39个樱花品种亲缘关系的ISSR分析
2016-11-09王志龙林乐静
林 立 王志龙 付 涛 林乐静
(宁波城市职业技术学院宁波市园林植物开发重点实验室,宁波 315502)
* 通信作者:E-mail:wangzhl01@163.com
39个樱花品种亲缘关系的ISSR分析
林 立 王志龙*付 涛 林乐静
(宁波城市职业技术学院宁波市园林植物开发重点实验室,宁波 315502)
樱花为世界著名的早春观赏花木之一,品种繁多、来源复杂,造成了樱花品种在分类、鉴定上的困难。为探讨ISSR分子标记方法在樱花品种分类上应用的可行性,对引自日本的39个樱花品种进行ISSR亲缘关系分析。选用14条扩增带型清晰且重复性好的引物共获得109条谱带,其中102条呈多态性,多态性比例(PPB)为93.58%,表明樱花品种遗传多样性较高。39个樱花品种的Nei’s遗传相似性系数介于0.493~0.942,平均值为0.727,表明品种间亲缘关系较近。根据Nei’s遗传相似性系数在0.697处,UPGMA聚类结果显示39个品种分为两个类群,在0.738处可进一步分为4个亚类群,聚类结果较好地支持了川崎哲也的樱花7组分类标准,也表明ISSR分子标记方法适用于樱花品种的分类。聚类结果还表明樱花的花序、花型和花色也可作为品种分类的重要指标。
樱花;ISSR;亲缘关系;UPGMA
樱花是世界著名的早春观赏花木之一,隶属于蔷薇科(Rosaceae)李亚科(Prunoideae)樱属(Cerasus),主要分布于中国、日本和朝鲜等国家[1]。据统计,目前全世界培育的樱花品种有200多个,其中大部分来自日本[2]。樱花品种繁多,花形、花色美丽,树姿洒脱开展,盛开时如玉树琼花,堆云叠雪,甚是壮观,被广泛用于公园、学校、街道、庭院等区域栽植绿化[3]。
自上世纪早期开始,国际樱花市场不断扩大,野生樱花资源的发掘及杂交育种工作也逐步加强,樱花栽培新品种不断涌现。但是,在新品种数量快速增加的同时也造成了诸如栽培品种混乱、命名不规范、种系定位不一、亲本来源不明等问题,严重影响了樱花产业的发展。从上世纪70年代开始,品种认定工作已经成为了樱花研究的重要内容[1~7]。1974年,本田正次和林弥荣将日本樱花品种分为六个组[4],奠定了日本樱花品种分类的基础。后来,Kawasaki对该分类标准进行了相应的完善,将江户彼岸群(组)独立分为一个组,形成了樱花的7个群(组)分类标准[5],得到了国际社会的广泛认可[6]。但该分类方法主要依据樱花品种的种系进行划分,对许多起源不明确或具有多亲本来源品种的分类鉴定具有局限性。2007年,时玉娣对樱属部分品种资源进行了分类研究,依据萼筒、花序、毛被、花型等表型性状建立了樱属品种分类的五级标准,较好地对一些种系及其下品种进行了分类研究[7]。但该方法易受品种的栽培环境、营养状况及砧木品种等条件造成的表型差异所影响,也具有一定的局限性。
目前,国内外有关樱花的研究多集中于形态分类[3~7]、化学成分分析[8~10]、病虫害防治[11~13]以及繁殖技术[14~15]等方面。近年来,分子标记方法凭借其高效性和可靠性逐渐被用于樱属植物的分类研究[16~23]。然而,相关研究主要针对于樱属原种[16~20],对樱花品种进行分子水平的分类研究仍然少见[21~23]。Wünsch等[21]和Antonius等[22]通过SSR分子标记分别对西班牙的一些甜樱桃栽培品种和芬兰地区的樱花栽培品种进行了品种鉴定。Kato等[2]利用SSR分子方法鉴定了222个日本樱花品种的无性系遗传变异情况,结果表明大部分品种的无性系具有单一而独特的基因型,因此SSR标记适用于大部分樱花品种的鉴定,但SSR标记较低的多态性不利于鉴定具有相近遗传背景的品种。周春玲等[23]通过RAPD分子标记方法研究了青岛市19个樱花品种的分类,较好地对樱花品种进行了区分,但所选品种数量有限,许多国内常见品种未被研究。ISSR作为一种更新型、便捷、多态性更高的分子标记方法,在许多植物品种的鉴定和亲缘关系分析等方面得到了广发应用[24~25]。本研究通过ISSR分子标记方法对国内主栽的39个日本樱花品种进行亲缘关系分析,以期了解樱花品种的遗传背景,并探讨ISSR分子标记方法在樱花品种分类上应用的可能,为今后樱花品种的鉴定、分类以及育种等方面的研究提供一定的理论依据。
1 材料与方法
1.1 材料
供试材料采自宁波绿野山庄樱花种植园,采集39个国内主栽的日本樱花品种的嫩叶样本(表1),硅胶干燥处理后放-40℃冰箱中保存备用。各品种的命名主要参考王贤荣[1]和Kato等[2]的研究并依据《国际栽培植物命名法规》进行规范,同时记录各品种的部分表型特征。
1.2 DNA提取与PCR
参考南程慧[14]的方法提取樱花基因组DNA。从美国哥伦比亚大学设计的通用引物(UBC set no.9)中筛选出14条ISSR引物(表2),由生工生物工程(上海)股份有限公司合成。ISSR-PCR反应体系确定为:10×Buffer缓冲液2 μL,25 mmoL·L-1MgCl21.6 μL,2 U Taq酶0.5 μL,10 mmoL·L-1dNTP 0.5 μL,10 μmoL·L-1引物0.6 μL,50 ng·L-1DNA模板1 μL,14.8 μL无菌水。PCR扩增程序为,94℃预变性5 min;94℃变性30 s,各种温度退火40 s,72℃延伸1.5 min,38个循环;72℃延伸8 min,4℃保存。扩增产物在5 V/cm电压下电泳1 h,电泳结果在凝胶成像系统(GelDoc-IT2 310)中观察并拍照记录。
1.3 数据处理与分析
统计条带时参照石颜通等[26]的方法将电泳图谱信息转化成0/1二元矩阵。用POPGEN1.32软件计算多态性条带百分比PPB(The percentage of polymorphic bands)、有效等位基因数(Ne)、Nei’s基因多样性指数(H)、Shannon’s信息指数(S)和品种间的Nei’s遗传相似性系数(I)。利用NTSYSpc2.10e软件根据品种间的遗传相似性系数按UPGMA(Unweighted pair group method using arithmetic averages)法构建聚类图,并进行主坐标分析(PCoA)。
2 结果与分析
2.1 扩增产物的多态性分析
39份樱花样品进行ISSR-PCR扩增后,14条引物共扩增出109条带,片段长度范围集中在100~2 000 bp,其中多态性条带102条,多态性条带百分比(PPB)为93.58%(表2)。每条引物可得4~15条DNA带,平均得7.79条带,表明樱花基因组DNA多态性较高。

表1 供试材料
注:“*”表示命名参考王贤荣《中国樱花品种图志》最新命名方法。
Note:“*”represents the name of some cultivars according to An Illustrated Monograph of Cherry cultivars in China.
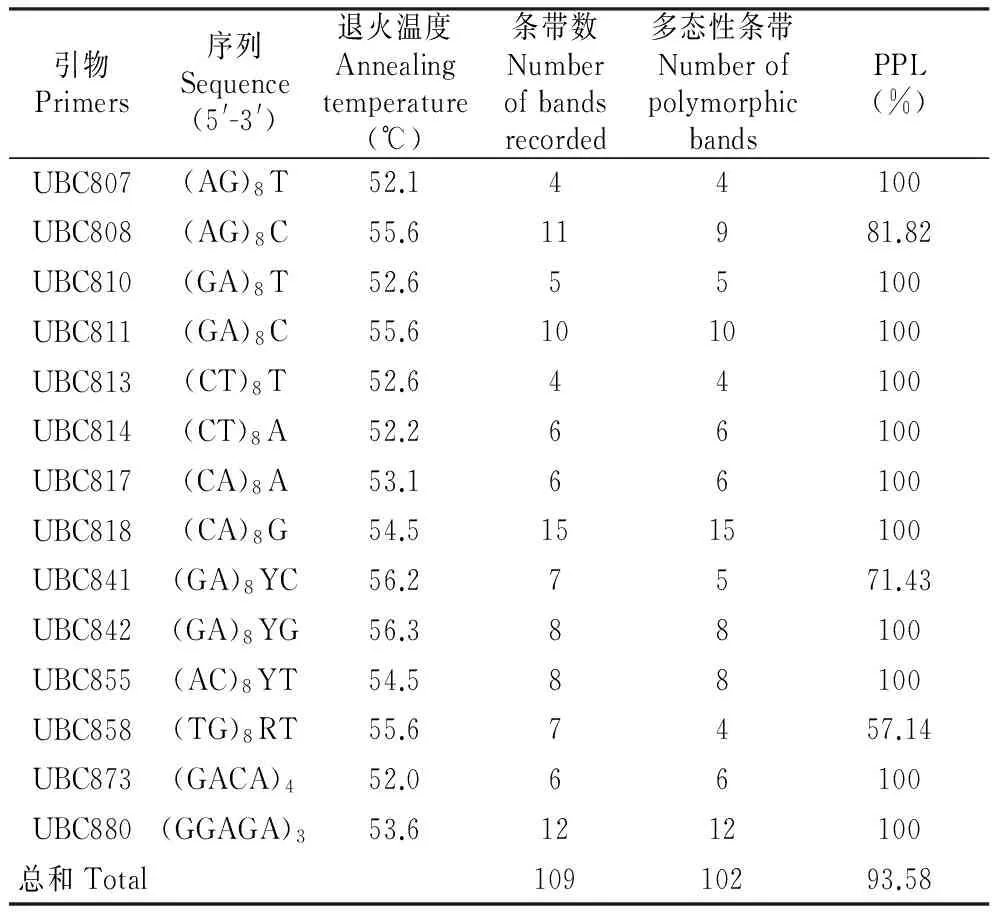
表2 ISSR分析的引物序列
Y=(C,T);R=(A,G)
利用软件包POPGENE1.32计算各位点的有效等位基因数(Ne)、Nei’s基因多态性指数(H)以及Shannon’s信息指数(S)。结果表明供试樱花品种平均有效等位基因数(Ne)为1.440 5(0.347 4),平均Nei’s基因多样性指数为0.265 6(0.171 5),平均Shannon’s信息指数为0.410 5(0.223 7)。
2.2 品种间亲缘关系分析
利用POPGEN1.32软件计算供试品种间的遗传相似性系数。39个样品间Nei’s遗传相似性系数介于0.493~0.942(表3),平均遗传相似性系数为0.727。一叶与天之川之间遗传相似性系数最大(I=0.942),显示两者具有较近的亲缘关系。八重豆樱与染井吉野之间遗传相似性系数最小(I=0.493),表明它们彼此存在较大的遗传差异。
2.3 品种间聚类分析结果
基于ISSR扩增结果,按UPGMA聚类法进行品种间遗传关系分析,得到39个供试品种的亲缘关系树状图(图2)。以遗传相似性系数0.697为阈值,39个品种可划分为两大类群,冬樱和八重豆樱聚为类群I,其余品种聚为类群Ⅱ。进一步以遗传相似性系数0.738为阈值,则类群Ⅱ可分为4个亚类群,亚类群Ⅱb包含雨情枝垂、十月、红华和小松乙女4个品种;亚类群Ⅱc包含花笠、一叶、天之川、大岛樱、御衣黄和松月6个品种;亚类群Ⅱd包含江户彼岸(即大叶早樱)[1,20]、神代曙、白雪、思川、红枝垂和染井吉野6个品种,其余品种归于亚类群Ⅱa。亚类群Ⅱa在阈值为0.787处,又可分为6个小组,其中小组1包含寒绯樱、修善寺寒樱、寒樱、大渔樱、大寒樱和河津樱6个品种;小组2包含骏河台匂、杨贵妃和关山3个品种;小组3包含静匂、福绿寿、红笠、奈良八重樱和小彼岸5个品种;小组4包含米国和嘉奖2个品种;小组5含有及时雨、兰兰、手弱女3个品种,其余聚于小组6中。

图1 39个樱花品种的UPGMA聚类图Fig.1 UPGMA dendrogram for 39 cultivars of Cerasus plants

2.4 主坐标分析
基于39份供试材料的遗传相似性系数,利用NTSYSpc2.10e软件进行主坐标分析,第一、第二、第三主坐标的贡献率分别为12.45%,10.54%和9.12%。对39份材料做主坐标分析并构建二维散点图(图2),其结果与聚类分析结果基本一致,只有大岛樱品种有一定差异。在UPGMA聚类中,大岛樱聚于亚类群Ⅱc中,而主坐标分析结果却显示大岛樱与亚类群Ⅱc中品种亲缘关系更近,可能是第四、第五贡献值忽略的结果。

图2 39个樱花品种的主坐标分析图Fig.2 Principal coordinate analysis for the 39 cultivars of Cerasus
3 讨论
ISSR分子标记技术以SSR为引物扩增微卫星之间的区域,其原理与SSR、RAPD相似,只是引物要求不同,具有信息量大、结果稳定性高、不受环境条件控制的优点[27],因此这种检测方法可以对常规形态学、细胞学特征等分类方法起到一个补充的作用。本研究中供试樱花品种的多态性条带百分比(PPB)为93.58%,平均Nei’s基因多样性指数为0.268 3,表明所选樱花品种间存在丰富的遗传变异。并且ISSR在樱花品种间的扩增位点多态性主要由品种间的差异引起,可有效地反映品种间的亲缘关系,因而可作为樱花品种间亲缘关系分析的有效手段。
樱花栽植历史悠久,在长期的生产实践中选育出了许多优良品种,其中许多品种具有共同的祖先。Kawasaki在本田正次和林弥荣分类标准基础之上,按种系将樱花品种分为七个组,即寒绯樱组、山樱组、豆樱组、丁字樱组、樱桃组、深山樱组和江户彼岸组[5]。按此分类标准,39份日本樱花品种分别为寒绯樱组(6种)、山樱组(20种)、豆樱组(2种)以及江户彼岸组(11种)品种[1~2,5],UPGMA聚类结果也较好地支持了该分类标准,并且聚类分析结果也得到了主坐标分析结果的验证。在阈值为0.712时,豆樱组、山樱组和江户彼岸组区分较好。冬樱的亲本之一为豆樱[1],与八重豆樱都属于豆樱组,故而聚为类群Ⅰ。类群Ⅱ中为寒绯樱组、山樱组和江户彼岸组混合品种,其中寒绯樱组和山樱组品种间的亲缘关系较近,与江户彼岸组樱花则相对较远。在亚类群Ⅱa中主要为寒绯樱组和山樱组品种,寒绯樱组聚于第1小组,其中的修善寺寒樱、寒樱、大渔樱、大寒樱和河津樱,具有共同的祖先寒绯樱[1],遗传背景较为相近,故6个品种亲缘关系较近(平均Nei’s遗传相似性系数I=0.827),花序、花型、花色等表型也相似,其中花色最为典型,都带紫色。其余5个小组中除了第3小组中的小彼岸以及第4小组中的米国和嘉奖为江户彼岸组品种外其余皆为山樱组品种,但米国和嘉奖都具有山樱组樱花的遗传背景。在亚类群Ⅱb和亚类群Ⅱd中,除白雪外9个品种的来源都较为明确,有共同的祖先江户彼岸,9个品种的花色都是淡红色,树型以江户彼岸的伞形为主,花序大多为伞形。白雪品种一般认为是江户彼岸经过复杂的杂交而形成的,UPGMA聚类结果支持了该推断。两个亚类群之间的表型差异主要体现在花型,亚类群Ⅱd中花型单瓣为主,而亚类群Ⅱb以半重瓣为主,与山樱组樱花以半重瓣、重瓣为主的表型特征相近,推测该亚类群品种可能受山樱组的遗传影响更大。亚类群Ⅱc中,一叶、天之川、御衣黄、松月和大岛樱花序都是伞房或伞形花序,亲缘关系也较近,由于大岛樱为单瓣型较古老的品种[1],因此推测大岛樱可能为另4个品种的共同祖先。
基于ISSR分子标记扩增结果的聚类分析表明遗传背景相近的樱花品种间具有较近的亲缘关系。花序、花型、花色相近的樱花品种也显示出了一定的亲缘关系,表明这些表型性状可作为樱花品种分类的关键指标之一。本研究中,单瓣品种主要为寒绯樱组品种和江户彼岸组品种,其中寒绯樱组花色偏紫,江户彼岸组樱花花色偏淡红,重瓣品种主要为山樱系。本研究未发现一些与樱花瓣型、花色等相关的特异的扩增片段,无特征谱带,因此还需开发更多的引物应用于樱花品种遗传背景分析。研究结果还表明,樱花品种基因型具有高度杂合性,品种间多型性基因位点较多,难以建立基因位点与主要表型性状如花型、花色之间的联系。
我国樱花资源丰富,樱属植物有48种,但我国开展樱花研究工作起步较晚,较日本等国家有一定差距[7]。目前,国内樱花品种以日本引进为主,品种自主选育、改良能力还不够,严重制约了我国樱花产业的发展。同时,在引进新品种时,对于品种(系)情况了解欠缺,不利于我国今后樱花选育工作的开展。针对这些问题,笔者认为今后在引进新品种时严格区分品种的种系,查清其遗传背景,建立科学、规范的命名标准。同时,加强我国野生樱属植物资源的开发力度,积极开展樱花新品选育、病虫害防治等工作。
4 结论
研究表明ISSR分子标记可以较好地用于樱花品种间亲缘关系的分析,对于追溯品种起源、厘清品种的分类情况具有较高的可行性。基于ISSR研究的UPGMA聚类结果支持了日本学者Kawasaki对日本樱花品种的分类标准,并证实了白雪品种起源于江户彼岸的说法,推测出了大岛樱可能为一叶、天之川、御衣黄和松月等品种的祖先。在寒绯樱组樱花与山樱组樱花之间分类研究上,ISSR分子标记方法未能表现出良好的区分度,但寒绯樱组樱花自身表型特点突出,开花较早,花型以单瓣为主,花色略带紫红色,易与山樱组品种区分,因此ISSR分子标记方法可作为樱花品种五级分类方法[1,7]之外的一种辅助手段。研究结果也证实了花序、花型、花色等表型性状为樱花品种分类的关键指标。
1.王贤荣.中国樱花品种图志[M].北京:科学出版社,2014.
2.Kato S,Matsumoto A,Yoshimura K,et al.Clone identification in Japanese flowering cherry(PrunussubgenusCerasus) cultivars using nuclear SSR markers[J].Breeding Science,2012,62(3):248-255.
3.刘晓莉,赵绮,舒美英,等.18个樱花品种花部形态性状初步研究[J].江苏农业科学,2012,40(4):185-187.
4.林田正次,林弥荣.日本的樱花[M].东京:城文堂新光社,1974.
5.Kawasaki T.Flowering cherries of Japan[M].Tokyo:Yama-kei Publishers,1993.
6.Ohba H,Kawasaki T,Tanaka H.Flowering cherries of Japan(New Edition)[M].Tokyo:Yama-kei Publishers,2007.
7.时玉娣.樱属品种资源调查及分类研究[D].南京:南京林业大学,2007.
8.Yook H S,Kim K H,Park J E,et al.Antioxidative and Antiviral Properties of Flowering Cherry Fruits(PrunusserrulataL.var.spontanea)[J].The American Journal of Chinese Medicine,2010,38(5):937-948.
9.翁仕洋,成露露,查文,等.日本晚樱叶挥发油化学成分的GC-MS分析[J].安徽农业科学,2013,41(7):3111-3112.
10.Lei G M,Wang L,Liu X,et al.Chemical composition of essential oils and hydrosols from fresh flowers ofCerasussubhirtellaandCerasusserrulatafrom East China[J].Natural Product Research,2014,28(21):1923-1925.
11.Matic S,Minafra A,Sánchez-Navarro J A,et al.‘Kwanzan Stunting’ syndrome:Detection and molecular characterization of an Italian isolate of Little cherry virus[J].Virus Research,2009,143(3):61-67.
12.Song G Q,Sink K C,Walworth A E,et al.Engineering cherry rootstocks with resistance toPrunusnecrotic ring spot virus through RNAi-mediated silencing[J].Plant Biotechnology Journal,2013,11(5):702-708.
13.王志龙,金杨唐,谭志文,等.宁波樱花根癌病病原细菌鉴定[J].植物保护,2014,40(3):147-150.
14.南程慧.迎春樱居群变异与繁殖生物学研究[D].南京:南京林业大学,2012.
15.李蒙.山樱花高山居群生态学特种及组织培养[D].南京:南京林业大学,2013.
16.刘艳玲,徐立铭,程中平.基于ITS序列探讨核果类果树桃、李、杏、梅、樱的系统发育关系[J].园艺学报,2007,34(1):23-28.
17.Gharahlar A S,Zamani Z,Fatahi R,et al.Estimation of genetic diversity in some Iranian wildPrunussubgenusCerasusaccessions using inter-simple sequence repeat(ISSR) markers[J].Biochemical systematic and ecology,2011(8),39:826-833.
18.Öz M H,Vurgun H,Bakir M,et al.Molecular analysis of East Anatolian traditional plum and cherry accessions using SSR markers[J].Genetics and Molecular Research,2013,12(4):5310-5320.
19.Shi S,Li J L,Sun J H,et al.Phylogeny and Classification ofPrunussensulato(Rosaceae)[J].Journal of Integrative Plant Biology,2013,55(11):1069-1079..
20.Cho M S,Kim C S,Kim S H,et al.Molecular and morphological data reveal hybrid orgin of wildPrunusyedoensis(Rosaceae ) From Jeju island,Korea:implications[J].American Journal of Botany,2014,101(11):1976-1986.
21.Wünsch A,Hormaza J I.Molecular characterisation of sweet cherry(PrunusaviumL.) genotypes using peach(Prunuspersica(L.) Batsch) SSR sequences[J].Heredity,2002,89(3):56-63.
22.Antonius K,Aaltonen M,Uosukainen M,et al.Genotypic and phenotypic diversity in Finnish cultivated sour cherry(PrunuscerasusL.)[J].Genetic Resources and Crop Evolution,2012,59(3):375-388.
23.周春玲,陈芳,韩德铎,等.青岛市19个樱花品种的RAPD分析[J].西北植物学报,2007,27(12):2559-2563.
24.季志仙,王美兴,范宏环,等.基于ISSR指纹的甘薯食用品种的遗传多样性分析[J].核农学报,2014,28(7):1197-1202.
25.胡仲义,林立,刘夔,等.26个川茶花品种亲缘关系的ISSR分析[J].植物研究,2013,33(5):629-634.
26.石颜通,周波,张秀新,等.牡丹89个不同种源品种遗传多样性和亲缘关系分析[J].园艺学报,2012,39(12):2499-2506.
27.Hu Z Y,Lin L,Deng J F,et al.Genetic diversity and differention among populations ofBretschneiderasinensis(Bretschneideraceae),a narrowly distributed and endemic species in China,detected by inter-simple sequence repea(ISSR)[J].Biochemical Systematics and Ecology,2014,56(6):101-110.
GeneticRelationshipsof39CerasusCultivarsbyISSRAnalysis
LIN Li WANG Zhi-Long*FU Tao LIN Le-Jing
(Ningbo Key Laboratory of Landscape Plant Development,Ningbo City College of Vocational Technology,Ningbo 315502)
Cerasustrees are famous ornamental plants in the world with a large number of cultivars, but the origin of mostCerasuscultivars are poorly known, resulting in some difficulties in cultivars classification. We analyzed the genetic relationships among 39 cultivars ofCerasusby inter-simple sequence repeat(ISSR) molecular-marked technique, and screened 14 ISSR primers to assess the genomes of 39 cultivars ofCerasus. The result showed that a total of 109 DNA bands were amplified and 102 of which(93.58%) were polymorphic. The Nei’s genetic similarities of 39 cultivars ranged from 0.493 to 0.942 with the average of 0.727, suggesting that the genetic similarities among the 39 cultivars were relatively close. According to the Nei’s genetic similarity of 0.697, by UPGMA method cluster analysis, 39 cultivarss were classified into 2 cluster groups, and the group Ⅱ were classified into 4 subcluster groups with the genetic similarity of 0.738. The result not only supported the classification method of Kawasaki Tetsuya which classifiedCerasuscultivars into 7 groups according their origin, but also showed that ISSR technique was applied to classifyCerasuscultivars. From cluster results, flower inflorescence, form and colour were important indicators forCerasuscultivars classification.
Cerasuscultivars;ISSR;genetic relationships;UPGMA
宁波市科技局农业重大专项(2014C11002);宁波市农业社会发展重点项目(2012C10010);国家星火项目(2012GA701014)
林立(1985—),男,博士研究生,助理实验师,主要从事植物分子生物学研究。
2015-10-08
Q949.751.8
A
10.7525/j.issn.1673-5102.2016.02.020
