基于内部优先的麻疯树构型可塑性的结构限制研究
2016-09-09何亚平费世民王乐辉陈秀明孟长来
何亚平,费世民,王乐辉,陈秀明,孟长来
(四川省林业科学研究院,四川 成都 610081)
·研究报告·
基于内部优先的麻疯树构型可塑性的结构限制研究
何亚平,费世民*,王乐辉,陈秀明,孟长来
(四川省林业科学研究院,四川成都 610081)
适应假说能解释木本植物构型对生境资源斑块的适应性,但树体发育早期结构对后期发育结构存在限制,这种看法还未受到普遍关注。本文研究了干热河谷灌木粮油能源麻疯树的树体构型对6种生境资源斑块的适应性响应,并分析了这种响应的结构限制性。麻疯树一级枝与二级枝数量对生境响应异常明显,个体分支率2∶1、分支率3∶2、总体分支率、枝径比2∶1、枝径比3∶2、总体枝径比、枝长比2∶1、枝长比3∶2、总体枝长比都有极为显著的差异表现。相关分析表明,分支率2∶1、枝径比2∶1、枝长比2∶1都与各类分支格局参数在不同生境间呈现显著正相关、负相关与无关关系,分支率2∶1副作用明显,而枝径比、枝长比的正作用相对明显。早期树体这种结构限制可能归因于分支数量与质量的权衡与正偶联。麻疯树构型对不同生境资源斑块适应是通过分支数量调控个体内资源配置格局与正负偶联建立的,具有复杂性。
麻疯树;构型;内部优先;可塑性;结构限制
植物大多数是依赖于光截取与光合作用来获得能量供应的固定有机体。陆地植物发育了庞大的构型来展示光合叶片,且植株很长寿,在整个生命史中存活(Moulia et al.,2006)。植物构件伸展与形态可塑性类似于动物觅食行为(Hutching&de Kroon,1994;董鸣,1996),绿色植物趋水、趋肥、趋光性是可塑性与生境适应的生理机制。很多研究探讨了克隆植物形成的生境适应性,即分株对水肥光照的适宜性而形成分株富集性与可塑性(董鸣,1996;董鸣等,2011)。木本植物构型是固定遗传生长方案表达的产物(陈波等,2002),种内可塑性受到生态因素的调节(何维明,董鸣,2001;苏建荣等,2005;孙书存,陈灵芝,1999a;袁秀英等,2009;胡喜生等,2006),特定植物在特定生境中的构型形态及其可塑性是进化限制与生境适应(董鸣,1996)的综合结果。种内生境资源斑块间构型分析主要是基于分支格局的生境适应性,这种调查分析国内外报道案例为数不多(Halle,1978;Halle et al.,1978;Bell& Tomlinson,1980;Whitney,1976;孙书存&陈灵芝,1999a,1999b;李亚莉等,2007;朱选伟等,2001;卢康宁&张怀清,2010),尽管有一些试验研究(何维明,董鸣,2001;苏建荣等,2005;袁秀英等,2009)。
麻疯树构型的动态变化及其优化参数受到研究(蔡小虎等,2012),树体早期发育结构对后期结构的限制作用很少受到关注。木本植物构型度量技术,提出并试验相关的计量概念与方法,如分支率、枝径、枝长、分支角度等,用外部优先枝序确定方法来测定不同枝级的分支格局的Strahler法(McMahon &Kronauer,1976)。实际上,木本植物构型研究枝序确定有两种方法(卢康宁&张怀清,2010),另外一种是内部发育起始的内部优先枝序确定方法(何亚平等,2010;卢康宁等,2011),内部优先枝序确定树体结构的研究案例极少。内部优先的枝序确定方法利于认识土壤生境而不是大气生境对构型发育的生态限制作用,树体发育早期面临的生境在基部分支格局中能得到体现;同时,内部优先的枝序确定更能分析早期起源构件间的相互作用效果。整个树体构型在光照环境中的可塑性在个体内不同遮阴程度构件分支格局上也有所表现,并共同塑造树体骨架与分支格局。朱万泽等(2007)提出了萌生分支的结构限制假说,来表明在结构限制存在时,富集水分和无机盐输出受到结构的限制而诱发萌生分支进而增加了树体构型的复杂性。从和谐思想看,早期发生枝条具有母枝决定性,对后发生枝条和构型建成影响是必然的;后发枝条对早期构型分支影响也是显著。从木本粮油树木的幼树建成看,早期发育构型对晚期发育构件存在影响,构型自身对构型发育的限制作用在构型分析中也不容忽视。
灌木树种基部发育枝条对后续分支影响较为明显,往往一级枝数量与主杆有无直接影响着树体高度与外部形态。木本粮油树种树体管理中也涉及到诸多树体构型,如自然开心形、主杆明显形等。麻疯树是一种生物柴油树种,广泛地适生于南北纬32度之间的热带地区,我国西南地区干热河谷为集中分布区与多度中心(向准等,2008)。麻疯树能源林基地的微地形异质性较为强烈,塑造了生境资源异质性斑块(尤其是水分),生殖物候差异大(何亚平等,2008),在落叶物候上形成了明显斑块,并作为基地内微观产量格局划分的重要依据(何亚平等,未发表资料)。麻疯树高产稳产壮枝培育离不开优化的构型分支,尤其是一级枝数量与格局的培育,初步用构件生长分析法研究了群体分支与个体构型对生境肥沃性的适应性(费世民等,2009;何亚平等,2010)。实践中,麻疯树构型分支特性认识不足,不同生境资源异质性斑块对树体构型作用表现认识并不清楚,限制了树体管理与构型优化技术的研发与运用。
高产稳产所依赖的壮枝新梢培育依赖于粗壮母枝与疏松开敞的树体结构,这种树体结构的发育需要肥沃的生境资源与精细的人工优化管理(何亚平等,2010)。麻疯树先发育构型分支对后续发育新梢存在限制作用或促进作用,影响了树体构型发育动态规律的认识。基于理性研究与生产实践的重要性,采用内部优先的枝序确定方法,对攀枝花市马店河5a麻疯树基地6类生境资源斑块下树体构型进行研究,探讨以下两个问题:1)不同生境下麻疯树树体构型的可塑性;2)基于不同生境的麻疯树构型发育的结构限制。综合分析麻疯树构型对不同生境资源斑块的响应特点,并分析早期发枝对晚期发枝的结构限制性,丰富木本植物构型研究,并为麻疯树树体管理提供理论依据与技术支撑。
1 材料与方法
1.1样地概况
调查样地位于攀枝花市仁和区大龙潭乡马店河,海拔940 m,经度为101°51′601″E,维度为26° 28′132″N。试验区域为西南山地金沙江干热河谷攀枝花段,处于元谋干热河谷向雅砻江流域干暖河谷过渡地带,在宽谷江边为稀树灌木草丛,峡谷地带为灌木林,宽阔坝塬直线坡面为扭黄茅草丛。该区域自然地理概况、人为活动与林业生态工程现状参考文献(张旭东等,2006)。
调查样地位于金沙江干流中山丘陵区,坡度在5°~45°度之间,坡位为中坡,坡向南,土壤为山地红壤,粘粒紧实,坡积土,母岩为砂岩。马店河人工林自然植被为稀树灌木草丛,灌木树种有台湾相思、蜜柚子、麻疯树,造林后以麻疯树为主体;草本植物主要是双花草、扭黄茅,在双花草草丛富集区域,雨季形成积水湿地,一般位于凹坡与陡峭山崖下方的厚土层缓平堆积区。
马店河麻疯树人工林为2005年栽植,栽植规模25.3 hm2,栽植坑大小为40 cm×40 cm,栽植当年施底肥,2006年~2009年在下部进行施肥试验或抚育性施肥。抚育管理措施是除草做防火带,禁止人为放牧,但放牧难以避免,存在明显的土壤干扰,形成斑块状的积水地,并使得树体存在明显的擦痕,存在一定数量的断枝、枯死枝。
1.2调查方法
2010年11月份对马店河基地从下端到顶端部分不同立地类型的麻疯树林分进行调查,样地面积10 m×10 m,对样地中每一株麻疯树个体测定地径、树高、冠幅长与宽,并以主干为中心,划分一级枝,以每一个一级枝为单位,调查一级枝的枝径、枝长、分支角度,统计二级枝、三级枝数量,选择平均枝作为标准枝,来测定枝径、枝长,分支角度用多种情形下角度的平均值估测值来代替。共计调查样地数量为39个。
依照实际情况,将调查立地划分为六类,第一类为农林复合工程,密度较低,进行了玉米、番薯套种,混栽桂圆、芒果、大青枣,且有砌石工程保持水土,共计3个样地;第二类为混交林,栽植麻疯树时为灌木林,栽植后灌木林与麻疯树同时发育,共计5个样地;第三类为上凸坡,位于坡体上部,水文上形成了源地,是水肥流失的地带,4块样地;第四类为缓斜坡地,位于坡体中上部,无法汇集水肥,水肥高于上凸坡,但低于缓平坡,样地共计11个;第五类为凹坡侧面与大块缓平坡,水肥路过,能形成水肥汇集区与吸收区,样地共计10个;第六类为凹坡或平地、洼地积水区,样地共计4个。
1.3分支格局计算方法
麻疯树分支格局调查是采用了内部优先法,主干分支为一级枝,一级枝上分支为二级枝,二级枝上分支为三级枝,三级枝上分支为四级枝。由于该基地麻疯树四级枝并不是所有枝条都能分支,发生数量少,调查只进行到三级枝。
(1)分支率计算
本文用外部优先枝序确定方法来测定不同枝级的分支格局的Strahler法(McMahon&Kronauer,1976)确立的分支率(Whitney在1976年提出,经Steingraceber和Waller1986年修改)计算公式(孙书存和陈灵芝,1999;卢怀青等,2011),对其进行相应的修改(将原来公式中外部的改为内部的):
总体分支率Rb=(NT-N1)/(NT-NH),其中NT为所有枝级中枝条的总数,N1为一级枝数量,NH为末级枝的枝条数量;
逐步分支率为高一枝级枝条数量与低一枝级枝条数量之比(Ri+1∶i):
Ri+1∶i=Ni+1∶Ni,其中Ni为第i级的枝条总数,Ni+1为第i+1级的枝条总数。
(2)枝径比计算公式
木本树体分支时,分支大小总是小于母枝。枝径比、枝长比反映了枝条分支时产生的枝径与长度衰退程度。依照分支率计算公式,编制一个枝径比、枝长比计算公式:
总体枝径比(Rd):Rd=(DT-D1)/(DTDH),其中DT为各枝级标准枝枝条枝径的总和,D1为一级枝枝径,DH为末级枝的枝径;
枝径比 (Rdi+1∶i):Rdi+1∶i=Ndi+1∶Ni,其中Ndi为第i级枝平均枝径,Ndi+1为第i+1级枝的平均枝径;
(3)枝长比计算公式
总体枝长比(Rl):Rl=(LT-L1)/(LT-LH),其中LT为各枝级标准枝枝条枝长的总和,L1为一级枝枝长,LH为末级枝的枝长;
枝长比(Rli+1∶i):Rli+1∶i=Nli+1∶Ni,其中Nli为第i级枝平均枝长,Nli+1为第i+1级枝的平均枝长;
1.4 数据统计
将数据录入计算机,划分生境类型,依照类型来汇集数据,用SPSS for Windows ANOVA分析差异;用Pearson Correlations分析相关性。
2 结果与分析
2.1不同生境构型分支的基本特点
枝长、枝径、分支率是麻疯树树体构型分支格局的基本参数,其中分支角度是主干与分支夹角。马店河6种生境麻疯树树体分支基本参数如表1所示。从一级枝到二级枝,分支角度从13.52°~22.04°增加到31.71°~38.46°,但二级枝与三级枝之间角度相似(31.67°~37.23°)。除了分支角度外,这些基本参数在缓斜坡、农林复合工程、积水坡面相对较大,在天然混交林与干燥凸坡则相对较小,陡峭斜坡基本居中,方差分析表明,不同生境斑块间,各类参数差异均达到极显著水平(p值均为0.000)。
6种生境麻疯树构型分支率、枝径比与枝长比如表2所示,统计分析后,各类生境斑块间差异极其显著(p值均为0.000)。麻疯树分支率2∶1农林复合与天然混交林较高,分支率3∶2与总体分支率则缓斜坡也相对较高,但3种分支率干燥凸坡都最低。枝径比缓斜坡、积水坡面相对较高,农林复合、天然混交林、干燥凸坡样地相对较低。枝长比则缓斜坡、积水坡面明显较高,干燥凸坡相对最低,其余居中。总体上看,缓斜坡、积水坡面样地枝径比、枝长比相对较高,缓斜坡、农林复合与天然混交林分支率相对较高,干燥凸坡所有参数都较低(枝径比3∶2除外)。

表1 马店河6种生境麻疯树树体分支基本参数Tab.1 Base parameters of tree branching in Jatropha curcas L.under six habitats in Madianhe base

表2 马店河6种生境麻疯树构型分支率、枝径比与枝长比Tab.2 Bifurcation ratio,branch diameter ratio and branch length ratio in Jatropha curcas L.under six habitats in Madianhe base
2.2不同生境构型参数相关分析
栽植后,麻疯树个体在存活与发育过程中,先产生一级枝,进而产生二级枝、三级枝。二级枝分支格局可能会影响到三级枝的分支格局,如二级枝分支率大,分支数量相对较多,枝条相对较小,则三级枝分支率、枝径比和枝长比可能会降低。分支率2∶1、枝径比2∶1与枝长比2∶1对分支率、枝径比与枝长比3∶2可能会产生3种作用,一种是毫无关系,无相关性;一种是促进作用,呈现正相关;一种是抑制作用,呈现负相关。通过Pearson相关分析,6种生境中麻疯树分支格局相关系数(与显著性P值)矩阵中能找到这3种关系,黑色加粗表示显著负相关,黑色没有加粗表示毫无关系,黑色加粗斜体为显著正相关。这种复杂的关系在表4天然混交林样地分支格局相关分析中可以反映,如分支率2∶1增加时,分支率3∶2不变,总体分支率增加,分别表现了无关与正增长关系,但枝径比2∶1、枝径比3∶2、总体枝径比、枝长比2∶1、枝长比3∶2、枝长比3∶1随之呈现降低性显著负相关。一个逐步分支参数对总体参数有促进作用,但却降低了另外一些类型的参数的值,形成了早期分支结构对后续分支结构的限制或抑制。
分支率2∶1与分支率3∶2的关系存在明显的生境间差异,除了上凸坡灌草丛样地呈现显著负相关外,其余样地均呈现无关关系。分支率2∶1与总体枝径比在所有样地均呈现显著负相关关系,但与枝径比2∶1、枝径比3∶2仅在农林复合工程与积水坡面呈现无关关系,在天然混交林、上凸坡样地、陡斜坡样地与缓坡地都呈现显著负相关关系(上凸坡样地枝径比3∶2为无关关系)。分支率2∶1与枝长比关系存在明显的生境间差异,农林复合工程、上凸坡样地、陡斜坡样地、积水坡面样地均呈现无关关系,但在缓坡地与天然混交林样地呈现显著负相关。二级枝分支比例对枝径比、枝长比的影响存在明显的样地一致性,表明这种早期结构限制实际上作用还受到样地生境资源状况的调节。
枝径比越大,表示分支枝径较大,表示分支枝条粗壮;枝长比也是类似的;早期枝径枝长比可能会影响后期枝条大小,反映分支退化速度与树体衰退速度。枝径比2∶1与分支率无相关(农林复合工程,天然混交林与缓坡地分支率2∶1例外);上凸坡、陡斜坡、积水地(分支率2∶1例外)全部相关,表明除了分支率2∶1(负相关)外,都随着枝径比2∶1增加而显著增大。枝径比2∶1与枝径比3∶2、总体枝径比、枝长比2∶1、枝长比3∶2、总体枝长比之间呈现两种相关性农林复合样地、天然混交林(枝径比3∶2例外)、陡斜坡样地(枝径比3∶2例外)、积水坡面,除了枝长比3∶2外都显著相关;上凸坡、缓坡地除了枝径比3∶2外,都呈现显著的相关关系。显著性结果表明,枝径比3∶2与枝径比2∶1增加而降低(抑制作用,仅在农林复合与积水坡面),其余均随着增加而递增,呈现促进作用(如表3至表8所示)。统计分析表明,枝长比2∶1与分支率2∶1、分支率3∶2、总体分支率、枝径比2∶1、枝径比3∶2、总体枝径比、枝径比3∶2、总体枝径比之间呈现促进、抑制与无关3种关系,在不同样地存在复杂的矩阵格局。产生新梢分支的相对大小直接影响了新分支的大小,进而限制或促进了新分支的发育,枝径比、枝长比在生境之间一致性关系并不明显,可能树体内部因素,如萌生发生、外界干扰、生殖输出等也都有所影响。

表3 农林复合工程样地麻疯树分支格局参数之间的相关分析Tab.3 The correlation analysis between bifurcation pattern of Jatropha curcas in agr-forestry plots
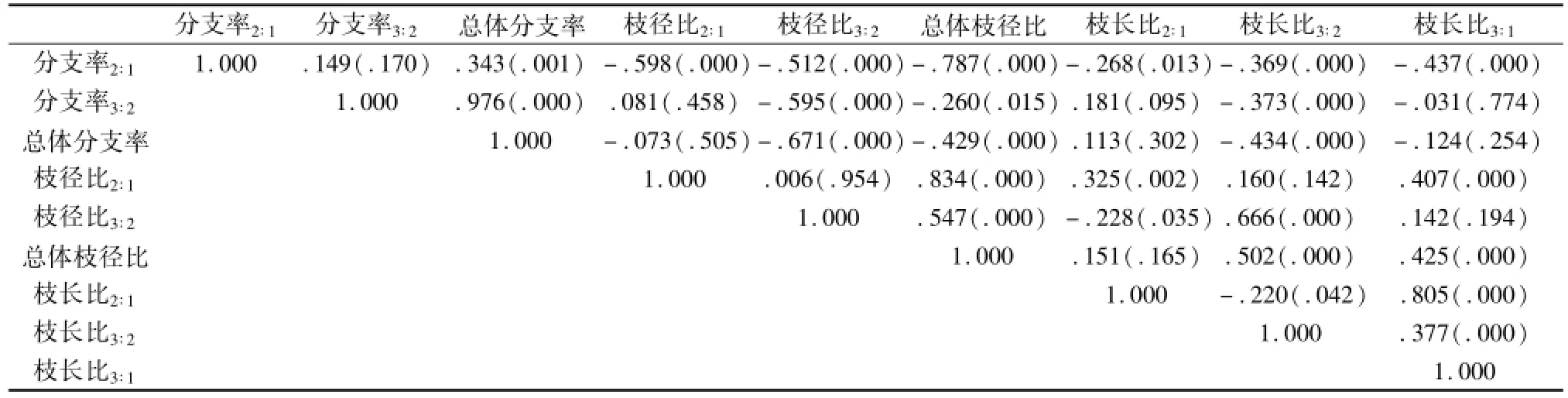
表4 天然混交林样地麻疯树分支格局参数之间的相关分析Tab.4 The correlation analysis between bifurcation pattern of Jatropha curcas in naturalmixed forest plots

表5 上凸坡样地麻疯树分支格局参数之间的相关分析Tab.5 The correlation analysis between bifurcation pattern of Jatropha curcas in upper convex slope plots

表6 陡斜坡样地麻疯树分支格局参数之间的相关分析Tab.6 The correlation analysis between bifurcation pattern of Jatropha curcas in steep slope plots

表8 积水坡面样地麻疯树分支格局参数之间的相关分析Tab.8 The correlation analysis between bifurcation pattern of Jatropha curcas inmoisture-abundant slope plots
3 讨论与结论
木本植物构型存在3种类型,第一种是结构构型,类似于传统构型;第二种是质态构型,物质配置格局;第三种是动态构型,反映上述二者的时间变化(叶万辉,关文彬,1995)。国内外在结构构型上进行了大量的探索,并构建了种间构型类型谱与种内外部优先法构型计量指标公式(Whitney,1976;Mc-Mahon&Kronauer,1976;孙书存&陈灵芝,1999a,1999b;卢康宁&张怀清,2010)。内部优先法能反映树体发育的自然过程,依照构件分支习性来度量枝级及其分支格局,能反映树体从幼树建成为成树的基本格局。麻疯树采用了内部优先法测定构型(何亚平等,2010),杉木研究系统建立各类构型参数特性(卢康宁等,2011)。麻疯树适生于干热河谷或干燥气候生境,遭受火烧、人为与牲畜干扰较为严重,萌生能力强,且伴随着生殖器官形成而形成二叉分支格局,并在分支结构度退化至一定阈值以下时,仅能保持自身生长功能而无法分支(未发表资料)。麻疯树栽植时,当年存活,抽发一级枝,建立了树体基本骨架;来年一级枝分支而水肥充足与干扰发生时,主干与一级枝上产生萌枝,增加新的骨架组成。麻疯树在栽植1 a~2 a内建立的骨架是适应水肥条件的产物,形成了自耗性与互补性与镶嵌性的分支格局,且在短期内无人为干扰时,基部分支格局很难打破。二级枝与一级枝分支数量及其相应的树体资源配置格局就在这一发育时间短确立了,形成配置树体有机资源与利用无机资源的树体骨架。
实际上,在此基础上持续发育并产生三级枝,前期结构可能会对后期发育结构形成限制,基本骨架相对固定了分支发生的母枝范围与空间大小,并在构件数量持续增加时枝径减小,枝长缩短,同化能力减弱,而无法保持持续分支与功能增强,形成了分支终止而枝长、枝径适当变化的终端。在存在自疏或干扰时,会引起树体骨架结构的变化,或者外部分支部分的损伤,迅速发生萌枝,引起分支的局部调整。前期分支对后期分支的限制是基于数量与质量权衡的生活史法则,一级枝数量多而枝径、枝长相对减小;二级枝数量多而枝径、枝长相对减小;三级枝数量多而枝径、枝长减小(如图1所示)。相关分析表明,一级枝数量分别与枝径、枝长呈现负相关与不相关关系(r=-0.287,p=0.000;r=0.04,p= 0.372);二级枝数量分别与枝径、枝长呈不相关与正相关关系(r=-0.029,p=0.523;r=0.154,p= 0.001),表明数量与质量权衡在一级枝枝径中发生,但二级枝枝长呈现正偶联关系。
早期幼树存活与一级枝、二级枝发生时,空间利用相对充足,后续分支在无干扰时受到限制是必然的,这种现象在分支格局之间相关分析中也得到了证明(如表3~表8所示)。麻疯树分支还发生一类现象,就是萌生枝大发生,不但使自然二叉分枝的树体紊乱,还通过高生长量当年“上林”,更加剧了生殖枝分支的结构限制;同时,内部原有分支继续维持,对萌生枝又形成了资源共享局面,结构限制在加剧,容易形成“扫把枝”,即下部长而上部发生短小分支。萌生发生是基于结构限制,顶端输出受到结构萎缩的抑制而中下部与基部产生新的输出端;或营养积累产生局部萌生枝(朱万泽等,2007)。显然,早期发育结构对晚期发育结构的限制最终导致了萌生枝发生,而加剧结构限制诱发枯枝与自疏,进而演化为新树体构型。这种情形需要资源充足能形成扰动性萌生枝与波动性生境资源条件,而在资源贫瘠时,只能持续促成分支的结构限制。现有空间资源条件下,数量与质量权衡可能是结构限制发生的理论解释,长度容易受到构件密度的影响而容易发生正偶联反映。

图1 一级枝、二级枝数量与各自枝径、枝长的散点趋势线拟合Fig.1 Scattering and the congressing trend line between first and second-class branches and their branch diameter and length
在不同生境资源斑块下,麻疯树分支格局之间相关性构建的正相关、负相关与不相关关系,暗示了结构限制发生的情形与格局。分支率增加导致二级枝数量增多,空间利用密集,维持耗费增加,再次发生分支时同化物质生产用于该部分干枝维持的增多,自身生长的减少,引发了负相关关系,即分支率2∶1引起了后续的分支率3∶2、枝径比、枝长比等多项参数在多个样地发生递减。枝径比2∶1、枝长比2∶1增加,引起了后续枝径比、枝长比正相关关系,但由于萌生分支、生殖输出、结构干扰等扰动,在某些关系中形成无关与偶尔负相关关系(如表3至表8所示)。对于树体分支而言,充足的土壤资源供应,利于构型分支的持续发生,并为结构发育提供强劲的水分渗透压;在此基础上,还有树体结构骨架与母枝特性的影响,分支发生的方位、位置等也都直接影响了后续发育,此类也称之为空间资源。构型分支与构件也有空间生态位,其宽度与质量直接影响了分支的发育;在一个骨架日趋密集的树体内部,一些分支往往因为新分支产生而形成更强的结构限制性;结构限制与立地尺度上的密度效应基本一致,区别在于部位与方向差异,可能更具有复杂性。
麻疯树早期发生一级枝都受属于萌生分支,只有顶部分支与生殖开花习性有关,二级枝发生是在一级萌生枝发生基础上发生的。这种早期分支发生具有明显的生境资源依赖性,各类样地一级枝发生数量与分支率2∶1都具存在极为明显的生境差异。一级枝、二级枝发生数量在缓斜坡、积水坡面与农林复合样地较高,分别达到8.33枝·株-1、7.29枝·株-1、6.25枝·株-1与32.86枝·株-1、30.22枝·株-1、29.66枝·株-1,在个体尺度上形成了对生境水肥资源的一次与二次分割;天然混交林、上凸坡与陡峭斜坡(一级枝数量大于农林复合,可能与农林复合开始较晚有关)一级枝、二级枝数量相对较低,上凸坡干燥生境最低。与分支数量相对应,不同生境资源斑块也形成相应的一级枝、二级枝枝径与枝长,农林复合与缓坡地生境最大,积水坡面与混交林次之,而上凸坡与陡峭斜坡最低(如表1所示)。分支率2∶1、分支率3∶2、总体分支率在缓斜坡、农林复合相对较高,干燥凸坡与陡斜坡较低,积水坡面与天然混交林相对居中(缓斜坡分支率2∶1相对较低,例外)。枝径(长)比2∶1、枝径(长)比3∶2与总体枝径(长)比在缓斜坡与积水坡面最高,天然混交林、干燥凸坡最低(枝径比3∶2例外),而农林复合、陡峭斜坡相对居中(如表2所示)。表明,麻疯树分支格局发生也与生境资源斑块肥沃性紧密相连,在肥沃生境中分支率、枝径比、枝长比相对较大,贫瘠生境中相对较小。麻疯树构型分支生境差异用适应假说能很好的给予解释,构型分支指标大总是与生境肥沃性紧密联系,在很多木本植物构型研究也总是如此(何维明,董鸣,2001;苏建荣等,2005;孙书存,陈灵芝,1999a;袁秀英等,2009;胡喜生等,2006)。
木本植物与克隆草本植物不同,不仅具有多年维持的骨架结构,还具有机械力学特性,但这一骨架不断夯实的长期维持的结构为个体基因组占据了相对稳定的空间资源,生态位相对稳定,形成了生境资源持续利用的结构体。木本植物自身也具有分支特性,受到干扰后引起树体内部新枝发生返祖性的新枝发生——萌生分支,这种克隆形式与克隆草本发生的原因类似。草本克隆植物觅食行为是受到生境资源富集性驱动的,进而形成了克隆分支的趋水性、趋肥性、趋光性现象,这种趋性效应是克隆植物行为的适应性与可塑性(董鸣,1996,;董鸣等,2011)。适应性假说揭示了克隆发生的生境诱因,解释了克隆行为可塑性的外因。但与草本克隆植物相比,木本麻疯树构型分支更多的受到自身机械力学载体的限制,或者说结构限制更为明显,发生更为频繁,但不能改变生境资源决定的命运,生境相对于构件分支而言,是大系统。且树体干扰往往也归因于结构性损伤与扰动,或者资源性打破,使得原有结构限制与资源限制加强或者宽松,进而构建新的资源利用与适宜结构的平衡体系,如游击型与密集型(李镇清,1999)。麻疯树构型与生境资源斑块之间构建了适应性的对应体系,形成了结构-生境斑块平衡体系。在群体水平与个体幼树水平上,麻疯树构件与个体构型也都与生境肥沃性建立了对应联系(费世民等,2009;何亚平等,2010)。
农林复合样地耕作松土、施肥除草、石块挡土墙水土保持,形成系统降水高效截留与利用的系统工程,水肥条件良好。缓斜坡大都位于马店河基地下方,水肥汇集与路过,加之连年施肥试验,生境资源相对良好。天然混交林个体小,密柚子与台湾相思混生抑制了中间分枝与侧枝发育,光照不足(可能保墒有作用)。上凸坡为水肥遗失地带,风力大,水分蒸发严重,土壤干燥。陡斜坡坡度较大,水、肥保蓄难度大,生境肥沃性偏低。积水坡面在凹坡与浸水台地下侧,能受到渗水的影响,水分适宜,草丛发达。这些样地组成了马店河麻疯树基地的生境资源斑块,并形成与之相对应的个体构型结构体系,形成了个体与生境相适应的生活史结构型。基于克隆植物概念在木本植物构件中也具有一般性,二者术语相似,麻疯树构型分支伴随着克隆分支的发生,也是资源限制与结构限制的产物(未发表资料)。麻疯
树栽植时形成结构性损伤与资源富集性干扰,促成了一级枝萌生大发生,形成了个体下一级枝资源配置格局,限制后续分支发生是数量与质量权衡的产物,但在特定地上空间资源利用匹配上,分支次数增加结构限制性本身在增强,麻疯树分支具有速生性,经历短暂的旺盛树冠发育后,便迅速进入结构限制状态,并且逐渐加强。早期结构制约的早期结构优化就根本上优化了构型,无需优化细枝末节,就是矛盾要抓关键,提纲挈领,顶层设计。
[1] Bell A D and Tomlinson P B.Adaptive Architecture in Rhixomatous Plants.Botanical Journal of the Linnaean Society,1980,80(20:125-160.
[2] Halle F.Architecture variation at the specific level in tropical trees.In Tom linson PB and M H Zinmeman eds Tropical trees as a living systems.London:Cambridge University Press,2009:209~222.
[3] Halle F,Oldeman R A A,Tom linson PB.Tropical trees and forests an architectural analysis.Berlin:Springer-Verlag,1978.
[4] McMahon S T A,Kronauer R E.Tree structures:Deducing the principle of Mechanical design.J.Theor.Biol.,1976,59:443~466.
[5] Moulia B,Cout C,Lenne C.Posture control and skeletalmechanical acclimation in terrestrial plants:implications for mechanical modeling of plant architecture.American Journal of Botany,2006,93(10):1477~1489.
[6] Whitney GG.The bifurcation ratio as an indicator ofadaptive strategy in woody plant species.Bulletin of Torrey Botanical C1ub,1976,103:67~72.
[7] 蔡小虎,张启珍,陈翰林.基于非线性数学模型的麻疯树树体构型优化设计[J].四川林业科技,2012,33(1):9~13.
[8] 陈波,宋永昌,达良俊.木本植物的构型及其在植物生态学研究的进展[J].生态学杂志,2002,21(3):52~56.
[9] 董鸣.植物克隆生态学[M].北京:科学出版社,2011.
[10] 董鸣.资源异质性与克隆植物生长对异质性资源响应:觅养行为[J].植物学报,1996,38:828~835.
[11] 费世民,蔡小虎,何亚平,等.生物柴油植物麻疯树群体构型对资源性环境因素的反应[J].四川林业科技,2009,30(5):1~8.
[12] 何明珠,王辉,张景光.民勤荒漠植物枝系构型的分类研究[J].西北植物学报,2005,25(9):1827~1832.
[13] 何明珠,张景光,王辉.荒漠植物枝系构型影响因素分析[J].中国沙漠,2006,26(4):625~630.
[14] 何维明,董鸣.不同气温条件下旱柳(Salix matsudana Koidz)幼苗的水分和构型特征[J].生态学报,2001,21(7):1084~1090.
[15] 何亚平,王乐辉,费世民,等.麻疯树2年生人工林构筑型对修枝与土壤肥沃性的响应[J].四川林业科技,2010,31(5):21~29.
[16] 胡喜生,洪伟,吴承祯,等.不同光环境下木荷幼苗树冠结构的可塑性响应[J].植物资源与环境学报,2006,15(2):55~59.
[17] 李亚莉,张钦弟,翟静娟,等.山西濒危植物翅果油树植冠的构型分析[J].植物资源与环境学报,2007,16(4):43~46.
[18] 李镇清.克隆植物构型及其对资源异质性的响应[J].植物学报,1999,41(8):893~895.
[19] 卢康宁,张怀清.植物构筑型研究综述[J].世界林业研究,2010,23(1):17~20.
[20] 卢康宁,张怀清,刘闽.基于实测数据的杉木构筑型研究[J].林业科学研究,2011,24(1):132~136.
[21] 苏建荣,邓疆,罗香,等.元宝槭幼树施肥研究Ⅰ.不同施肥处理对生长与构型的影响[J].林业科学研究,2005,18(2):147~152.
[22] 孙书存,陈灵芝.不同生境中辽东栎的构型差异[J].生态学报,1999a,19(3):359~364.
[23] 孙书存,陈灵芝.辽东栎植冠的构型分析[J].植物生态学报,1999b,23(5):433~440.
[24] 袁秀英,袁登胜,白玉明.固定和半固定沙丘花棒克隆生长构型的比较研究[J].西北林学院学报,2006,21(6):32~34.
[25] 张旭东,费世民,周金星.川西南山地生态脆弱区植被恢复研究[M].北京:科学出版社,2006.
[26] 朱万泽,王金锡,罗成荣,等.森林萌生更新研究进展[J].林业科学,2007,43(9):74~82.
The Structural Lim itation of Architectural Plasticity for Jatropha curcas L.Based on Internal Priority
HE Ya-ping FEIShi-min*WANG Le-hui CHEN XIU-ming MENG Chang-lai
(Sichuan Academy of Forestry,Chengdu 610081,China)
Adaptation hypothesis proposed in clone grass plants also could interpret the adaptation of architecture of tree plants to habitat resource patches,the earlier developed tree architecture would limit the latter structure,and common attentionswould not be paid to this view.In this paper,studies weremade of the adaptation of tree architecture for shrub Jatropha curcas to six patches of habitat resources and analysis wasmade of the structural limitation of this response.38 plots of 10 m×10 m forests of J.curcas were surveyed and classified into six typicalhabitats for differences in November of2010.The branch diameter,length,angle,amounts weremeasured by a unit of every first-class branch,counting the second and third class branch amounts and selecting themean branch andmeasuring the diameter,length and angle.The amounts of the first and second class branch for J.curcas had obvious responses to habitats,and there were extremely differences presented among six habitats in individual bifurcation2∶1,Rb3∶2,total Rb,Rd2∶1,Rd3∶2,total Rd,Rl2∶1,Rl3∶2,total Rl.Correlation analysis indicated that,Rb2∶1,Rd2∶1and Rl2∶1had a negative,positive or null relationship to all types of parameters in six habitats,and totally Rb2∶1had significantly negative effects and Rd(l)2∶1had relatively and obviously positive effects.The structural limitation of earlier established architecture in J curcas could attribute to tradeoff and positive tradeoff between bifurcation amounts and their diameter and length.The adaptation of J curcas architecture to different habitat re-source patches firstly established resource allocation patterns and branching pattern within individuals through branching amount control and positive and negative tradeoff with a complexity.
Jatropha curcas L.,Architecture,Internal priority,Plasticity,Structural limitation
S718.45
A
1003-5508(2016)03-0001-09
10.16779/j.cnki.1003-5508.2016.03.001
2016-01-29
麻疯树能源林生殖构型优化技术与应用。
何亚平(1977-),男,硕士,助理研究员,主要从事森林生态,森林培育研究工作。
,feism@163.com
