金沟岭林场3种林型不同郁闭度林下灌草生物量1)
2016-11-07季蕾亢新刚郭韦韦汪晶张青
季蕾 亢新刚 郭韦韦 汪晶 张青
(北京林业大学,北京,100083)
金沟岭林场3种林型不同郁闭度林下灌草生物量1)
季蕾 亢新刚 郭韦韦 汪晶 张青
(北京林业大学,北京,100083)
以金沟岭林场云冷杉林、杨桦次生林和落叶松人工林为研究对象,分别设置郁闭度0.20、0.40、0.60、0.80、1.00样地,采用灌木生物量模型法和草本生物量平均木法,估算出不同林型不同郁闭度林下灌草生物量。结果表明:云冷杉林郁闭度为0.02时灌草生物量最大,郁闭度1.00时,灌草生物量最小,灌草生物量随林分郁闭度的增加而减小;杨桦次生林郁闭度为0.20时灌草生物量最大,郁闭度1.00时,灌草生物量最小,灌草生物量随林分郁闭度的增加而减小;落叶松人工林在林龄较小时,灌草生物量最小,母树林灌草生物量最大。不同林型在同一郁闭度林下灌草生物量差异显著。郁闭度为0.20和0.40时,杨桦次生林灌草生物量均大于云冷杉林;郁闭度为0.60时,灌草生物量由大到小的顺序为云冷杉林、杨桦次生林、落叶松人工林;郁闭度为0.80时,3种林型灌草生物量几乎相等;郁闭度为1.00时,落叶松人工林灌草生物量最大,且其灌木和草本生物量均大于其他两种林型。
云冷杉林;杨桦次生林;落叶松人工林;郁闭度;灌草生物量
With the spruce fir forest, polar-birch secondary forest and larch plantation in Jingouling Forest Farm, we, respectively, arranged the samples of different crown densities of 0.20, 0.40, 0.60, 0.80, and 1.00. T shrub biomass model and average herb method were used to estimate the understory biomass of different forests with different crown densities. When the crown density is 0.20, the understory biomass of spruce fir forest is maximum. When the crown density is 1.00, the understory biomass is minimum. The understory vegetation biomass is decreased with the increase of crown density. In poplar birch secondary forest, the understory biomass is maximum when the crown density is 0.20, and the understory biomass is minimum when the crown density is 1.00, and the understory vegetation biomass is also decreased with the increase of crown density. In different age of larch plantation, when the age is small, the understory biomass is minimum, and in the mother trees, the understory biomass is maximum. Under the same density, the biomass of different forest types is significantly different. When the crown density is 0.20 and 0.40, the understory biomass of poplar birch secondary are greater than that of spruce fir forest. When the crown density is 0.60, the descending order of understory biomass is spruce fir forest, poplar birch secondary forest, and larch plantation. When the crown density is 0.80, the understory biomass of three types of forest is almost equal. When the crown density is 1.00, the understory biomass of larch plantation is maximum, and its shrub biomass and herb biomass are greater than those of the other two types.
灌木层是森林生态系统的重要组成部分,它参与了森林生态系统的物质循环和能量的转化[1]。森林的林下微环境是不断变化着的,它对森林生态系统的稳定发展、演替过程以及物种的多样性起着重要的作用[2]。植物群落生物量是森林生态系统研究的重要内容之一,是研究森林物质生产和群落养分动态的基础[3]。生物量的测定成为生态系统结构和功能研究中不可缺少的一项基础性工作[4]。虽然林下灌草生物量在整个森林总生物量中所占的比例较小,但由于灌草层在维持森林群落的稳定性、森林物种的多样性以及水土保持效益上的重要作用,林下灌木层生物量的估算与研究对森林生态群落物质的循环与固定、积聚与消耗以及能量的转换具有相当重要的意义[5-13]。森林生态系统中林下植被的分布和生长特征受到林分乔木层特征的限制[2],而同一林型不同林分郁闭度或不同林型同一郁闭度下灌木和草本的生物量不同[14-15]。张胜伟等[16]通过研究桉树人工林与天然林林下植被生物量的比较中发现,桉树人工林林下植被生物量在不受人为以及其他因素的干扰下,均小于相同立地条件的天然林林下植被的生物量,而草本层生物量比例大于天然林草本层生物量的比例。文中以2015年野外调查的大量实测数据为基础,根据灌木生物量模型法和草本平均木法估算了不同郁闭度云冷杉林、杨桦次生林和不同年龄落叶松人工林下灌木和草本的生物量,为研究金沟岭林场不同森林类型不同郁闭度林分与林下灌草层生物量的关系提供了基础依据。森林碳储量与生物量存在一定的转换关系且对碳储量的研究不断受到关注,而本文为以后研究该地区的森林碳储量打下一定基础。
1 研究区概况
研究地区位于吉林省汪清县金沟岭林场,森林面积16 286 hm2。低山丘陵地貌,海拔300~1 200 m,坡度5°~25°。年平均气温约3.9 ℃;7月份气温最高,约22 ℃;年降水量600~700 mm。研究区主要乔木有冷杉(Abiesnephrolepis)、鱼鳞云杉(Piceajezoensis)、红皮云杉(Piceakoraiensis)、红松(Pinuskoraiensis)、白桦(Betulaplatyphylla)、枫桦(Betulacostata)等;灌木主要有毛榛(Corylusmandshurica)、暴马丁香(Syringaamurensis)、忍冬(Lonicerajaponica)、溲疏(Deutziascabra)等。主要地被植物为少数灌木和禾本科草类,如珍珠梅(Sorbariasorblia)、柳叶绣线菊(Spiraeasalicifolia)等。
2 研究方法
以天然云冷杉林、杨桦次生林和落叶松人工林为研究对象,设置5种郁闭度0.20(0.10~0.21)、0.40(0.30~0.41)、0.60(0.50~0.61)、0.80(0.70~0.81)和1.00(0.90~1.00),云冷杉林和杨桦次生林样地大小为20 m×20 m,每种郁闭度3块和17年生、38年生、52年生及母树林的落叶松人工林样地各4块,灌草样方设置在样地的中心及四角,灌木样方5 m×5 m,草本样方1 m×1 m。调查前记录样地所有特征因子,如郁闭度、坡向、坡度等因子。记录每株灌木的名称并测量每株灌木的地径(D,mm)或胸径、冠幅(C,C1长轴、C2短轴,m)、株高(H,m)、盖度等,调查草本样方中每个草本物种的高度或冠幅、株数和盖度,求出平均木的高度或冠幅,然后整株挖出每个草本物种的平均木,深度达根系所到之处,将根系土全部清除后再称量,将称量的植株装入信封带回实验室,在烘箱内105 ℃烘干得到每个物种草本的干质量。云冷杉林和杨桦次生林林分各自调查了25个灌木和25个草本样方,落叶松人工林中调查了20个灌木和20个草本样方,共调查了70个灌木和70个草本样方。3种林型不同郁闭度或林龄样地基本情况见表1。

表1 3种林型各郁闭度样地基本情况
注:—.表示无。
灌木生物量模型选取:所有林型灌木生物量计算均选用何列艳[17]研究该地区各灌木单一物种混合生物量模型,没有涉及的灌木物种,则根据灌木的形态—木型灌木和典型灌木[18],选用该地区乔木型灌木生物量模型或典型灌木生物量模型[17]。各灌木生物量模型见表2。
3 结果与分析
3.1 天然云冷杉林林下灌草生物量
3.1.1 不同郁闭度云冷杉林林下灌草生物量分布
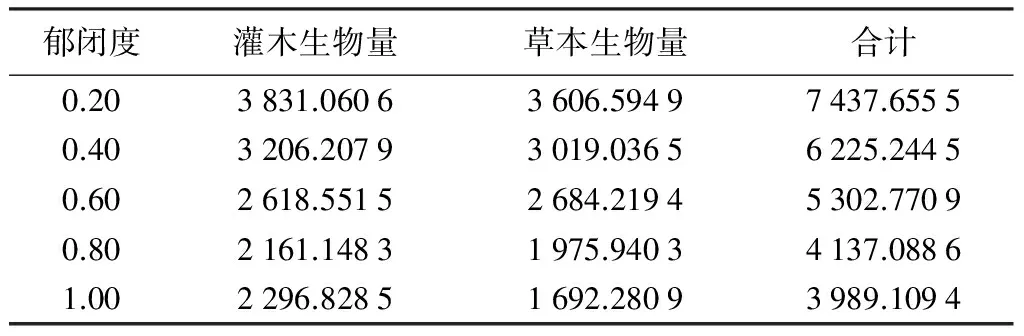
从表3可知,云冷杉林郁闭度为0.20时林下植被生物量最大,为7 437.655 5 kg·hm-2,因为,此时林下光照等条件较好,灌草营养空间较大。郁闭度1.00时,灌草生物量最小,为3 989.109 4 kg·hm-2,林分郁闭度高,林下灌草生长相对较弱,林下灌草生物量随林分郁闭度的增加而减小。灌木生物量在郁闭度0.20时最大,为3 831.060 6 kg·hm-2,在0.80时,灌木生物量最小,为2 161.148 3 kg·hm-2,由调查可知,这是因为在该郁闭度下,林下更新苗较多。除了郁闭度0.80以外,林下灌木生物量随林分郁闭度增加而减小。林下草本生物量也在郁闭度0.20时最大,为3 606.594 9 kg·hm-2,当郁闭度为1.00时,林下草本生物量最小,为1 692.280 9 kg·hm-2,整体上林下草本生物量也随林分郁闭度的增加而减小。灌木生物量和草本生物量在云冷杉林林下植被生物量组成中均占有相当大的比例。

表2 灌木生物量计算模型
注:D表示植株地径;H表示植株高;AC=πCICw/4表示植株冠面积,其中,CI、Cw分别表示冠幅的长短轴;VC=Ac×H,表示植株体积。
3.1.2 林下灌木生物量各物种分配特征
由表4可知,不同郁闭度云冷杉林林下各物种生物量分配不一致。林分郁闭度为0.20时,对灌木生物量贡献率最大的是毛榛,贡献率达22.31%,其次为暴马丁香,贡献率达18.41%,且花楷槭和青楷槭的贡献率也相对较高;郁闭度为0.40时,对灌木生物量贡献率最大是花楷槭,达23.77%,其次为青楷槭,贡献率达19.76%,再次,暴马丁香和毛榛的贡献率也相对较大;郁闭度为0.60时,灌木生物量组成主要是暴马丁香、花楷槭和青楷槭,其贡献率分别为26.57%、18.59%、16.88%,且从该郁闭度开始,茶藨子、楤木和野蔷薇开始渐逝;郁闭度为0.80时,对灌木生物量贡献最大的是忍冬,占总生物量组成比例为22.76%,绣线菊次之,贡献率达15.75%,且在该郁闭度下,刺五加、暴马丁香、花楷槭、毛榛生物量较为相似,分别约占10%;郁闭度为1.00时,忍冬占林下灌木生物量组成的主要部分,贡献率达38.25%,其次为绣线菊。总之,在郁闭度较小时,像毛榛、花楷槭和暴马丁香类的乔木型灌木对林下生物量的贡献率都很大,随着郁闭度的增大,耐阴树种在云冷杉林林下灌木生物量组成中占相当大的优势。
表3 不同郁闭度云冷杉林林下灌草生物量
kg·hm-2

表4 不同郁闭度云冷杉林林下各灌木生物量贡献率
3.2 杨桦次生林林下灌草生物量
3.2.1 不同郁闭度杨桦次生林林下灌草生物量分布
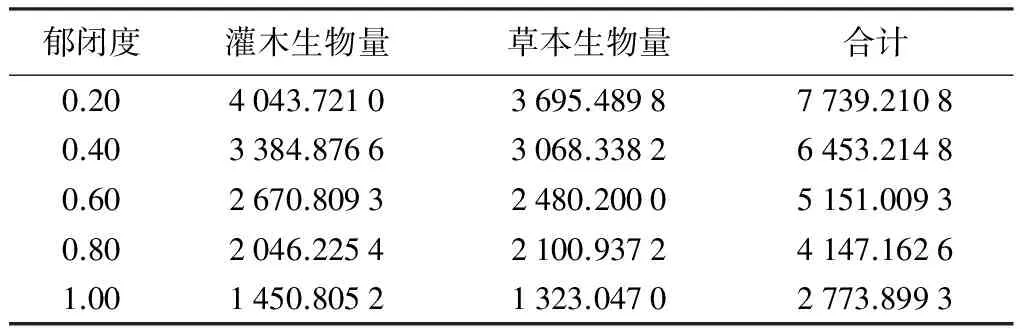
由表5可知,不同郁闭度杨桦次生林林下灌草生物量分配差异显著。郁闭度为0.20时,灌草生物量最大,为7 739.210 8 kg·hm-2;郁闭度1.00时,灌草生物量最小,为2 773.899 3 kg·hm-2。灌草生物量随林分郁闭度增加而减小。灌木生物量在郁闭度0.20时最大,为4 043.721 0 kg·hm-2;在1.00时,生物量最小,为1 450.805 2 kg·hm-2,由调查可知,因为在该郁闭度下,林内光照条件不好,下层木过于稀疏和矮小,林下灌木生物量也随林分郁闭度的增大而减小。草本生物量也在0.20时最大,为3 695.489 8 kg·hm-2;在1.00时,草本生物量最小,为1 323.047 0 kg·hm-2,整体上草本生物量随林分郁闭度的增加而减小。灌木生物量和草本生物量在杨桦次生林林下植被生物量组成中也均占有相当大的比例,草本生物量略低于灌木生物量。
表5 不同郁闭度杨桦次生林林下灌草生物量
kg·hm-2
3.2.2 灌木生物量各物种分配特征
由表6可知,不同郁闭度杨桦次生林林下各物种生物量分配不一致。郁闭度为0.20时,对灌木生物量贡献率最大的是绣线菊,贡献率达24.98%,其次为茶藨子,贡献率达19.03%,且暴马丁香和忍冬的贡献率也相对较高;郁闭度为0.40时,对灌木生物量贡献率最大的也是绣线菊,达23.84%,其次为暴马丁香和毛榛,贡献率分别为17.17%和16.04%;郁闭度为0.60时,对林下植被生物量组成贡献最大的也是绣线菊,贡献率为25.19%,暴马丁香、毛榛和溲疏生物量居于其次,贡献率分别达到16.26%、12.37%和15.23%;郁闭度为0.80时,对灌木生物量贡献最大的是花楷槭,为40.95%,暴马丁香次之,贡献率达30.92%,且在该郁闭度下,其他物种生物量比例相对较小;郁闭度为1.00时,忍冬占林下灌木生物量组成的主要部分,其贡献率达30.17%,毛榛,绣线菊和暴马丁香生物量也较高。总之,在郁闭度较小时,绣线菊的生物量占主导地位,像毛榛、花楷槭和暴马丁香类的乔木型灌木对林下生物量的贡献率相对次之,随着林分郁闭度增大,占林下植被生物量组成主导地位的物种也在发生着变化,林分郁闭度越大,耐阴树种或乔木型树种占林下植被生物量组成的比例越大。

表6 不同郁闭度杨桦次生林林下各物种生物量贡献率
3.3 落叶松人工林林下灌草生物量
3.3.1 不同林龄落叶松人工林林下灌草生物量分布
由表7可知,不同林龄落叶松人工林林下灌草生物量分配差异显著。当林龄较小时(17年生),灌草生物量最小,为219.896 6 kg·hm-2,且此时林下草本生物量大于灌木生物量,林下几乎没有灌木;母树林时,灌草生物量最大,为5 383.461 8 kg·hm-2。灌木生物量在母树林中最大,为3 400.585 7 kg·hm-2,在17年生时,灌木生物量最小,为12.792 2 kg·hm-2。由调查可知,因为在该林龄时,林分株数郁闭度较大,林内枯枝落叶全为针叶,难分解[19],林下腐殖质较少,几乎无灌木生存。林下灌木生物量随林龄的增大而增加。草本生物量也在17年生时最小,为207.104 4 kg·hm-2,在母树林中,草本生物量值最大,为1 982.876 1 kg·hm-2,除了17年生林分外,林下草本生物量随林龄的变化差异不显著。灌木和草本生物量在落叶松人工林林下植被生物量组成中也均占有相当大的比例,在17年生和38年生的林分中,草本生物量均大于灌木生物量,草本生物量占总生物量的比例分别为94.18%和68.18%,在较成熟的林分中,林下草本生物量均小于灌木生物量,灌木生物量占总生物量的比例分别为55.26%和63.17%。
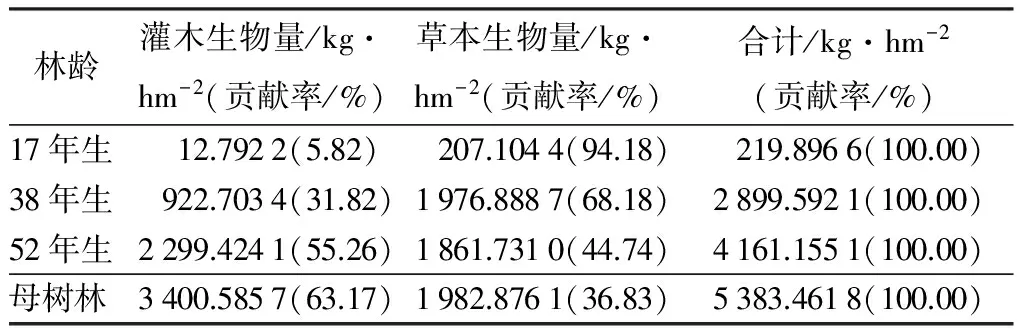
表7 落叶松人工林不同林龄林下灌草生物量
3.3.2 林下灌木生物量各物种分配特征
由表8可知,不同林龄落叶松人工林林下各物种生物量分配不一致。17年生时,林下灌木稀少,仅有忍冬和珍珠梅,贡献率分别为53.11%和46.89%,灌木生物量在此时最小;38年生时,绣线菊对灌木生物量贡献率最大,达93.15%,在该林龄林分中,灌木开始增多,因此灌木生物量骤增,但除大量涌现绣线菊外,其他物种还相对较少;52年生时,对林下植被生物量组成贡献率最大的也是绣线菊,为38.76%,其次为忍冬,贡献率为20.37%,同时花楷槭、毛榛和茶藨子对该林龄下灌木生物量贡献也相对较大,且此时灌木也开始丰富,除了绣线菊外,其他物种生物量所占的比例也开始显著;在母树林中,灌木生物量贡献率最大的是暴马丁香,占总生物量组成的比例为48.19%,花楷槭次之,贡献率达37.95%,暴马丁香和花楷槭是该林龄下灌木生物量的主要部分,其他物种所占的比例相对于52年生的林分,均略有降低。且在母树林中,绣线菊所占的比例相对于前3种林龄的林分,其比例下降的相当明显。总之,在林龄较小时,林下灌木种类和数量均较少,因此,灌木生物量也少。当林龄较大时,灌木生物量出现骤增的现象,且在38年生和52年生的林分中绣线菊都占有绝对的主导优势。而到了母树林时,像花楷槭和暴马丁香类的乔木型灌木对林下生物量的贡献率占据了绝对优势。随林龄增加,占林下生物量组成主导地位的物种也在发生着变化,落叶松林龄愈大,耐阴树种或乔木型树种占林下植被生物量组成的比例越大。
3.4 3种林型林下植被生物量比较分析
由表9可知,郁闭度为0.20和0.40时,杨桦次生林林下灌草总生物量、灌木生物量、草本生物量均大于云冷杉林的,两林型草本生物量近似;郁闭度为0.60时,灌草生物量由大到小的顺序为云冷杉林、杨桦次生林、落叶松人工林,但灌木生物量由大到小的顺序为杨桦林、云冷杉林、落叶松人工林,草本生物量由大到小的顺序是云冷杉林、杨桦林、落叶松人工林;郁闭度为0.80时,3种林型林下灌草生物量几乎相等,落叶松林下灌木生物量最大,云冷杉林灌木生物量略大于杨桦林,而草本生物量由大到小的顺序为杨桦林、云冷杉林、落叶松林;郁闭度为1.00时,人工落叶松母树林灌草生物量最大,云冷杉林灌草生物量大于杨桦林,落叶松灌木和草本生物量均大于其他两种林型,云冷杉林灌木和草本的生物量又均大于杨桦林。
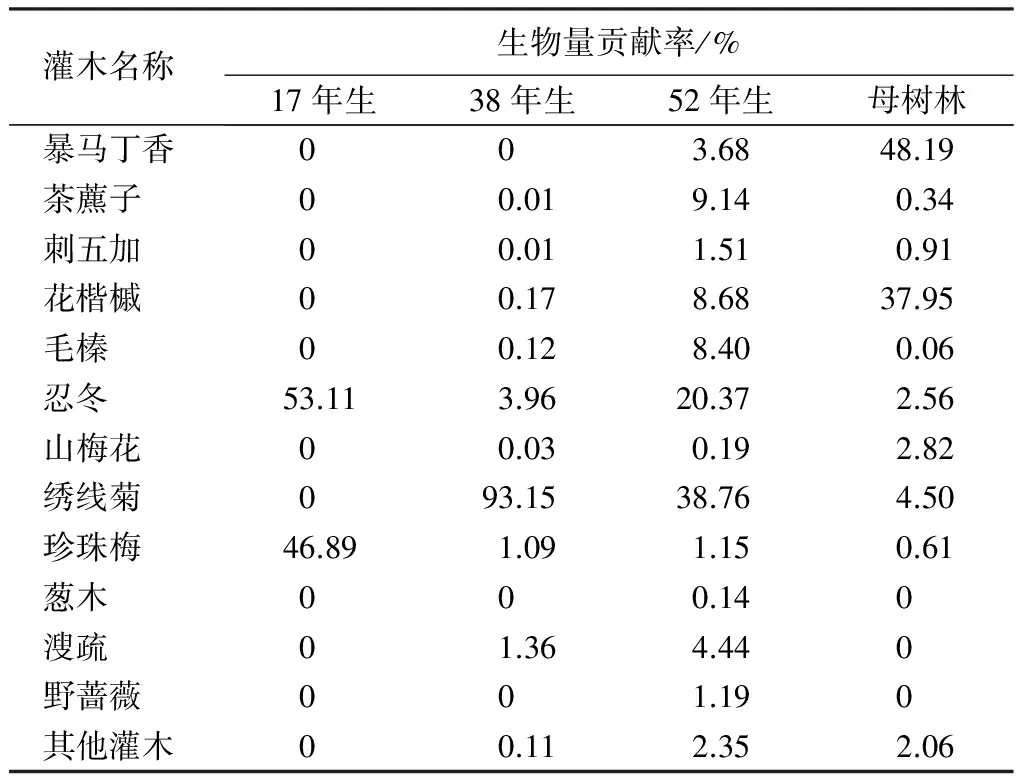
表8 落叶松人工林不同林龄林下各物种生物量贡献率
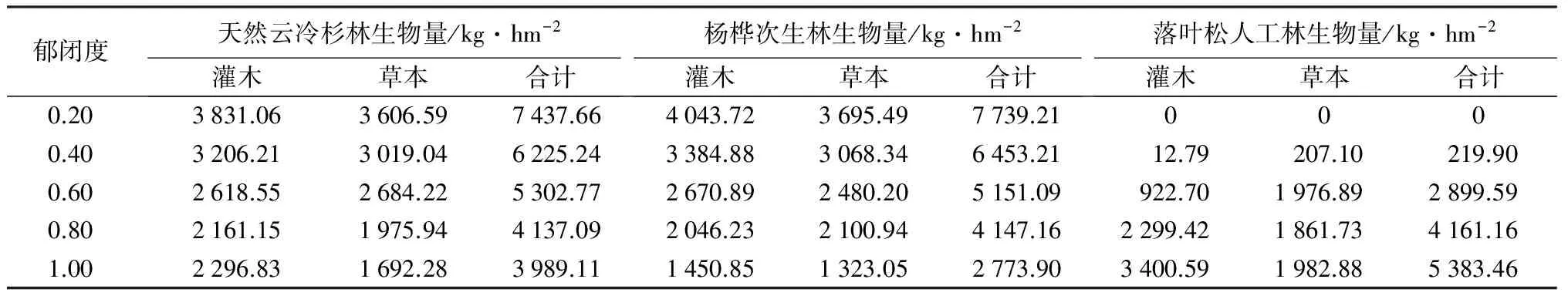
表9 3种林型不同郁闭度林下灌草生物量
4 结论与讨论
为了研究林下植被的物质结构特征,揭示不同森林类型或同一森林类型不同郁闭度的结构与功能的关系,文中选用了金沟岭林场分布较广的10种灌木生物量模型和草本平均木法,将其应用到该林区3种典型林型不同郁闭度林下灌木中。结果表明,天然云冷杉林不同郁闭度林下灌草生物量差异显著。云冷杉林郁闭度为0.20时林下植被的生物量最大,郁闭度为1.00时,林下灌草生物量最小。整体上,林下灌木草本生物量随着林分郁闭度的增加呈现逐渐减小的趋势;杨桦次生林不同郁闭度林下灌草生物量分配差异显著。郁闭度为0.20时,灌草生物量最大,郁闭度为1.00时,灌草生物量最小,林下灌草生物量随林分郁闭度的增加而减小;不同林龄落叶松人工林林下灌草生物量分配差异显著。当林龄较小时,林下灌草生物量最小,且此时林下草本生物量大于林下灌木生物量,林下几乎没有灌木,由调查可知,这是因为在该林龄时,林分郁闭度较大,林内枯枝落叶全为针叶,不易于分解[19],林下腐殖质较少,几乎无灌木生存;母树林灌草生物量最大。
不同林型在同一郁闭度下林下灌草生物量差异显著。郁闭度为0.20和0.40时,杨桦次生林下灌草生物量均大于云冷杉林,杨桦次生林下灌木和草本生物量也均大于云冷杉林,两林型草本生物量近似;郁闭度为0.60时,灌草生物量由大到小的顺序为云冷杉林、杨桦次生林、落叶松人工林,但杨桦林灌木生物量略大于云冷杉林,云冷杉林又大于落叶松人工林,而云冷杉林草本生物量大于杨桦林;郁闭度为0.80时,3种林型林下灌草生物量几乎相等,落叶松林下灌木生物量最大,云冷杉林次之,而草本生物量由大到小的顺序为杨桦林、云冷杉林、落叶松林;郁闭度为1.00时,落叶松人工林灌草生物量最大,云冷杉林大于杨桦林,落叶松灌木和草本生物量均大于其他两种林型,云冷杉林灌木和草本生物量又均大于杨桦林。在郁闭度较大的林分中,林下灌木生物量研究结果与前人研究一致[14-15]。
金沟岭林场天然森林结构较为复杂,林下灌草物种丰富,不同林型或不同郁闭度林下灌草分配不同。杨昆等[20]研究表明,林分特征因子光照条件影响着森林下层木的光照,从而影响着林下灌草的生物量。刘磊等[21]通过研究认为不同森林类型和同一森林类型在不同的发育阶段林下灌草生物量的差异是显著的,人工林林分的冠层结构和林分郁闭度影响着森林下层木的组成和林下植被生物量的格局,文中研究结果与其一致。3种林型林下植被生物量均以灌木生物量为主,但草本生物量也占有相当大的比例。3种林型同一郁闭度林下或同一林型不同郁闭度或林龄下灌草生物量具有一定的差异。文中是在尽量控制其他林分特征因子一致的情况下研究不同郁闭度林下灌草生物量的,对于该地区近原始林不同郁闭度林下灌草生物量以及乔木层树种组成、林内微环境和灌木树种组成与灌草生物量的关系均有待进一步研究。
[1] 潘辉,洪伟,陈国荣,等.相思人工林生物量分配格局与林下植被多样性分析[J].福建林业科技,2008,35(1):6-10.
[2] 何艺玲,傅懋毅.人工林林下植被的研究现状[J].林业科学研究,2002,15(6):727-733.
[3] 梁妮,王卫斌,倪金碧,等.西南桦纯林与西南桦+肉桂混交林幼林期的生物量研究[J].西部林业科学,2007,36(2):44-49.
[4] 赵振勇,王让会,张慧芝,等.天山南麓山前平原柽柳灌丛地上生物量[J].应用生态学报,2006,17(9):1557-1562.
[5] 郑海富.林下灌木生物量方程的验证和生物量分布格局研究[D].哈尔滨:东北林业大学,2010.
[6] 项文化,田大伦,闫文德.森林生物量与生产力研究综述[J].中南林业调查规划,2003,22(3):57-60.
[7] 潘攀,牟长城,孙志虎.长白落叶松人工林灌丛生物量的调查与分析[J].东北林业大学学报,2007,35(4):1-2,6.
[8] 赵蓓,郭泉水,牛树奎,等.大岗山林区几种常见灌木生物量估算与分析[J].东北林业大学学报,2012,40(9):28-33.
[9] FANG J Y, WANG Z M. Forest biomass estimation at regional and global levels, with special reference to China’s forest biomass[J]. Ecological Research,2001,16(3):587-592.
[10] PATON D, NUNEZ J, BAO D, et al. Forage biomass of 22 shrub species from Monfragüe Natural Park (SW Spain) assessed by log-log regression models[J]. Journal of Arid Environments,2002,52(2):223-231.
[11] BUECH R R, RUGG D J. Biomass relations of shrub components and their generality[J]. Forest Ecology and Management,1989,26(4):257-264.
[12] 曾慧卿,刘琪璟,冯宗炜,等.红壤丘陵区林下灌木生物量估算模型的建立及其应用[J].应用生态学报,2007,18(10):2185-2190.
[13] 潘攀,牟长城,孙志虎.长白落叶松人工林灌丛生物量的调查与分析[J].东北林业大学学报,2007,35(4):1-2.
[14] 何列艳,亢新刚,范小莉,等.长白山区林下主要灌木生物量估算与分析[J].南京林业大学学报(自然科学版),2011,35(5):45-50.
[15] 范小莉.长白山地区云冷杉林和近原始林林下灌草生物量预测模型的研究[D].北京:北京林业大学,2011.
[16] 张胜伟,赵一鹤,侯明明,等.桉树人工林与天然林林下植被生物量比较研究[J].资源开发与市场,2008,24(5):385-387.
[17] 何列艳.长白山过伐林区杨桦次生林与落叶松人工林林下灌草多样性和生物量研究[D].北京:北京林业大学,2011.
[18] 贺金生,王其兵,胡东.长江三峡地区典型灌丛的生物量及其再生能力[J].植物生态学报,1997,21(6):512-520.
[19] HEMINGWAY R W, PETER E L. Plant polyphenols[M]. New York: Springer,2013.
[20] 杨昆,管东生,周春华.潭江流域森林碳储量及其动态变化[J].应用生态学报,2006,17(9):1579-1582.
[21] 刘磊,温远光,卢立华,等.不同林龄杉木人工林林下植物组成及其生物量变化[J].广西科学,2007,14(2):172-176.
Biomass of Shrub and Herb under Three Forest Types with Different Canopy Densities in Jingouling Forest Farm//
Ji Lei, Kang Xingang, Guo Weiwei, Wang Jing, Zhang Qing
(Beijing Forestry University, Beijing 100083, P. R. China)//Journal of Northeast Forestry University,2016,44(9):29-33,39.
Natural spruce fir forest; Polar-Birch secondary forest; Larch plantation; Canopy density; Biomass of shrub and herb
1)国家林业局“948”项目(2013-4-66)、“十二五”国家科技支撑计划项目(2012BAD22B02-3)。
季蕾,女,1990年2月生,北京林业大学林学院,硕士研究生。E-mail:645908709@qq.com。
亢新刚,北京林业大学林学院,教授。E-mail:xingangk@163.com。
2016年1月24日。
S757
责任编辑:任 俐。
