进展[J].植物遗传资源学报,2012,13(4):695-698.
[3]李璞,陈宇琼,黄火强.艾纳香化学成分与药理活性研究进展[J].实用中医内科杂志,2012,26(10):3-6.
[4]严启新,谭道鹏,康晖.艾纳香中的黄酮类化学成分[J] .中国实验方剂学杂志,2012,18 (5):86-89.
[5]邓芹英,丁丛梅,张维汉.艾纳香中黄酮化合物的研究[J].波谱学杂志,1996,13(5):447-452.
[6]赵金华,康晖,姚光辉,等.艾纳香化学成分研究[J].中草药,2007,38(3):350-352.
[7]白志文,朱露,孙济平,等.苗药艾纳香研究进展[J].中国民族医药杂志,2012,7(7):65.
[8]Gershenzon J,Dudareva N.The function of terpenenatural products in the natural world [J].Nat Chem Biol,2007,3(7):408-414.
[9]海广范,张慧,郭兰青.二萜类化合物药理学作用研究进展[J].新乡医学院,2015,32(1):77-80.
[10]陈建,赵建刚,植物萜类生物合成相关酶类及其编码基因的研究进展[A].分子植物育种,2014,2(6):757-764.
[11]马利刚,赵乐.北葶苈子GGPS基因的克隆、序列分析与原核表达[J].中医药现代化,2015,3(7):154-156.
[12]化文平,宋双红.丹参 SmGGGPS3 基因的克隆及表达分析[J].植物科学学报 2014,32(1):50-57.
[13]李翠萍,夏蓓蓓.番茄 GGPS2 基因的克隆与表达载体的构建[J].西南大学学报.2010,32(10):65-67.
[14]闵丹丹,唐美琼.三七香叶基香叶基焦磷酸合酶基因的克隆及表达分析[J].中国中药杂志.2015,40(11):54-58.
Analysis of geranylgeranyl pyrophosphate synthase (GGPS) sequence of Blumea balsamifera(L.)·DC based on transcriptome information
XIA Qi-feng1,ZHAO Zhi2*,LIU Hong-chang2,GUAN Ling-liang3*,PANG Yu-xin3
(1.Guizhou University,College of Life Sciences,Guiyang,Guizhou 550025,China;2.Guizhou University College of Agriculture,Guiyang,Guizhou 550025,China;3.Tropical Crops Genetic Resources Institute,Chinese Academy of Tropical Agricultural Sciences,Danzhou,Hainan 571737,China)
A geranylgeranyl diphosphate synthase gene,designatedBbGGPS,has been isolated from Blumea balsamifera(L.) DC using reverse transcription polymerase chain reaction approach (RT-PCR) and rapid amplification of cDNA ends (RACE) methods.The results showed that the BbGGPS cDNA had a full length of 1 475 bp,and contained an open reading frame predicting a polypeptide of 334 amino acids.Hydropathy and subcellular localization prediction showed that the BbGGPS belongs to hydrophilic protein and is located in Chloroplasts.It is neither a membrane protein nor secretory protein.BbGGPS protein showed a high similarity with other plant GGPS genes.Phylogenetic analysis indicated that all the amino acid sequence were divided into five categories and BbGGPS was grouped with that of Cynara cardunculus var.
Blumea balsamifera;geranylgeranyl diphosphate synthase;amino acid sequence;phylogenetic analysis
2016-05-21;
2016-06-14
国家自然科学基金(NO.81202910);中央级公益性科研院所基本科研业务费专项资金(1630032014015);海南省中药现代化专项(ZY201410)。
赵致(1957-),男,教授,主要研究方向:作物生理生态理论及其应用;E-mail:zzhao@gzu.edu.cn。
Q756
A
1008-0457(2016)04-0023-07国际
10.15958/j.cnki.sdnyswxb.2016.04.005
