华西亚高山植物园多鳞杜鹃的开花物候与生殖特征1)
2016-08-17陶锐蔡仕珍马文宝姬慧娟郭洪英刘兴良
陶锐 蔡仕珍 马文宝 姬慧娟 郭洪英 刘兴良
(四川农业大学,成都,611130) (四川省林业科学研究院)
华西亚高山植物园多鳞杜鹃的开花物候与生殖特征1)
陶锐蔡仕珍马文宝姬慧娟郭洪英刘兴良
(四川农业大学,成都,611130)(四川省林业科学研究院)
通过运用开花物候指标,于2014年和2015年分别对分布于华西亚高山植物园内的多鳞杜鹃进行开花物候观察、统计和分析。结果表明:多鳞杜鹃花期为4月上旬至5月初,种群、个体、花序和单花水平上花期持续时间分别为24~25、22、13~15、7~9 d。单花开花进程依其开花形态和散粉特征可分为散粉前期、散粉初期、散粉盛期和散粉末期。多鳞杜鹃具有较高的花期同步指数0.815,开花进程呈单峰曲线,表现出一种“大量、集中”的开花式样;个体相对开花强度分布频率集中在10%~50%。开花物候指数与座果数之间的相关性分析表明,始花时间与花期持续时间呈显著负相关,花期持续时间及开花数与座果数均呈显著正相关。多鳞杜鹃开花物候在种群内年度间的相似性说明,植物个体开花物候进程很大程度上是由其遗传因子决定的,而开花物候在年度间的变异,可能是由于气候环境的差异(主要是温度和降水量)引起的。“集中开花模式”、较高的同步指数和较长花期持续时间是多鳞杜鹃适应高山环境的生殖特性。
多鳞杜鹃;开花物候;相对开花强度;开花同步性;座果数
By applying flowering phenology indicators, observation, statistics and analysis of flowering phenology ofRhododendronpolylepisdistributed in West China Subalpine Botanical Garden were conducted in 2014 and 2015, respectively. The flowering period ofR.polylepisis in the first ten days in April to early May, and flowering duration period of species, individual, single flower level is 24-25 d, 22 d, 13-15 d and 7-9 d, respectively. Single flowering processes, according to their flowering shape and pollinating characteristics, can be divided into pre-pollinating, initial pollinating, peak pollinating and late pollinating phases.R.polylepishas a relatively high index of synchronous flowering of 0.815, and blooming process is with a single peak curve, showing a “substantial, concentrated” flowering pattern, and individual relative flowering intensity frequency distribution is concentrated in 10%-50%. By the correlation analysis between flowering phenology indicators and fruiting number, initial flowering time has significant negative correlation with flowering duration time, and flowering duration time is significantly positively correlated with fruiting number. By comparingR.polylepisflowering phenology among populations in different years, plant individual flowering phenology process is largely determined by genetic factors, and the inter-annual variation of flowering phenology may be caused by differences in climate and environment (mainly temperature and rainfall). “Centralized flowering mode”, higher synchrony index and longer flowering duration are reproductive characteristics ofR.polylepisto adapt to high mountain environment.
开花物候是植物中研究最多的特征之一,具有植物生态学方面的潜在进化意义[1-2]。开花模式能影响到植物的生殖适合度[3],因此关系到植物的生存发展。开花物候不仅与植物类群遗传特性及属内系统发生相关[4-5],还与环境因子密切联系,主导因子是光照[6]、温度[7]和降水[8]等,同时也可能受到外界环境条件和植物系统内在因素的交互作用[9]。植物个体或种群的开花物候可以用一系列参数如开花数、开花时间、开花持续时间等来量化[10-11]。有关植物开花物候的研究国外已有许多报道,国内对云锦杜鹃[12]、雪莲[13]、蒙古莸[14]、五唇兰[15]、大花百子莲[16]、蓝花丹[17]等植物进行了开花物候方面的研究,但总量相对较少。
1 试验地概况
样地设在华西亚高山植物园内,地理坐标为东经103°34′~103°48′,北纬31°11′~31°18′,海拔1 720 m,地处川西平原向青藏高原过渡地带的四川盆地西部边缘的华西雨屏带地区。年均气温10.0 ℃,1月份平均气温0 ℃,7月份平均气温18.7 ℃,极端最高温26.8 ℃,极端最低温-8.3 ℃。年均降水量1 800 mm,年均湿度81.7%,年均日照时间800 h。气候湿润,雨水多,云雾重,日照少,湿度大。土壤主要为山地黄棕壤。2014年和2015年华西亚高山植物园3—6月份主要气象因子见表1。
表12014年和2015年华西亚高山植物园3—6月份主要气象因子

时间湿度/%平均气温/℃极端最高温/℃极端最低温/℃降水量/mm2014年3月份81.24.57.22.477.04月份74.38.112.55.768.45月份69.111.515.28.778.36月份66.419.121.416.3106.92015年3月份79.55.112.5-0.880.04月份75.29.615.33.670.45月份67.513.617.39.868.66月份65.320.623.517.0113.1
注:数据源于华西亚高山植物园内放置的小型气象站(美国WatchDog,2000)。
2 材料与方法
2.1材料
2014年3月下旬在华西亚高山植物园内,选择大小一致,长势旺盛的多鳞杜鹃株丛为观测样株,株高2.5~3.2 m,冠幅约3.0 m。多鳞杜鹃为两性花,伞形总状花序,顶生,有3~6朵花;雄蕊10枚,不等长伸出花冠外;花梗和花萼红色,密被鳞片;花冠宽漏斗状;子房5室;蒴果圆锥状;花期4—5月份,果期6—8月份。
2.2群体、个体和花序水平的开花物候
参照Dafni et al.[24]的方法,分别于2014年和2015年4—5月份在样地内的多鳞杜鹃种群内,于开花前随机标记人为影响小、大小基本一致的30株个体,在已标记的样株上随机标记1个花序(共30个),开花后每天统计新开花数及花期持续时间。观察和记录以下物候指标。
始花时间:群体上以样地内25%的个体开花时为始花时间;个体和花序水平上以第一朵花在个体和花序上开花的日期为始花时间。
开花高峰时间:群体上以50%的个体达到开花高峰时为开花高峰时间;个体和花序水平上以开花数大于或等于50%时为开花高峰日期。
总之,固定资产投资项目管理必须依靠信息化的手段将管理贯穿整个项目的始终,实现对项目的各阶段和环节科学的、规范的、有效的管理,解决目前面临的问题,提高项目建设总体水平,为科研活动的开展起到基础保障作用。
终花时间:群体上以95%的植株开花结束时为终花时间;个体和花序水平上以无花开放时为终花时间。
个体和花序水平的物候指数以所标记的全部个体的平均值计算。根据开花数观测计算开花物候参数。
开花同步指数(Si)用于检测开花同步性高低,根据McIntosh[25]的方法,并稍作调整:

式中:ej表示个体i和j花期重叠时间(d);fi表示个体i开花的总时间(d);n表示样地中个体总数。Si的变异范围为0~1,其中“0”表示种群内个体花期无重叠,“1”则表示完全重叠。
相对开花强度的计算:该植株开花高峰日产生的花数与该种群中植株在其开花高峰日产生的单株最大花数之比。
开花振幅的计算参考Newstrom et al.[26]的定义,文中是指标记个体和花序上每天的开花数,用花数·株-1·d-1表示。
2.3单花水平的开花物候
在已标记的每个花序上观察1个花蕾(共30个),开花后持续定位观察并记录花朵开放至凋谢的持续时间。花开放后,每2~3 h观察1次,记录花朵开放次序、花瓣大小及颜色变化、花丝伸长时间、花药散粉时间、花气味开始出现和持续时间以及柱头与花药的位置。
始花日期的统计参考肖宜安等[27]的方法并稍作调整。文中2014年多鳞杜鹃以4月8日为第1天(计为1),4月9日为第2天(计为2),依此类推;2015年以4月3日为第1天(计为1),4月4日为第2天(计为2),依此类推。
2.4数据分析
对2014年和2015年所标记个体随后发育成幼果的数目进行座果数的统计。用Pearson相关系数对始花日期、花期持续时间、开花数和座果数之间的关系进行相关分析。用单因素方差分析方法(One-way ANOVA)比较年度间开花物候特征的差异。所有数据均用Excel和SPSS 19.0统计软件进行统计分析。
3 结果与分析
3.1种群、个体和花序水平的开花物候
多鳞杜鹃每年开花1次,花期从4月上旬至5月初,其种群、个体和花序水平的开花物候特征见表2。在种群水平上,2014年始花日期比2015年晚6 d,花期持续时间少1 d、终花日期晚5 d;在个体水平上,2014年的始花日期和开花高峰日期均比2015年的晚5 d,2014年和2015年花期持续时间均为22 d,终花日期分别为5月1日和4月26日,前者开花同步指数高于后者,开花振幅为28~34朵花;花序水平上,2014年始花日期比2015年的晚5 d,终花日期比2014年晚3 d,开花振幅为2~3朵花。

表2 2014年和2015年多鳞杜鹃植物种群、个体和花序水平的开花物候
2014年和2015年多鳞杜鹃个体的开花进程基本相似,均呈单峰曲线,开花比例总体上呈先上升后下降的趋势(图1)。2015年始花后开花比例快速上升,于第12 d(4月19日)到达开花高峰,然后快速下降;而2014年则有所不同,在第10 d(4月17日)至第14 d(4月21日)内开花进程缓慢上升至高峰,然后迅速降低。
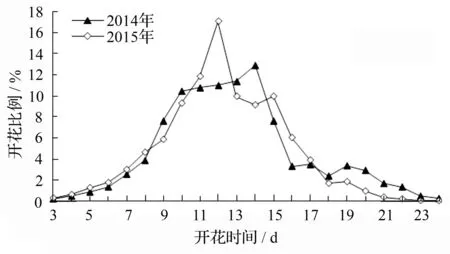
图1 2014年和2015年多鳞杜鹃植物个体的开花物候曲线
2014年和2015年多鳞杜鹃个体的始花时间、开花高峰时间、终花时间及花期持续时间的变化规律见图2。对多鳞杜鹃个体始花时间、开花高峰时间、相对开花强度等进行单因素方差分析表明,始花时间(F=0.145,P>0.05)、开花高峰时间(F=0.077,P>0.05)、相对开花强度(F=1.090,P>0.05)在年度间差异不显著,但花期同步指数(F=6.856,P<0.05)、终花时间(F=10.427,P<0.05)和花期长度(F=7.670,P<0.05)在年度间存在显著差异。
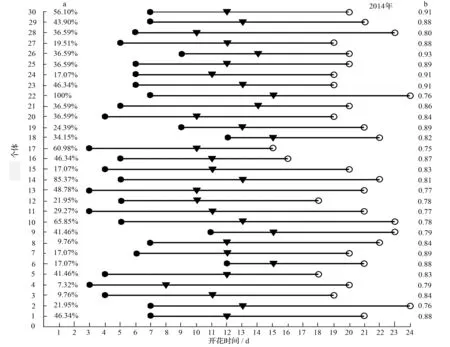
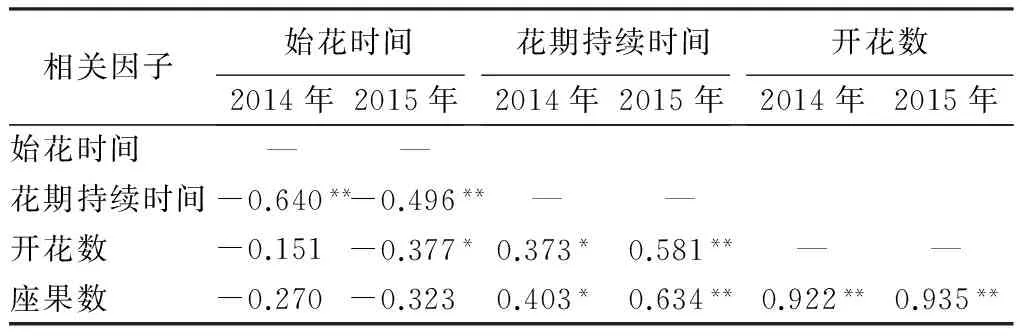
两年间多鳞杜鹃个体的平均开花同步指数为0.815,同步指数较高,表明其“大量、集中”的开花式样(图2)。2014年,多鳞杜鹃个体的相对开花强度(Q)主要集中在10% 表3 2014年和2015年多鳞杜鹃个体相对开花强度分布 3.2单花水平的开花物候 根据多鳞杜鹃花部特征及散粉过程将单花开放进程分为4个时期,即散粉前期、散粉初期、散粉盛期和散粉末期(图3),单花花期为7~9 d(表4)。在开花过程中,花丝和花柱都有弯曲和伸长,但生长速度不同,位置存在动态变化。散粉前期,柱头略高于花药,花药未散粉;散粉初期和盛期花丝的生长速度要慢于花柱,柱头始终高于花药,在此过程中花药由开始散粉并大量散粉;在散粉末期即凋谢期,花丝和花药继续生长,花丝的生长速度要高于花柱,柱头近高于花药,花药停止散粉。散粉前柱头颜色为黄色,花药和花瓣颜色为紫红色;散粉后柱头和花药成褐色,花瓣颜色变淡。花朵无明显香味,花瓣基部有花蜜,无甜味,但含量较少,其性质有待进一步研究。 表4 多鳞杜鹃单花的开花动态 左端数值a为相对开花强度;右端数值b为花期同步指数。 1.散粉前期;2.散粉初期;3.散粉盛期;4.散粉末期。 3.3物候指数和座果数的相关性 多鳞杜鹃两年度的始花时间、花期持续时间、开花数及座果数的相关分析结果(表5)显示,2014年始花时间与座果数、开花数呈负相关,与花期持续时间呈极显著负相关;花期持续时间与开花数、座果数呈显著正相关;开花数与座果数呈极显著正相关。2015年始花时间与座果数、开花数呈显著负相关,花期持续时间与座果数呈极显著正相关,其余与2014年相同。相关性分析结果表明,始花时间早的比始花时间迟的开花数多和拥有更长的花期持续时间;花期长的比花期短的开花数多,座果数高;开花数多的个体座果数高。2015年与2014年相比这一结果更加突出。 表5 2014年和2015年多鳞杜鹃物候指数及座果数的相关分析 注:*相关性达0.05的显著性水平(双尾检验);** 相关性达0.01的极显著水平(双尾检验)。 多鳞杜鹃的始花时间、开花高峰时间、相对开花强度在年度间差异并不显著,并且个体开花物候进程都呈现单峰式渐进曲线,说明植物在不同生存环境压力下表现出的适应能力主要受遗传因素影响。而开花物候在年度间出现的变异,可能与持续降水、温度、光照、霜冻显著相关[28]。华西亚高山植物园地处四川盆地西部边缘的华西雨屏带地区,气候阴湿为主,持续的阴雨天气会影响花朵开放和传粉昆虫的传粉效率。在多鳞杜鹃花期的4—5月份,亚高山植物园年度间的温度和降水量变化较大,但湿度变化不明显。2014年4月份和5月份的平均气温分别为8.1 ℃和11.5 ℃,降水量分别为68.4、78.3 mm,2015年4月份和5月份平均气温分别为9.6、13.6 ℃,降水量分别为70.4、68.6 mm,前者平均气温显著低于后者,前者降水量在4月份低于后者,而在5月份高于后者。2015年较高的气温可使多鳞杜鹃的花期提前,年度间花期变化如下,2015年种群水平上的始花日期较2014年提前6 d,个体和花序水平上较2014年提前5 d。2015年4月份极端最高温15.3 ℃,极端最低温3.6 ℃;2014年4月份极端最高温12.5 ℃,极端最低温5.7 ℃,前者温度波动幅度大于后者,致使2015年4月份阴雨天气增多,降水量增加,试验时观测到许多花朵及花蕾在此环境条件下脱落,进而直接影响座果数。由此可看出多鳞杜鹃开花可能主要受到温度和降水量的影响,温度可能是引起年度间物候差异的重要原因。 相对开花强度是植物花资源空间分布的一个指标,也可能影响花粉的运动模式。多数植物具有较低的相对开花强度[29],也有表现出高、低两种开花强度分异趋势的[27]。本研究中,多鳞杜鹃具有较低的相对开花强度,为10% 个体的开花模式常在两个极端之间变化,一个极端是“集中开花模式”,或被称为“总巢式开花模式”[30]。另一个极端是“多重高峰模式”[31]。前者在单位时间内大量开花,吸引更多传粉者访问,进而获得更多生殖成功的机会,是对受干扰和特殊环境条件的一种适应[32]。后者能够在单位时间内控制开花数量,通过延长花期,多次开花高峰的方式来获得更多的生殖机会。在连续两年的多鳞杜鹃开花物候研究中,个体水平的开花物候进程表现出较高的相似性,都呈单峰曲线模式,并且具有较高的开花同步指数0.815,表现出一种“集中开花模式”。这与准噶尔无叶豆[33]、雪莲[34]、金城山柔毛淫羊藿[35]的开花模式一致。 花期同步性指数是提高种子生殖成功和杂交水平的重要因子,同时也是种群遗传变异的决定性因素之一[36]。多鳞杜鹃两年间的开花同步指数分别为0.841和0.788,开花同步指数较高,可以保证单花开放时有足够的花资源进行雌雄交配,增加了传粉者的访花频率和异交成功的机会,同时也增加了花粉躲避捕食的能力,提高了生殖适合度。对大多数个体来说,高的同步性指数可以在一定区域内提高异花授粉的几率和有利于遗传变异的发生[37],反过来则促进遗传多样性,从而提高物种适应恶劣条件的能力[38]。 花期持续时间与植物繁殖成功率密切相关[39]。许多高山植物为了适应多雨雪、少昆虫的严酷环境,大多会选择延长花期来保证有性生殖的成功。云南黄连花期长达4个月,其中近两个多月时间处于多雨雪的低温天气,传粉昆虫的活动受到严重限制。为了适应于高山生态环境并保证其有性生殖的顺利进行,不得不采取延长花期的生殖策略[40]。2014年和2015年多鳞杜鹃个体花期持续时间均为22 d,较长的花期有利于其在高山环境下保证生殖成功。此外,多鳞杜鹃单花花期持续时间为7~9 d,较长的花寿命有利于花接受更多的拜访提高异交成功的几率。 相关性分析表明,多鳞杜鹃花期持续时间、开花数与座果数均呈显著正相关。开花数多有利于吸引更多传粉者,花期长则有利于提高传粉效率,进而提高座果数。这与绝大多数植物的研究结果相似[33-35,41-43]。 综上所述,多鳞杜鹃的生殖特性受开花模式、花期同步性指数、花期持续时间的影响,具体表现在它们与开花数和座果数的相关性上。“集中开花模式”、较高的同步指数、较长花期可以提高开花数,吸引更多传粉者,进而提高座果数,是多鳞杜鹃适应高山环境的生殖特性。 [1]FENNER M. The phenology of growth and reproduction in plants[J]. Perspectives in Plant Ecology,1998,1(1):78-91. [2]SAKAI S. Phenological diversity in tropical forests[J]. Population Ecology,2001,43(1):77-86. [3]RATHCKE B, LACEY E P. Phenological patterns of terrestrial plants[J]. Annual Review of Ecology and Systematics,2003,16(4):179-214. [4]KOCHMER J P, HANDEL S N. Constraints and competition in the evolution of flowering phenology[J]. Ecological Monographs,1986,56(4):303-325. [5]OLLERTON J, DIAZ A. Evidence for stabilizing selection acting on flowering time inArummaculatum(Araceae): the influence of phylogeny on adaptation[J]. Oecologia,1999,119(3):340-348. [6]MARCO D E, PAEZ S A. Phenology and phylogeny of animal-dispersed plants in a Dry Chaco forest (Argentina)[J]. Journal of Arid Environments,2002,52(1):1-16. [7]MARQUES M C M, ROPER J J, SALVALAGGIO A P B. Phenological patterns among plant life-forms in a subtropical forest in southern Brazil[J]. Plant Ecology,2004,173(2):203-213. [8]BORCHERT R. Phenology and flowering periodicity of Neotropical dry forest species: evidence from herbarium collections[J]. Journal of Tropical Ecology,1996,12(1):65-80. [9]ARMBRUSTER W S. The origins and detection of plant community structure: reproductive versus vegetative processes[J]. Folia Geobotanica & Phytotaxonomica,1995, 30(4):483-497. [10]OLLERTON J, LACK A J. Flowering phenology: an example of relaxation of natural selection?[J]. Trends in Ecology and Evolution,1992,7(8):274-276. [11]WHITEHEAD D R. Wind pollination: some ecological and evolutionary perspectives[M]. Orlando: Pollination Biology, Academic Press,1983:97-108. [12]边才苗,金则新.天台山云锦杜鹃的开花与结实特性[J].园艺学报,2006,33(1):101-104. [13]戴攀峰,谭敦炎.雪莲的开花生物学特性及其生态适应意义[J].植物生态学报,2011,35(1):56-65. [14]郭春燕,贺晓,高建平,等.蒙古莸的开花物候与生殖特征[J].西北植物学报,2012,32(10):2040-2046. [15]张哲,宋希强,朱国鹏,等.东亚特有种五唇兰的开花物候与繁殖特性[J].热带作物学报,2013,34(7):1223-1227. [16]孙颖,王阿香,刘颖竹,等.大花百子莲的开花物候与生殖特性[J].西北植物学报,2013,33(12):2423-2431. [17]高素萍,张硕,吴佩纹.二型植物蓝花丹开花物候特征观察及其生态意义分析[J].植物资源与环境学报,2015,24(1):84-90. [18]胡琳贞,方明渊.中国植物志(第57卷)[M].北京:科学出版社,1994:81. [19]钟国华,胡美英.杜鹃花科植物活性成分及作用机制研究进展[J].武汉植物学研究,2000,18(6):509-514. [20]周兰英,王永清,张丽.26种杜鹃属植物花粉形态及分类学研究[J].林业科学,2006,44(2):55-63. [21]庄平.中国杜鹃花属植物地理分布型及其成因的探讨[J].广西植物,2012,32(2):150-156. [22]赵冰,张果,司果臣,等.秦岭野生杜鹃花属植物种质资源调查研究[J].西北林学院学报,2013,28(1):104-109. [23]庄平.华西亚高山植物园迁地保育的42种杜鹃属植物开花-展叶物候节律[J].广西植物,2014,22(4):1-9. [24]DAFNI A, KEVAN P G, HUSBAND B. Practical pollination biology[M]. Cambridge: Enviroquest Ltd.,2005:3-26. [25]MCINTOSH M E. Flowering phenology and reproductive output in two sister species ofFerocactus(Cactaceae)[J]. Vegetatio,2002,159(1):1-13. [26]NEWSTROM L E, FRANKIE G W, BAKER H G, et al. Diversity of long-term flowering patterns[M]. Chicago: Chicago University Press,1994:142-160. [27]肖宜安,何平,李晓红.濒危植物长柄双花木开花物候与生殖特性[J].生态学报,2004,24(1):14-21. [28]TORRES C, GALETTO L. Flowering phenology of co-occurring Asteraceae: a matter of climate, ecological interactions, plant attributes or of evolutionary relationships among species[J]. Organisms Diversity & Evolution,2011,11(1):9-19. [29]BUIDE M L, DIAZ J A, GUITIAN J. Flowering phenology and female reproductive success inSileneacutifoliaLink ex Rohrb[J]. Plant Ecology,2002,163(1):93-103. [30]OKULLO J B L, HALL J B, OBUA J. Leafing, flowering and fruiting ofVitellariaparadoxasubsp. nilotica in savanna parklands in Uganda[J]. Agroforestry Systems,2004,60(1):77-91. [31]AUGSPURGER C K. Phenology, flowering synehrony and fruit set of six neotropical shrubs[J]. Biotropica,1983,15(4):257-267. [32]BAWA K S. Patterns of flowering in tropical plants. Handbook of experimental pollination ecology[M]. New York: Van Nostrand Reinhold Company,1983:394-410. [33]马文宝,施翔,张道远,等.准噶尔无叶豆的开花物候与生殖特征[J].植物生态学报,2008,32(4):760-767. [34]戴攀峰,谭敦炎.雪莲的开花生物学特性及其生态适应意义[J].植物生态学报,2011,35(1):56-65. [35]高泽梅,权秋梅,肖娟,等.金城山柔毛淫羊藿的开花物候与生殖特性[J].广西植物,2014,34(1):46-50. [36]KHANDURI V P, KUMAR K S, SHARMA C M, et al. Annual variation in flowering phenology, pollination, mating system, and pollen yield in two natural populations ofSchimawallichii(DC.) Korth[J]. Scientific World Journal,2013(11):1653 -1656. [37]RANJITKAR S, LUEDELING E, SHRESTHA K K, et al. Flowering phenology of tree rhododendron along an elevation gradient in two sites in the Eastern Himalayas[J]. International Journal Biometeorology,2013,57(2):225-240. [38]STINSON K A. Natural selection favors rapid reproductive phenology inPotentillapulchrrima(Rosaceae) at opposite ends of a subalpine snowmelt gradient[J]. American Journal of Botany,2004,91(4):531-539. [39]TORRES-DIAZ C, GOMEZ-GONZALEZ S, STOTZ G C, et al. Extremely long-lived stigmas allow extended cross-pollination opportunities in a highAndeanplant[J]. Plos One,2011,6(5):e19497. [40]杨艳娟,谢世清,孟珍贵,等.濒危药用植物云南黄连传粉生态学研究[J].西北植物学报,2012,32(7):1372-1376. [41]李向前,贾鹏,章志龙,等.青藏高原东缘高寒草甸植物群落的开花物候[J].生态学杂志,2009,28(11):2202-2207. [42]李晓红,曾小玲,曾建军.观赏植物紫藤开花物候与生殖特征[J].井冈山大学学报(自然科学版),2010,31(4):48-55. [43]权秋梅,黎云祥.巫山淫羊藿开花物候与生殖特征[J].生态学杂志,2013,32(4):859-866. 1)北京市教育委员会科学研究与研究生培养共建项目资助(BLCXY201528)、国家“十二五”科技支撑计划项目(2013BAD01B07)。 Flowering Phenology and Reproductive Features ofRhododendronpolylepisin Huaxi Sub-alpine Botanical Garden// Tao Rui, Cai Shizhen (Sichuan Agricultural University, Chengdu 611130, P. R. China); Ma Wenbao, Ji Huijuan, Guo Hongying, Liu Xingliang(Sichuan Academy of Forestry)//Journal of Northeast Forestry University,2016,44(1):56-61. Rhododendronpolylepis; Flowering phenology; Relative flowering intensity; Flowering synchrony index; Fruit number 陶锐,男,1991年2月生,四川农业大学风景园林学院,硕士研究生。E-mail:277470442@qq.com。 蔡仕珍,四川农业大学风景园林学院,高级实验师。E-mail:415455088@qq.com。 2015年9月17日。 S685.21;Q945.6+4 1)四川省省级公益性科研院所基本科研业务费专项(JB201207和JB201409)。 责任编辑:任俐。



4 结论与讨论
