两种病因弱视幼猫视网膜中NOS和GABA的表达
2016-11-09王晗敏莫利娟李青松张兴儒
王晗敏,荣 翱,莫利娟,李青松,张兴儒
·实验论著·
两种病因弱视幼猫视网膜中NOS和GABA的表达
王晗敏1,荣翱2,莫利娟1,李青松1,张兴儒1
1Department of Ophthalmology, Putuo Hospital, Shanghai University of Traditional Chinese Medicine, Shanghai 200062, China;2Department of Ophthalmology, Tongji Hospital, Shanghai 200065, China
•METHODS:A total of 18 aged 3-week kittens were randomly divided into monocular deprivation, strabismus and normal groups. All types of amblyopia were developed in the experimental eyes that were detected by P-VEP 12wk later. The cats were killed and the immunocytochemistry staining method were applied to observe under the light microscope the changes of distribution and positive cells areas of NO and GABA across the amblyopic retinal, compared to that from the normal cats of identical age.
•RESULTS: The P-VEP showed that the amplitude of wave P1 was lower (P<0.05) and the P1 latent time was longer (P<0.05) in two types of amblyopic cats than those in the normal cats. Compared to the normal cats, the NO and GABA positive cells areas were obviously reduced (P<0.05) across the retina in the amblyopic cats. But no significant difference was found between two kinds of amblyopic cats.
•CONCLUSION:The NO and GABA play an important role in the formation of amblyopia in the level of retinal.
目的:分别利用单眼睑缝合法和外直肌离断法建立两种不同病因的幼猫弱视模型,探讨一氧化氮(NO)和γ-氨基丁酸(GABA)这两种神经递质在两种弱视形成过程中的作用。
方法:3周龄幼猫18只,随机分为正常组、形觉剥夺组、斜视组,利用单眼睑缝合法和外直肌离断法建立两种病因的幼猫弱视模型,12wk后行图形视觉诱发电位检测。摘取眼球做病理切片,使用免疫组化染色法检测两组弱视眼和正常组视网膜中NOS和GABA的表达。
结果:形觉剥夺组、斜视组12wk与同龄正常组图形视觉诱发电位(P-VEP)比较,均可见P1波潜伏期延长、振幅下降,差异有统计学意义(P<0.05)。弱视组视网膜内的NOS和GABA免疫阳性神经元染色淡,平均阳性面积减少,与同龄正常组相比差异有统计学意义(P<0.05)。弱视组间视网膜中的NOS和GABA平均阳性面积差异无统计学意义(P>0.05)。
结论:NO和GABA的水平降低可能导致弱视眼视网膜内神经兴奋性的改变,二者在弱视形成过程中起着重要作用。
弱视;图形视觉诱发电位;一氧化氮;γ-氨基丁酸
引用:王晗敏,荣翱,莫利娟,等.两种病因弱视幼猫视网膜中NOS和GABA的表达.国际眼科杂志2016;16(11):2006-2009
0 引言
弱视是儿童的常见眼病,据文献报道其患病率为2%~4%[1]。临床上以斜视性弱视与形觉剥夺性弱视最为常见。目前弱视发病机制的研究主要认为是神经网络、神经环路、突触连接等不同水平的可塑性学说,其具体机制并不完全清楚。研究发现,在连接精细化发育(突触连接的可塑性)过程中,一氧化氮(nitric oxide,NO)作为逆行信使参与了突触前后的相互通讯,而γ-氨基丁酸(γ-aminobutyric acid,GABA)则作为一种重要的视觉神经递质参与了视觉这种复杂的神经活动。本研究建立两种病因弱视模型,通过观察一氧化氮合酶(NOS)和γ-氨基丁酸(GABA)在视网膜中的表达,探讨NO和GABA这两种神经递质在弱视形成过程中的作用。
1 材料和方法
1.1材料实验选用3周龄普通家养幼猫18只,雌雄不论,常规检查双眼,确认外眼、屈光间质以及眼底无异常。随机分为3组:单眼形觉剥夺组、单眼斜视组、正常对照组,每组各6只。
1.2方法
1.2.1斜视幼猫模型制备[2]每只幼猫随机挑选1眼,共6只6眼。30g/L戊巴比妥钠30mg/kg腹腔麻醉,侧卧位固定,常规消毒,开睑,颞侧球结膜下注射少量利多卡因,沿角膜缘外2mm剪开球结膜并分离周围筋膜组织,充分暴露外直肌并勾出后沿附着点离断,予氧氟沙星眼液滴眼,每日3次。术后采用角膜映光法测得平均内斜15°~25°。
1.2.2形觉剥夺性斜视模型制备每只幼猫随机挑选1眼,共6只6眼。30g/L戊巴比妥钠30mg/kg腹腔麻醉,20g/L利多卡因0.4mL睑缘皮下浸润麻醉,侧卧位固定,剪除睑缘周围毛发,常规消毒,剪除中内1/3处至外眦1~1.5mm宽的上下睑缘,分层缝合皮肤及皮下组织,封闭睑裂。术眼纱布包敷,庆大霉素冲洗眼睑每日1次。
1.2.3弱视模型鉴定各组幼猫饲养12wk后,采用重庆国特视觉电生理仪完成图形视觉诱发电位(pattern visual evoked potential,P-VEP)的记录。将麻醉后的猫固定,调整猫的头位,置刺激显示屏于被测眼前50cm,并保证与被测眼在同一水平,遮挡对侧眼。以翻转棋盘格作为刺激,对比度96%,屏幕亮度140cd/m2,刺激频率2.4Hz,分析时间250ms,叠加100次,所采集的电信号输入计算机处理后并打印。
1.2.4眼球标本制备及免疫组化染色40mg/kg戊巴比妥钠腹腔注射进行深度麻醉处死动物,完整摘除眼球,沿角膜缘剪开眼球,去除晶状体及玻璃体,取眼球壁,以中央凹为中心取直径5mm的中央区,放入组织固定盒,标号后置于4%多聚甲醛的PBS缓冲液中,常温固定24h。行石蜡包埋切片,常规脱蜡至水,30mL/L H2O2浸泡10min,磷酸盐缓冲液(PBS)洗3次×3min,微波辐射抗原修复大约20min。滴加正常山羊血清封闭后加1∶100稀释的兔抗人NOS及兔抗人GABA,以PBS代替一抗,水洗,苏木素衬染30s,水洗,盐酸乙醇蓝化2s,水洗,微波蓝化,常规树脂封片。光镜下拍摄图片,计算机图像分析测量免疫组化图像中阳性区域面积。
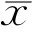
2 结果
2.1P-VEP记录结果统计学分析,12wk时,单眼形觉剥夺组的P波潜伏期较正常组明显延长,差异有统计学意义(t=7.478,P<0.05),单眼斜视组的P波潜伏期较正常组明显延长,差异有统计学意义(t=6.558,P<0.05),单眼形觉剥夺组的P波振幅较正常组明显降低,差异有统计学意义(t=5.168,P<0.05),单眼斜视组的P波振幅较正常组明显降低,差异有统计学意义(t=5.295,P<0.05,表1)。
表1形觉剥夺组、斜视组与正常组P1波比较
±s
表2弱视组与正常组视网膜NOS及GABA阳性面积比较
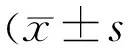
,μm2)
注:aP<0.05,bP<0.01vs正常组。
2.2弱视眼视网膜中NOS阳性细胞面积变化光镜下观察,正常组视网膜内的NOS免疫阳性神经元较多,胞体较大,多分布于神经纤维层及内丛状层,胞体和近端树突染色较深,呈棕褐色。阳性区域面积为1696.20±151.90μm2。两组弱视组视网膜内均可见NOS免疫阳性神经元减少,胞体染色淡。其阳性细胞区域面积为:形觉剥夺组1072.36±101.67μm2,斜视组1173.34±123.77μm2。两组弱视组的NOS阳性区域面积分别与正常组比较,差异有显著统计学意义(P=0.000)。两组弱视组间NOS阳性区域面积比较,差异无统计学意义(P=0.211)。
2.3弱视眼视网膜中GABA阳性细胞面积变化在光镜下,正常组视网膜内的GABA免疫阳性神经元较多,细胞染色深呈棕褐色,胞体较大,多位于内核层、内丛状层和神经纤维层,高倍镜下,阳性细胞呈圆形或卵圆形,胞体及轴、树突内有免疫阳性物质,细胞核着色不明显。阳性区域面积为:2589.81±239.28μm2。两组弱视组视网膜内均可见GABA免疫阳性神经元减少,胞体染色淡,胞核皱缩。其阳性区域面积为:形觉剥夺组1939.41±105.76μm2,斜视组1894.45±155.45μm2。两组弱视组GABA阳性区域面积分别与正常组比较,差异有显著统计学意义(P=0.000)。两组弱视组间GABA阳性区域面积比较,差异无统计学意义(P=0.758,表2,图1、2)。
3 讨论
3.1NO在弱视眼视网膜中的表达及作用NO作为一种逆行信使加强兴奋性神经递质谷氨酸的释放,稳定突触前后的连接;能依据不同的突触活性发挥兴奋性或抑制性作用,选择性地增强被激活的神经通路,而抑制周围不被刺激的神经通路。近年来研究表明,NO可影响视觉的发育,参与视觉的形成、整合和传递,推测NO可影响弱视的形成[3]。而NO是在一氧化氮合酶(NOS)催化下由L-精氨酸分解为L-肌氨酸和NO而来的,只有那些能够合成NO的细胞才存在NOS,因此可以利用纯化的NOS制备抗体进行免疫组织化学染色,显示含有NOS的神经细胞,这种NOS阳性细胞的分布面积能够间接反映内源性NO水平[4]。
本研究采用免疫组化的方法检测两种病因弱视猫视网膜中NOS的表达。结果发现,两种病因弱视,关键期内的异常视觉经验均造成了视网膜中NO及NOS含量减少,与正常眼相比,差异有统计学意义。说明NOS阳性细胞数受到异常视觉经验的影响,推测NO可能在各种病因弱视的形成过程中起重要作用。目前发现,在大鼠、兔、猫、猴及人的视网膜中均存在NOS阳性神经细胞,主要分布于水平细胞、无长突细胞和神经节细胞[5]。有实验证明,NO可能是无长突细胞的神经递质之一。这些NOS阳性细胞与双极细胞、神经节细胞形成突触,在视网膜内与一些神经递质或神经肽共同参与视觉形成、局部整合、信息传递和调节神经节细胞的on/off感受野的功能[6]。可见NO可能参与了视网膜视觉冲动突触传导的过程,并通过调节一些神经递质的表达,影响视觉信号传导的强弱。可能的影响机制为:(1)NO水平的降低与视网膜细胞突触传递的兴奋性降低有关。已有研究表明,内源性NO调节神经元功能的主要生理作用是易化兴奋[7],大量的电生理研究已经证实异常的视觉经验导致弱视眼视觉冲动传递的抑制及兴奋性降低,推测这可能与内源性NO水平降低有一定的联系。(2)NO通过介导兴奋性氨基酸递质引起cGMP的变化而起作用[8]。谷氨酸为视网膜最常见的兴奋性神经递质,关键期内的异常视觉经验使视网膜失去清晰的光刺激,兴奋性刺激的减少使谷氨酸生成减少,通过反馈机制导致NOS水平下降,从而导致NO合成不足,不足以升高细胞内的环鸟苷酸(cGMP)的含量,以致于不能活化依赖cGMP的蛋白激酶,影响视觉的形成、整合和传递。(3)NOS阳性神经元废用性发育障碍[9]。异常的视觉经验使弱视眼相应输入层长期失去正常视信息的刺激,导致NOS神经元发育障碍,使得正常的突触联系的建立受阻,进一步使视冲动的传递受到抑制,视功能减弱从而形成了弱视。我们认为,异常视觉经验能够降低视网膜中NO的水平,推测NO参与了弱视眼视网膜内神经兴奋性的改变,NO水平的降低可能是弱视对视网膜影响机制中的一个重要环节。
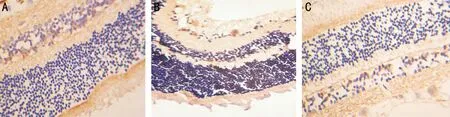
图1三组幼猫视网膜NOS免疫组化染色(×200)A:正常组:视网膜NOS免疫阳性神经元较多,胞体较大,多分布于神经纤维层及内丛状层,胞体和近端树突染色较深,呈棕褐色;B:形觉剥夺组:视网膜内NOS免疫阳性神经元减少,胞体染色淡;C:斜视组:视网膜内NOS免疫阳性神经元减少,胞体染色淡。
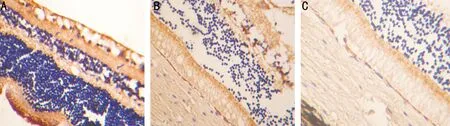
图2三组幼猫弱视眼视网膜GABA免疫组化染色(×200)A:正常组:视网膜GABA免疫阳性神经元较多,细胞染色深呈棕褐色,胞体较大,多位于内核层、内丛状层和神经纤维层,高倍镜下,阳性细胞呈圆形或卵圆形,胞体及轴、树突内有免疫阳性物质;B:形觉剥夺组:GABA免疫阳性神经元减少,胞体染色淡;C:斜视组:GABA免疫阳性神经元减少,胞体染色淡。
3.2GABA在弱视眼视网膜中的表达及作用GABA是谷氨酸在谷氨酸脱羧酶(GAD)作用下转化而成的,是一种重要的抑制性递质,其在中枢神经系统中广泛存在,对视觉发育起着重要作用。GABA抑制能力的减弱将直接影响猫视皮层神经元对视觉刺激的反应及对刺激反应的方向选择性[10]。Leventhal等[11]证实GABA、GABAA受体的激动剂可提高老年猴的视觉功能,而GABAA受体拮抗剂对年轻成年猴的视觉功能的抑制作用较强。提示视皮层中GABA抑制系统减弱很可能是导致老年猴视觉功能衰退的重要原因。此外,GABA受体可能还参与了双眼优势柱的转移[12]。除了抑制作用,GABA在发育早期神经元也可引起类似的去极化电位和兴奋性活动,其兴奋性作用主要是由谷氨酸介导的,通过升高胞内Ca2+浓度,从而对发育细胞产生兴奋性作用,此时GABA发挥了重要的神经营养性作用[13]。
GABA在视网膜主要分布于内核层内边缘的无长突细胞、内丛状层、神经节细胞和神经纤维层,其含量的变化可以反映视网膜的功能状态。本研究表明,剥夺正常视觉刺激12wk后,各组病因弱视眼视网膜GABA免疫阳性细胞染色淡,阳性面积减少,与正常眼相比,差异有统计学意义。提示GABA可能参与了视网膜神经元早期发育过程。有研究发现感光细胞-双极细胞-节细胞链使用兴奋性递质谷氨酸作为神经递质,而形成侧向链的水平细胞、无长突细胞等中间神经元使用甘氨酸或GABA作为神经递质[14]。猜测在幼猫视觉发育敏感期,异常的视觉经验使视网膜相应兴奋性神经元长期失去正常的视觉冲动,功能废用导致发育不良、胞体皱缩及数量减少,除向视中枢传导障碍外,与GABA能神经元形成的侧突冲动也减少,使它所支配的GABA能神经元发育障碍,如此形成退变循环,从而加速了弱视过程的发生。此外,GABA可能还介导视网膜神经突触的可塑性,长时程增强(long-term potentiation,LTP)和长时程抑制(long-term depression,LTD)是兴奋性谷氨酸能突触可塑性的两种重要形式。有研究发现[15],在新生海马LTD和LTP的产生是由GABAA受体所介导的,而Ca2+浓度的增加是GABA能突触可塑性的始动原因。我们认为,GABA水平对关键期内视网膜神经元的发育以及神经突触可塑性的改变起着重要作用。目前有关GABA在视网膜层面的研究较少,其在弱视形成过程中的具体神经机制还有待于进一步探讨。
弱视的发病是从视网膜到视皮质神经网络异常改变的复杂过程,视觉发育可塑性关键期内异常的视觉经验可能导致神经元突触分子水平的变化,NO及GABA的异常变化可能是其中的重要环节,这将为弱视发病机制的分子生物学研究提供重要基础。
1 von Noorden GK. The development of the art and science of strabismology outside North Ameriea:Part I.JAAPOS200l;5(2):65-69
2 Milleret C, Houzel JC. Visual interhemispheric transfer to areas 17 and 18 incats with convergent strabismus.EurJNeurosci2001;13(1):137-152
3 Yang Y, Ozawa H, Yuri K. Postnatal Development of NADPH-Diaphorase Activity in the Rat. The Role of Nitric Oxide in the Ontogeny of Arginine Vasopression and Oxytocin.EndocrJ2000;47(5):601-613
4 Bredt DS, Hwang PM, Synder SH.Localization of nitric oxide synthase indicating a neural role for nitric oxide. Nature 1990;347(6295):768-770
5 Zemel E, Lei B, Perlman I.NADPH diaphorase activity in the rabbit retina is modulated by glutamatergic pathways.JCompNeurol2001;431(1):28-38
6 Lau KC, So KF, Tay D. NADPH-diaphorase neurons in the retina of the hamster.JCompNeurol1994;350(4):550-558
7 Knott AB, Ella BW. Nitric Oxide in Health and Disease of the Nervous System. AntioxidRedoxSignal2009;11(3):541-553
8韩济生. 神经科学纲要. 北京:北京医科大学中国协合医科大学联合出版社 1993:399
9刘燕,张东杲.剥夺性弱视大鼠视中枢一氧化氮合酶阳性神经元分布与数量的变化.第四军医大学学报 1997;18(4):15
10 Eysel UT, Shevelev IA, Lazareva NA.Orientation tuning and receptive field structure in cat striate neurons during local blockade of intracortical inhibition.Neuroscience1998;84(1):25-36
11 Leventhal AG, Wang Y, Pu M. GABA and it sagonists improved visual cortical function in senescent monkeys.Science2003;300(5620):721-722
12 Heimel JA, Robin J, Hartman JM,etal. Screening mouse vision with intrinsic signal optical imaging.EurJNeurosci2007;25(3):795-804
13 Hales TG, Sanderson MJ, Charles AC. GABA Has Excitatory Actions on GnRH-Secreting Immortalized Hypothalamic (GT1-7) Neurons.Neuroendocrinology2008;59(3):297-308
14 Quesada A, Chmielewski C, Genis-Galvez JM. Immuno-histochemical localization of GABA in chameleon retina.CellBiolInt1996;20(6):395-400
15 Ben-Ari Y, Khazipov R, Leinekugel X. GABAA,NMDA and AMPA receptors: a developmentally regulated ’menage a trois’.TrendsNeurosci1997;20(11):523-529
Expression of nitric oxide and γ-aminobutyric acid in the retina of two kinds of amblyopia cats
Han-Min Wang1, Ao Rong2, Li-Juan Mo1, Qing-Song Li1, Xing-Ru Zhang1
Han-Min Wang. Department of Ophthalmology, Putuo Hospital, Shanghai University of Traditional Chinese Medicine, Shanghai 200062, China. langrisser_wang@163.com
2016-06-30Accepted:2016-09-29
•AIM: To study the role of nitric oxide (NO) and γ-aminobutyric acid (GABA) in the formation of amblyopia by establishing 2 different types of amblyopic models.
amblyopia; pattern visual evoked potentials; nitric oxide; γ-aminobutyric acid
1(200062)中国上海市,上海中医药大学附属普陀医院眼科;2(200065)中国上海市,上海同济大学附属同济医院眼科
王晗敏,毕业于同济大学医学院,硕士,主治医师,研究方向:眼表疾病。
王晗敏.langrisser_wang@163.com
2016-06-30
2016-09-29
Wang HM, Rong A, Mo LJ,etal. Expression of nitric oxide and γ-aminobutyric acid in the retina of two kinds of amblyopia cats.GuojiYankeZazhi(IntEyeSci) 2016;16(11):2006-2009
10.3980/j.issn.1672-5123.2016.11.06
猜你喜欢
杂志排行

国际眼科杂志的其它文章
- The effect of lowering target oxygen saturation on retinopathy of prematurity in neonatal respiratory distress syndrome
- 生物信息法推算青光眼小鼠视乳头及视网膜功能变化
- 不同切口超声乳化联合MI60型人工晶状体植入术对白内障患者视觉质量的影响
- 槲皮素对H2O2诱导人RPE细胞氧化应激损伤的保护作用
- Characteristics and visual outcomes of open globe injuries in children of amblyogenic age in Hong Kong
- 梅毒性眼病患者C-反应蛋白和血沉与白细胞联合检测的价值