花椒窄吉丁成虫触角感器超微结构1)
2016-08-17袁丽芳谢寿安刘绥鹏吕淑杰郭新荣宋晓斌
袁丽芳 谢寿安 刘绥鹏 吕淑杰 郭新荣 宋晓斌
(西北农林科技大学,杨凌,712100)
花椒窄吉丁成虫触角感器超微结构1)
袁丽芳谢寿安刘绥鹏吕淑杰郭新荣宋晓斌
(西北农林科技大学,杨凌,712100)
采用扫描电子显微镜对花椒窄吉丁(Agriluszanthoxylumi)雌雄成虫触角进行观察,明确了花椒窄吉丁雌雄成虫触角感受器的类型、数量和分布位置及两性感受器的差异性。结果表明:在雌雄成虫触角上均发现6类17种感受器:3种毛形感器、4种刺形感器、7种锥形感器、1种钟形感器、1种芽形感器和1种特殊感器。此外在雄虫触角上还发现Böhm氏鬃毛。花椒窄吉丁触角感受器的分布位置相对稳定,触角两侧面主要分布毛形感器,背腹面主要分布毛形感器和刺形感器。触角第Ⅳ-Ⅺ节端部有1个凹窝,凹窝内主要分布锥形感器,凹窝旁的凹陷中有1个锥形感器簇,但第Ⅹ-Ⅺ节的锥形感器簇不明显。雌雄成虫的6类17种感器在分布位置上无明显差异,但是触角锯齿部分的或第Ⅳ-Ⅺ节端部锥形感器在数量上雄虫显著多于雌虫。
花椒窄吉丁;触角;感器;扫描电镜;超微结构
We observed the sensilla on the antenna ofAgriluszanthoxylumiby using scanning electron microscopy to know the ultrastructure, type, number, distribution of sensilla on the antenna ofAgriluszanthoxylumi, and the differences of sensilla between male and female. The internal and external sides of the antenna have sensilla trichodeas, sensilla trichodeas and sensilla chaeticas were distributed over segmental venter and the back side. There is a fossa on the apex of every flagellomerefrom fourth to eleventh, and sensilla basiconicas were mainly distributed over it. Sensilla basiconicas cluster in the sunken place, where is the fossa, but the sensilla basiconicas cluster is not obvious on the tenth and eleventh flagellomeres. There are 6 types (17 kinds) of sensilla on the antenna of male and female including 3 kinds of sensilla trichodeas, 4 kinds of sensilla chaeticas, 7 kinds of sensilla basiconicas, 1 kind of sensilla campaniformia, 1 kind of sensilla gemmformium and 1 kind of unusual sensilla, and Böhm bristles were on the antenna of male. The distribution of sensilla is relatively stable, the 6 types of sensilla have no difference in distribution between male and female adults, but the number of sensilla basiconicas on apex of antenna of the male is significantly more than the number of the female.
昆虫敏锐的嗅觉依赖于触角感受器对外界刺激和内部神经系统的调控,以及对内外化学信号的感受识别[1],在昆虫的取食、交配和寄主定位等行为选择中具有重要意义。
目前对昆虫触角感器的研究主要集中在鞘翅目、鳞翅目、直翅目、同翅目、半翅目、膜翅目、双翅目和缨翅目[2-4]。鞘翅目中对天牛科、小蠹科、金龟科、步甲科等研究的较多[5],吉丁虫是重要的林木钻蛀害虫,但是对其触角感器的研究较少。Volkovitsh对吉丁虫科316属412种吉丁虫的触角端部进行扫描电镜研究,归纳出吉丁虫科10类常见的触角感器类型[6]。Crook在对白蜡窄吉丁(Agrilusplanipennis)雌雄成虫触角的扫描电镜中发现5种感器,且雌雄成虫触角感器在类型、分布上并无差异[7]。刘玉双利用扫描电镜对红缘吉丁(Buprestisfairmairei)触角进行观察,发现红缘吉丁具有毛形、刺形、锥形和芽形感器4种类型[8]。
花椒窄吉丁(Agriluszanthoxylumi),鞘翅目(Coleoptera),吉丁科(Bupresdae),窄吉丁属(Agrilus),是花椒上的先锋害虫。目前对花椒窄吉丁的研究主要在化学防治和生物学特性方面[9-11],关于花椒窄吉丁触角感器的研究尚未见报道。本试验以花椒窄吉丁为研究对象,利用扫描电子显微镜对其雌雄成虫触角感器进行系统研究,并探讨各类型感器的功能,旨在丰富花椒窄吉丁感器研究资料,完善花椒窄吉丁形态学研究,为今后进行化学生态学研究及信息素的研制提供理论基础。
1 材料与方法
1.1供试虫源
花椒窄吉丁成虫于2015年7月中下旬采自陕西凤县白石铺花椒基地附近山地的椒园。
1.2样品的制备和观察
将花椒窄吉丁成虫放于解剖镜下分辨雌雄,然后取下头部,用刀片将触角切下。分别将雌雄成虫的触角用70%的乙醇浸泡并于冰箱中4 ℃保存备用。将样品用70%的乙醇清洗2次,然后依次在80%、85%、90%、95%和100%(2次)的乙醇中进行梯度脱水,自然晾干。分别将雌雄成虫的触角分不同的侧面粘于样品台上,Hitachi E-1010离子溅射仪喷金,并于Hitachi S-3400N扫描电子显微镜(日立)下观察拍照,工作电压5~15 V。
1.3感器的命名
触角感觉器的类型主要采用Schneider、Zacharuk和Shields的触角感觉器命名系统[12-14]。
1.4数据与图片处理
利用统计分析软件SPSS 17.0对触角感受器的数量进行统计并作差异性分析,用Photoshop 7.0对图片进行标识和排版,利用Hitachi S-3400N扫面电子显微镜的系统软件SEM Data Manager对感器进行测量。
2 结果与分析
2.1触角的基本结构
花椒窄吉丁成虫触角较短,长2 mm左右,呈锯齿状,11节,分柄节、梗节和鞭节,着生在复眼间的触角窝内(图1A-B)。柄节最长,约250 μm,基部直径最大,相对粗壮。梗节比柄节稍短,与鞭节第1亚节(第Ⅲ节)等长,形状相似。鞭节分9个亚节(第Ⅲ-Ⅺ节),从第Ⅳ节开始明显呈锯齿状,且触角第Ⅳ-Ⅺ节端部具一个椭圆形的凹窝(雌虫凹窝长×宽为43.8 μm×27.6 μm;雄虫凹窝长×宽为57.8 μm×31.5 μm;凹窝深5~10 μm),凹窝内密布锥形感受器(图1J-L)。凹窝外侧有一个较浅的凹陷(凹陷深2~5 μm;面积为凹窝的1.5~2.0倍),其内部有一个锥形感器簇(图1L),但该感器簇在第Ⅹ节和第Ⅺ节上分布较分散,簇状结构不明显。触角表面具鳞片状刻纹,柄节上感器的数量和类型较少,柄节到鞭节感器的数量和类型逐渐增多。
2.2触角感受器
2.2.1毛形感受器
毛形感器Ⅰ(t1):感器呈毛发状,长50~80 μm,基部直径2.4~3.8 μm,着生在触角表面的圆窝中与其呈30°~60°夹角。毛体上有棱形纵脊,从基部到端部逐渐变细,端部尖锐,微弯或从中部呈微拱形弯曲(图1E、F、)。分布在触角梗节和鞭节的背面、腹面和两侧面,是触角表面数量较多的一类感器。
毛形感器Ⅱ(t2):较短,长10~35 μm,基部直径1.4~2.8 μm。从毛体2/3处到端部变细、弯曲,端部较钝,着生在柄节和鞭节外侧表面的窝中,与触角表面呈45°~80°夹角(图1K)。多分布在鞭节外侧面,数量较少。
毛形感器Ⅲ(t3):着生于鞭节腹面,由触角基部指向端部,与触角表面的夹角小于30°。感器长40~70 μm,端部呈90°弯曲,弯曲长度约10 μm(图1F)。
2.2.2刺形感器
刺形感器Ⅰ(ch1):着生于鞭节背面和腹面的圆窝中,与触角呈小于30°夹角。微倾斜,不弯曲,感器整体细长,端部尖,刚直如刺,长40~62 μm,基部直径1.4~2.0 μm,表面有纵向的刻纹(图1C、F)。
刺形感器Ⅱ(ch2):着生于鞭节背面和两侧面的圆窝内,与触角表面夹角30°~45°,刺体基部到端部逐渐变细,端部稍钝,表面纵向刻纹明显,长23~50 μm,基部直径2.1~3.5 μm(图1C、H)。
刺形感器Ⅲ(ch3):着生在触角表面的圆窝中,与触角表面夹角15°~30°,刺体由基部到端部的变细不明显,端部钝圆,表面有纵向刻纹。长10~25 μm,基部直径2.5~4.0 μm(图1C、E)。
刺形感器Ⅳ(ch4):与刺形感器Ⅰ形态类似,与之不同的是该感器较细小,长20~35 μm,基部直径约1.4 μm,生长在触角表面的圆窝中,但该圆窝直径较小(图1H)。
2.2.3锥形感器
锥形感器Ⅰ(sb1):直立,圆柱状,基部具圆台形隆起,端部钝圆。感器长7~9 μm,基部直径约1.5 μm,集中分布在锯齿状触角节端部的凹窝内(图1I)。
锥形感器Ⅱ(b2):与锥形感器Ⅰ大小接近或略长,但端部有一刺尖。感器长9~11 μm,基部直径小于等于1.5 μm,主要分布在锯齿状触角节端部的凹窝内和触角端部浅的凹陷内(图1I)。
锥形感器Ⅲ(b3):微倾斜,比锥形感器Ⅰ短小,圆锥状,基部具有圆台形隆起,端部钝尖。主要分布在锯齿状触角节端部的凹窝的边缘,数量相对较少(图1I)。
锥形感器Ⅳ(b4):直立,上下粗细均匀,较锥形感器Ⅰ长,感器长14~30 μm,基部直径约2.2 μm。具有圆台形隆起的底座,顶端钝圆或稍尖,主要分布在触角端部浅的凹陷中(图1D、L)。
锥形感器Ⅴ(b5):弯曲,锥体弯曲形态不同,从锥体中部微弯、从锥体端部90°弯曲。感器长10~20 μm,基部直径1.9~2.3 μm。具有圆台形隆起的底座,顶端钝圆或稍尖,主要分布在锯齿状触角端部浅的凹陷和端部表面的圆窝中(图1D、L)。
锥形感器Ⅵ(b6):基部具有大的圆台形隆起,中间细,上端呈花苞状,其上部有纵纹。集中分布在锯齿状触角节端部的凹窝内(图1I)。
锥形感器Ⅶ(b7):该感器明显短小,三角锥状,长约3.5 μm,基部直径约1.6 μm。具有圆台形隆起,但圆台形隆起的周围呈现明显的环形凹沟,与之前感器的隆起圆台不同。主要分布在锯齿状触角节端部的凹窝的边缘,数量较少(图1L)。
2.2.4钟形感器(SCA)
该感器着生于约1.2 μm直径的圆形凹陷中,在超出凹陷的圆台状结构上生长有乳头状突起。主要分布在锯齿状触角端部的凹窝内,数量较少(图1L)。
2.2.5芽形感受器(SG)
形态像植物种子刚萌发的芽,短小,顶端钝圆,直立或弯曲,有的弯曲呈S形,分布在锯齿状触角端部的凹陷内外,多与锥形感器混生(图1K)。
2.2.6Böhm氏鬃毛(BB)
直立、表面光滑、顶端钝圆的结构,着生于1个较宽的圆窝中。位于梗节的端部,鞭节第一、二亚节的基部。也常被描述为刺形感器(图1G)。
2.2.7特殊感器(US)
着生于触角表面的椭圆窝中(长×宽:5.0 μm×2.5 μm),窝内并排着2个圆锥形感器,端部钝圆,雄虫椭圆形窝内的锥形感器较短小(图1J)。雌虫在鞭节第2亚节端部与第3亚节连接处,雄虫在第8亚节端部大凹窝旁,并非所有雌雄成虫都存在该感器。

A.雌性触角正面观;B.雌性触角背面观;C.雌虫第Ⅷ鞭节上毛形、刺形感器;D.触角鞭节端部的锥形感器;E、F.毛形、刺形感器;E.中框为毛形感器基部放大图;G.雄虫第Ⅲ、Ⅳ鞭节上的感器;H.触角端部感器;I.触角端部凹窝内锥形感器;J.雌虫第Ⅳ节上毛形、刺形、特殊感器;K.雌虫第Ⅷ节上的毛形,芽形感器;L.雄虫第Ⅵ节上触角端部感器,右上框内为锥形感器簇,右下为钟形感器。
图1花椒窄吉丁触角感受器
2.3触角感器的数量和分布
花椒窄吉丁触角上各类感受器的位置分布和数量见表1和表2。花椒窄吉丁成虫触角柄节上分布的感受器的类型和数量都较少。梗节存在一定数量的毛形感器和刺形感器,鞭节的第1亚节(第Ⅲ节)和梗节上的感器类型相似,包括除t3外的2种毛形感器和所有的刺形感器。鞭节第Ⅳ-Ⅺ节,触角的两侧面主要分布毛形感器,背面、腹面存在毛形感器和刺形感器。触角端部有一个凹窝,凹窝内主要有5种锥形感器,包括b1、b2、b3、b6和b7。触角的端部并不平整,存在一个比凹窝浅的凹陷,凹陷内有2种锥形感器b4、b5,一个由锥形感器聚集而成的锥形感器簇,此外还有为数不多的钟形感器和与锥形感器混生的芽形感器。Böhm氏鬃毛仅在雄虫中发现3个,雌雄成虫均发现一个特殊感器(US),但并非所有雌雄成虫都存在。

表1 花椒窄吉丁两性成虫触角感受器的数量和分布
注:表中的数值为8根触角每节感器的平均值,8个不同虫体分别选取单根触角做样本。Böhm氏鬃毛和特殊感器(US)未在表中列出,触角呈锯齿状膨大的那一面为外侧面,其对面为内侧面。
2.4两性成虫触角差异性
花椒窄吉丁雌雄触角各节长度和基部直径见表3,经t检验分析得出,雌雄成虫触角在长度和基部直径上无明显差异(P>0.05)。由表1和表2可知,花椒窄吉丁雌雄成虫触角感受器在各节的种类和分布位置一致,但数量上存在雌雄二性型。经过t检验分析,触角背面、腹面和两侧面的毛形、刺形感器在数量上雌雄不存在明显差异(P=0.89,P>0.05),而鞭节端部锥形感器的数量和类型雌雄差异明显(P<0.05),在总数上,雄虫明显多于雌虫,但雌虫b2的数量显著多于雄虫,而雄虫b1、b3、b5和b6的数量显著多于雌虫(表4)。

表2 花椒窄吉丁两性成虫触角第Ⅵ-Ⅺ节端部感器数量与分布
注:表中的数值为8根触角每节感器的平均值,8个不同虫体分别选取单根触角做样本。Böhm氏鬃毛和特殊感器(US)未在表中列出。

表3 花椒窄吉丁两性成虫触角各节长度
注:两性成虫各选8个不同虫体的触角做样本,表中数据为平均值±标准误。
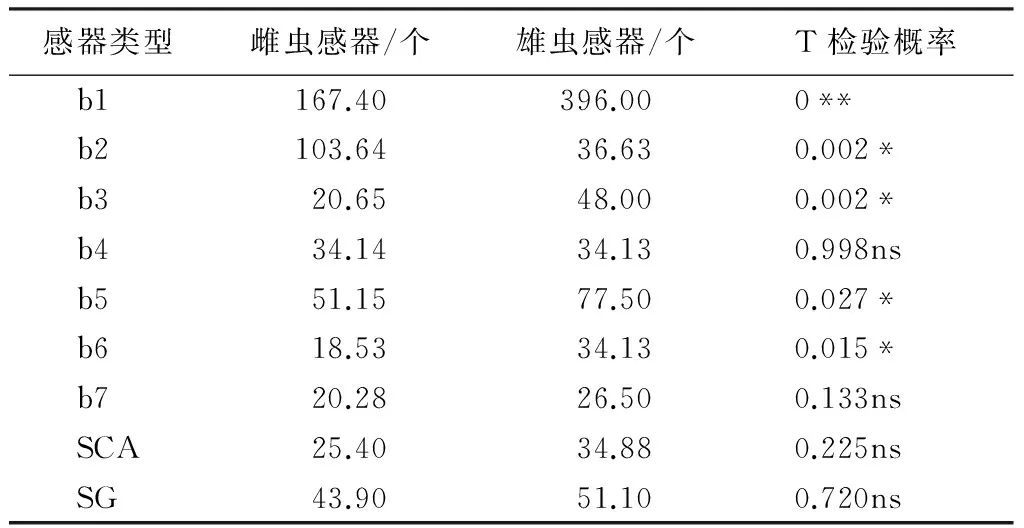
表4 花椒窄吉丁两性成虫触角第Ⅵ-Ⅺ节端部感器数量对比
注:表中数值是触角第Ⅳ-Ⅺ感器总数,** 表示差异性极显著(P<0.01),*表示差异性显著(P<0.05),ns表示无显著性差异。
3 讨论
在本研究中,笔者发现花椒窄吉丁触角感器有7种类型,分布数量较多的有毛形、刺形和锥形感器。触角第Ⅳ-Ⅺ节的端部有一个凹窝,凹窝内布满锥形感器,这种结构在吉丁虫总科很普遍,常用来做分类研究[6,15]。除Böhm氏鬃毛仅在雄虫中发现,其他感器在雌雄成虫触角上均有分布,且在分布位置上无明显差异。
毛形感器同时具有机械感器和化学感器的功能[16],为鞘翅目大多数昆虫所有,如绿豆象(Callosobruchuschinensis)、暗黑鳃金龟(Holotrichiaparallela)、红脂大小蠹(Dendroctonusvalens)、奇步甲(Carabusprodigus)等[17-20]。Crook认为白蜡窄吉丁毛形感器SC为机械感器,刘玉双推测红缘吉丁虫中毛形感器Ⅰ为机械感器,毛形感器Ⅱ为化学感器[7-8]。白蜡窄吉丁中感器SC、红缘吉丁中毛形感器Ⅰ与本研究中t1是同类型感器,且t1在雌雄之间无差异,以此推测t1为机械感器。许多文献显示毛形感器具有感知信息素的功能,如稻虱红鳌蜂(Callosobruchuschinensis)毛形感器Ⅰ[18,21]。t2在触角外侧面,可以更先接触到周围环境的的气味,红缘吉丁中毛形感器Ⅱ与本研究中t3是同类感器,所以推测t2、t3为化学感受器。同时笔者发现在毛形感器的基窝中有小孔(K)(图1I),这种小孔与毛形感器的关系及作用还不清楚。
刺形感器与毛形感器一样,几乎遍及每一种昆虫,具最基本的感受机械刺激功能,同时也是一种接触性化学感受器[17,22-23]。吉丁虫与叩甲的亲缘关系密切[8],且花椒窄吉丁刺形感器分布在的触角背面和腹面,其所占触角面积和与环境的接触面积都较大,因此推测其也具有接触感知化学刺激的作用[24-25]。
锥形感器具有化学感受器的功能,在生境中与寄主定位与选择中有重要作用。本研究中的b1与Crook中的味觉感器U是同一感器,分布位置相同,且b1在雄虫中的数量显著多于雌虫,与Crook的研究结果[7]一致。Crook的锥形感器B1与本研究中的b5是同一感器类型,但分部的位置不同,B1在白蜡窄吉丁中分部在凹窝内部外围,b5在花椒窄吉丁触角凹窝外的端部,同时该感器在筛胸梳爪叩甲(Melanotuscribricollis)上也有发现,被描述为鱼钩形感器[7,24]。b2与鞘翅目中的孟氏隐唇瓢虫(Cryptolaemusmontrouzieri)中描述的锥形感受器4型相似,与之不同的是从感器基部到端部没有明显的逐渐变细[26]。锥形感器b6为花椒窄吉丁特有的锥形感器,在报道过的吉丁虫中未发现,但该感器在豆象中都有描述[17]。此外,笔者发现在触角端部的凹陷中有一个锥形感器簇(图1N),组成该锥形感器簇的锥形感器端部平齐,中央有孔,该锥形感器簇在报道过的吉丁虫中没有发现。
本研究发现,钟形感器分布在触角端部的凹窝和边上的凹陷中,常几个聚集分布,与Volkovitsh的观察结果[6]一致。该感器在白蜡窄吉丁和红缘吉丁中均没有发现[7-8],但是在鞘翅目杨干象(Cryptorrhynchuslapathi)和光肩星天牛(Anoplophoraglabripennis)中均有描述[27-28]。有的研究认为此类感受器是温湿度感受器,为昆虫提供环境中的温湿度信息,以便其找到合适的栖息地,Volkovitsh认为其是一种化学感受器,但花椒窄吉丁钟形感器的具体功能还需进一步研究。
本研究中芽形感器分布在触角端部,而红缘吉丁中该感器位于梗节基部[8]。桑天牛(Aprionagermari)中也有描述该感器[29],但是与本研究及红缘吉丁的芽形感器形态[8]不同,却与本研究中b5形态类似。
Böhm氏鬃毛通常存在于头部与柄节、柄节与梗节的连接处,或柄节和梗节的基部[23,30]。Fu et al.在对半翅目小褐飞虱(Laodelphaxstriatellus)的研究中发现了触角柄节上仅单个存在的Böhm氏鬃毛[31]。本研究中仅在雄虫梗节的端部,鞭节第1、2亚节的基部各发现1个。
花椒窄吉丁中的特殊感器US与Volkovitsh中感器窝(SP)有些相似,但是在Volkovitsh中通常由很多SP聚集在一起形成感器域(SF)[6],但在本研究中仅在雌雄成虫触角上各发现1个的US,白蜡窄吉丁和红缘吉丁中并未报道,这可能与物种的种属性及生物进化有一定的关系。
在多数昆虫中雌虫释放性信息素,雄虫通过触角感受性信息素寻找伴侣进而与其交配,因而雌雄触角二型现象也普遍存在[32]。东北大黑鳃金龟(Holotrichiadiomphalia)雄虫触角鳃片长于雌虫,且鳃片上嗅感器总数明显多于雌虫,其中锥形感器数量几乎是雌虫的9倍[33]。光肩星天牛雌虫触角长于雄虫,雌虫中锥形感器b1的数量多于雄虫,钟形感器未在雌虫中发现[28]。白蜡窄吉丁触角感器在类型和分布不存在明显的性二型现象,但雄虫感器U的数量显著多于雌虫[7]。本研究中花椒窄吉丁雄虫触角第Ⅳ-Ⅺ节端部凹窝明显大于雌虫,凹窝内的锥形感器总数约为雌虫的2倍。此外,Böhm氏鬃毛只在雄虫中发现,而其他感器在类型、分布和数量上雌雄无显著差异。
Merivee认为昆虫触角感受器的分布模式与昆虫的行为有一定的关系[34]。花椒窄吉丁触角感器的这种分布模式对其行为的影响尚不清楚。吉丁虫触角感器的具体功能尚未有具体研究,所以关于花椒窄吉丁感器的功能也仅是根据相关文献中对同种感器功能的介绍进行推测,然而缺乏准确性,仍需要透射电镜及电生理试验加以考证。
[1]SATO K, TOUHARA K. Insect olfaction: receptors, signal transduction, and behavior[J]. Results & Problems in Cell Differentiation,2009,47:203-220.
[2]李竹,陈力.触角感器特征应用于昆虫分类的研究进展[J].昆虫分类学报,2010,32(增刊):113-118.
[3]李彝利.西藏飞蝗触角感受器及其聚集信息素研究[D].雅安:四川农业大学,2012.
[4]李维娜,冯纪年.三种花蓟马触角感器的超微结构[J].昆虫学报,2013,56(9):1088-1100.
[5]张晓军,孙伟,张健,等.鞘翅目昆虫触角感器研究进展[J].安徽农业科学,2013,41(7):2932-2935.
[6]VOLKOVITSH M G. The comparative morphology of antennal structures in Buprestigae (Coleoptera): evolutionary trends, taxonomic and phylogenetic implications. Part 1[J]. Acta Musei Moraviae Scientiae Biologicae (Brno),2001,86:43-169.
[7]CROOK D J, KERR L M, MASTRO V C. Distribution and fine structure of antennal sensilla in emerald ash borer (Coleoptera: Buprestidae)[J]. Annals of the Entomological Society of America,2008,101(6):1103-1111.
[8]刘玉双,石福明.红缘吉丁(鞘翅目:吉丁虫科)触角感器的扫描电镜观察[J].昆虫学报,2005,48(3):469-472.
[9]张益知,李忠锋.花椒吉丁虫的初步研究[J].植物保护,1988,14(2):23-24.
[10]李孟楼,李宗明,焦爱叶.花椒窄吉丁的危害规律及其分布型[J].西北林学院学报,1990,5(1):34-38.
[11]王燕,李仁娜,刘雪涛,等.韩城花椒窄吉丁虫的发生及防治[J].陕西林业科技,2012(1):76-77.
[12]SCHNEIDER D. Insect antenna[J]. Annu Rev Entomol,1964,9:103-122.
[13]ZACHARUK R Y. Antennae and sensilla[M]//KERKUT G A, GILBERT L I. Comprehensive insect physiology, biochemistry and pharmacology. Oxford, UK: Pergamon Press,1985:1-69.
[14]SHIELDS V D C. Ultrastructure of insect sensilla[M]//CAPINERA J L. Encyclopedia of entomology. 2nd ed. New York: Springer-Verlag,2008:4009-4023.
[15]BELLAMY C L. A catalogue of the higher taxa of the family Buprestidae (Coleoptera)[J]. Navors Nas Mus Bloemfontein,1985,4(15):405-472.
[16]马瑞燕,杜家纬.昆虫的触角感器[J].昆虫知识,2000,37(3):179-183.
[17]陈新.豆象科十种仓储害虫触角感器的超微结构研究[D].长春:东北师范大学,2013.
[18]宋月芹,董钧锋,孙会忠,等.暗黑鳃金龟成虫触角感器的超微结构特征[J].福建农林大学学报(自然科学版),2013,42(2):149-152.
[19]王玉刚,张真,王鸿斌,等.红脂大小蠹成虫触角扫描电镜的观察[J].林业科学研究,2003,16(6):726-730.
[20]刘桂清,田明义.奇步甲触角感器电镜扫描观察和触角电位反应[J].华南农业大学学报,2008,29(2):50-58.
[21]李帅,陈文龙,杨洪,等.稻虱红螯蜂触角感受器的扫描电镜观察[J].应用昆虫学报,2012,49:509-514.
[22]ISIDORO N, BARTLET E, ZIESMANN J, et al. Antennal contact chemosensilla inPsylliodeschrysocephalaresponding to cruciferous allelochemicals[J]. Physiological Entomology,2002,23(2):131-138.
[23]OCHIENG S A, PARK K C, ZHU J W, et al. Functional morphology of antennal chemoreceptors of the parasitoidMicroplitiscroceipes(Hymenoptera: Braconidae)[J]. Arthropod Structure & Development,2000,29(3):231-240.
[24]彭观地,舒金平,张爱良,等.筛胸梳爪叩甲触角感器的电镜扫描观察[J].林业科学,2012,48(8):106-116.
[25]MERIVEE E, RAHI M, LUIK A. Antennal sensilla of the click beetle,Melanotusvillosus(Geoffroy) (Coleoptera: Elateridae)[J]. International Journal of Insect Morphology and Embrylolgy,1999,28(4):41-45.
[26]刘平,张宇宏,吴红胜,等.孟氏隐唇瓢虫的触角感受器[J].环境昆虫学报,2013,35(4):478-485.
[27]常立儒,张璐,王涛,等.杨干象触角感器的扫描电镜观察[J].植物检疫,2014,28(3):23-29.
[28]闫雄飞,孙月琴,刘永华,等.光肩星天牛触角感受器的环境扫描电镜观察[J].林业科学,2010,46(11):104-109.
[29]诸葛飘飘,葛红梅,王满囷,等.桑天牛头部附器感器的扫描电镜观察[J].昆虫知识,2009,46(2):238-244.
[30]MERIVEE E, PLOOMI A, RAHI M, et al. Antennal sensilla of the ground beetleBembidionlamprosHbst (Coleoptera, Carabidae)[J]. Acta Zoologica,2000,81(4):339-350.
[31]FU B X, BELLIS G A, HONG J, et al. Morphology, distribution, and abundance of antennal sensilla of male and female macropterous and brachypterous small brown planthopper,Laodelphaxstriatellus(Fallén) (Hemiptera: Dephacidae)[J]. Microscopy Research & Technique,2012,75(11):1492-1512.
[32]KIM J Y, LEAL W S. Ultrastructure of pheromone-detecting sensillum placodeum of the Japanese beetle,PopilliajaponicaNewmann (Coleoptera: Scarabaeidae)[J]. Arthropod Structure & Development,2000,29(2):121-128.
[33]孙凡,胡基华,王广利,等.东北大黑鳃金龟嗅感器超微结构[J].昆虫学报,2007,50(7):675-681.
[34]MERIVEE E, PLOOMI A, RAHI M, et al. Anennal sensilla of the ground beetleBembidionproperansSteph. (Coleoptera, carabidae)[J]. Micron,2002,33(5):429-440.
Ultrastructure of the Antennal Sensilla ofAgriluszanthoxylumi//
Yuan Lifang, Xie Shouan, Liu Suipeng, Lü Shujie, Guo Xinrong, Song Xiaobin
(Northwest A&F University, Yangling 712100, P. R. China)//Journal of Northeast Forestry University,2016,44(1):94-99,102.
Agriluszanthoxylumi; Antenna; Sensilla; Scanning electron microscope; Ultrastructure
袁丽芳,女,1986年2月生,西北农林科技大学林学院,硕士研究生,E-mail:Ylifang1225@126.com。
谢寿安,西北农林科技大学林学院,教授。E-mail:shouanxie@163.com。
2015年10月14日。
S763.7;Q964
1)国家公益性行业(林业)科研专项(201504324);西北农林科技大学2013年度试验示范站(基地)科技创新与成果转化专项项目(XNY2013-31)。
责任编辑:程红。
