MDA对草鱼肠道黏膜结构屏障损伤和肝胰脏、肠道胆固醇、胆汁酸合成影响
2016-08-10黄雨薇叶元土蔡春芳陈科全徐登辉林秀秀罗其刚
黄雨薇 叶元土 蔡春芳 吴 萍 陈科全 吴 韬 徐登辉 彭 侃 林秀秀罗其刚
(苏州大学基础医学与生物科学学院, 苏州 215123)
MDA对草鱼肠道黏膜结构屏障损伤和肝胰脏、肠道胆固醇、胆汁酸合成影响
黄雨薇 叶元土 蔡春芳 吴 萍 陈科全 吴 韬 徐登辉 彭 侃 林秀秀罗其刚
(苏州大学基础医学与生物科学学院, 苏州 215123)
油脂为鱼类的生长提供能量和必需脂肪酸, 因此在饲料中得到广泛应用。然而, 油脂由于含有大量不饱和脂肪酸, 特别是鱼油, 在高温、高湿条件下特别容易氧化酸败, 产生多种初级和次级氧化产物, 这些氧化产物被鱼类摄食后, 会破坏其正常的生理功能, 危及健康生长[1, 2]。目前, 被广泛关注的次级氧化产物有四羟基壬烯醛(4-HNE)和丙二醛(MDA)等[3]。MDA作为多不饱和脂肪酸氧化的最主要产物, 具有半衰期长和反应性高的特点, 能通过细胞脂质过氧化, 破 坏细胞膜结构和功能引发蛋白质交联, 破坏酶活性, 损伤DNA等途径诱导细胞凋亡, 进而造成组织损伤[4, 5]。
肠道作为鱼体自身与外界环境接触的最大界面, 不仅具有消化吸收的作用, 还具有阻止肠腔内有害物质进入血液循环的屏障功能, 从而达到维护鱼体健康的目的。而完整的肠黏膜细胞和肠黏膜细胞间的连接作为肠黏膜机械屏障的结构基础, 共同构成了肠道的结构性屏障[6]。紧密连接是肠黏膜细胞间最为重要的连接方式,主要由跨膜蛋白Occludin[7]、Claudins[8, 9]和胞浆蛋白ZOs[10, 11]构成, 在维持肠道黏膜通透性和肠道屏障功能完整性方面具有重要作用[12]。
胆固醇又称胆甾醇, 是细胞膜的重要组成成分, 又是体内合成许多类固醇激素和胆汁酸的前体, 对于维持细胞膜的完整性和体内生命活动的正常运行具有重要意义。3-羟基-3-甲基辅酶A还原酶(3-hydroxy-3-methylglutaryl-Coenzyme A reductase, HMGCR)和胆固醇7α羟化酶(cholesterol 7alpha-hydroxylase, CYP7A1)分别是合成胆固醇和胆汁酸的关键限速酶, 在维持机体胆固醇、胆汁酸动态平衡中起着主要作用。肝胰脏不仅是鱼类重要的代谢和解毒器官, 也是合成机体胆固醇、胆汁酸的主要场所。
研究报道, 氧化油脂会对动物肠道的结构和功能造成损伤[13,14]。当草鱼肠道损伤时, 其肝胰脏、肠道的HMGCR、CYP7A1基因表达水平显著上调[15]。在草鱼灌喂氧化鱼油的试验中发现, 氧化鱼油会导致肠道黏膜胆固醇、胆汁酸合成基因通路表达上调[16]。在“氧化鱼油对草鱼肠道黏膜结构屏障损伤和肝胰脏、肠道胆固醇、胆汁酸合成影响”(已投稿)的试验中也出现了类似结果,表明氧化油脂会引起肠道黏膜结构屏障损伤和对肝胰脏、肠道胆固醇、胆汁酸合成造成影响。而MDA作为鱼油氧化的最主要产物, 是导致上述结果发生的主要因素吗?目前还未见相关报道。
本试验在添加不同浓度MDA的条件下, 采用荧光定量PCR技术(qRT-PCR)对草鱼肠道的6个紧密连接蛋白和肝胰脏、肠道的胆固醇、胆汁酸合成关键酶基因表达活性进行检测, 并结合紧密连接结构透射电镜、肠道通透性指标、血清、肝胰脏和肠道胆固醇、胆汁酸含量的变化进行综合分析, 旨在探讨MDA对草鱼肠道黏膜结构屏障损伤和肝胰脏、肠道胆固醇、胆汁酸合成的影响。
1 材料与方法
1.1 试验鱼
草鱼来源于浙江一星饲料有限公司养殖基地, 为池塘培育的1冬龄鱼种, 挑选体格健康、无畸形、体质量为(74.8±1.2) g的草鱼300尾鱼, 随机分为5组, 每组3个重复,每个重复20尾鱼。
1.2 试验饲料
以酪蛋白和秘鲁蒸气鱼粉为主要蛋白源, 豆油为主要脂肪源, 根据等氮等能的原则, 设置了一个对照组(6S)和3个MDA处理组的试验饲料, MDA-1、MDA-2、MDA-3是通过在6S组饲料中喷洒4 mL含不同浓度MDA的反应液后制得的3组试验组饲料, 具体配方及营养水平见表 1。饲料原料粉碎过60目筛, 用绞肉机制成直径1.5 mm的长条状, 切成1.5 mm×2 mm的颗粒状, 风干, 饲料置于-20℃冰柜保存备用。
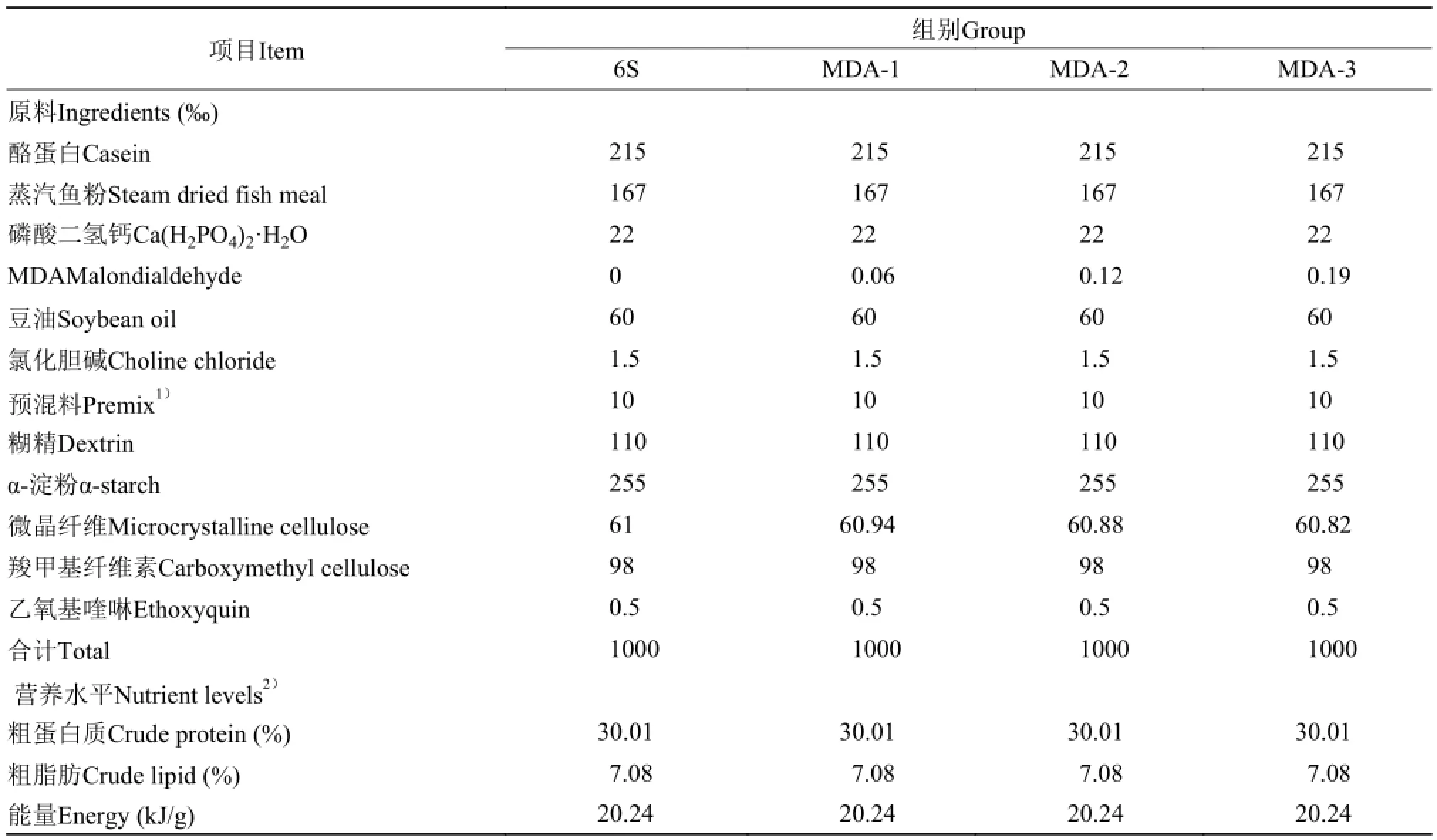
表 1 试验饲料组成及营养水平(干物质%)Tab. 1 Formulation and proximate composition of experiment diets (DM %)
1.3 MDA的制备与添加
MDA的制备方法:根据GB/T 5009.181-2003的方法, 精确量取1,1,3,3-四乙氧基丙烷 (Sigma-Aldrich 公司,浓度≥99%) 31.5 mL, 用95%乙醇溶解后定容至100 mL,搅拌15min, 此时每毫升溶液相当于MDA100 mg, 置于冰箱内保存备用。
MDA的添加:按照每日3%的投喂率配置相应的MDA, 采用现用现配的方式, 快速、均匀地喷洒在饲料当中。MDA的添加量是根据试验“氧化鱼油对草鱼肠道黏膜结构屏障损伤和肝胰脏、肠道胆固醇、胆汁酸合成影响”(已投稿)中氧化鱼油的实际MDA含量设置的。
1.4 饲养管理
饲养实验在浙江一星饲料有限公司养殖基地进行,在面积为5 m×667 m (平均水深1.8 m)的池塘中设置网箱, 网箱规格为(1.0 m×1.5 m×2.0 m)。饲养试验前用6S组饲料驯化一周, 正式饲养时间为72d, 每天7:00和16:00定时投喂, 投饲率为4%。每10天依据投饲量估算鱼体增重并调整投喂率, 记录每天投饲量。每周测定水质一次, 试验期间水温25—33℃, 溶解氧浓度>8.0 mg/L,pH7.8—8.4, 氨氮浓度<0.2 mg/L, 亚硝酸盐浓度<0.01 mg/ L, 硫化物浓度<0.05 mg/L。
1.5 主要试剂
总RNA提取试剂RNAiso Plus, PrimeScriptTMRT Mastetr Mix反转录试剂盒, SYBR Premix Ex TaqTMI都来自TaKaRa公司, 荧光定量PCR扩增引物由上海生工生物技术有限公司合成。
1.6 样品制备与分析
草鱼生长数据的采集与分析 分别捞取各网箱草鱼滤水后称重, 记录尾数, 用于生长速度计算, 并统计各组草鱼增重量和投喂饲料量, 计算饲料系数和特定生长率。计算公式如下。
饲料系数(FCR)=饲料摄入量/(网箱末总重-网箱初总重)
特定生长率(SGR, %)=100×(ln网箱末总重-ln网箱初总重)
血清样品的制备与分析 养殖72d、停食24h后,每网箱随机取出10尾鱼, 采用尾静脉采血法, 取其全血置于离心管中, 常温放置0.5h后, 3000 r/min离心10min制备血清样品, 经液氮速冻后, -80℃保存备用。血清二胺氧化酶(DAO)活性采用南京建成的试剂盒进行测定。血清D-乳酸、内毒素含量采用南京建成的Elisa试剂盒进行测定。血清总胆固醇(TC)、总胆汁酸(TBA)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)含量都采用雅培C800全自动生化分析仪测定。
肠道组织透射电镜样品制备与分析 每网箱取2尾鱼、每组6尾, 于中肠前四分之一处取1—2 cm肠管1段, 纵向剖开用磷酸缓冲液冲洗后, 立即将其投入4%戊二醛中固定, 用于透射电镜分析。透射电镜采用锇酸固定、丙酮脱水, 最后放入胶囊内包埋切片染色, 用日立HT7700透射式电子显微镜观察肠道组织结构并拍照。
草鱼肝胰脏和肠道组织匀浆样品制备与分析 分别取部分新鲜肝胰脏和肠道, 称重后加入10倍体积0.02 mol/ L磷酸缓冲液(pH7.4), 匀浆器10000 r/min匀浆1min, 3000 r/ min冷冻离心10min, 取上清液分装, 液氮速冻后-80℃冰箱保存。肝胰脏和肠道TC、TBA含量都采用雅培C800全自动生化分析仪测定。
草鱼肝胰脏和肠道组织基因样品制备 每网箱随机选取抽过血的3尾鱼活体解剖, 迅速取出内脏团置于冰浴中, 在肝胰脏中间部分和中肠的1/2处各取1.0 cm× 1.0 cm的一块组织于PBS中, 漂洗2—3次后, 一式两份, 迅速装于EP管中, 液氮速冻, 于-80℃保存。
总RNA的提取和反转录cDNA 利用总RNA提取试剂RNAiso Plus按照说明书提取肝胰脏、肠道样品总RNA。取1 μg总RNA为模板, 按照PrimeScriptTMRT Mastetr Mix反转录试剂盒的方法将RNA转录成cDNA,于-20℃保存备用。
qRT-PCR检测肠黏膜细胞间紧密连接蛋白和肝胰脏、肠道胆固醇、胆汁酸合成关键酶基因表达 根据本实验室草鱼肠道转录组测序(RNA-Seq)结果, 运用prime5.0软件设计了6个紧密连接蛋白和2个胆固醇、胆汁酸合成关键酶基因和内参基因 β-actin (GenBank 登录号:DQ211096)的荧光定量正、反向引物(表 2)。

表 2 实时荧光定量引物Tab. 2 Primers used for quantitative real-time PCR
实时定量检测采用CFX96荧光定量PCR仪(Bio-Rad,USA)进行, 反应体系为20 µL:SYBR Premix Ex TaqTMII (TaKaRa)10 µL, 候选引物各1 µL, cDNA 2 µL, 灭菌水6 µL。PCR反应采用两步法, 反应条件为:95℃预变性30s、95℃ 变性5s、60℃ 退火30s,共40个循环。同一样品重复3个反应, 以 β-actin作为参照基因。根据扩增曲线得到的Ct, 计算出目标基因和参照基因 β-actin Ct值的差异ΔCt; 最后计算出不同样品相对于参照样品基因表达倍数2-ΔΔCt, 制作出相对定量的图表。
1.7 数据分析
通过SPSS 21.0进行One-way ANOVA分析, 并进行LSD与Duncan氏比较, 结果以平均值±标准误(mean± SD)表示, 当 P<0.05时, 差异显著。
2 结果
2.1 MDA对草鱼生长性能的影响
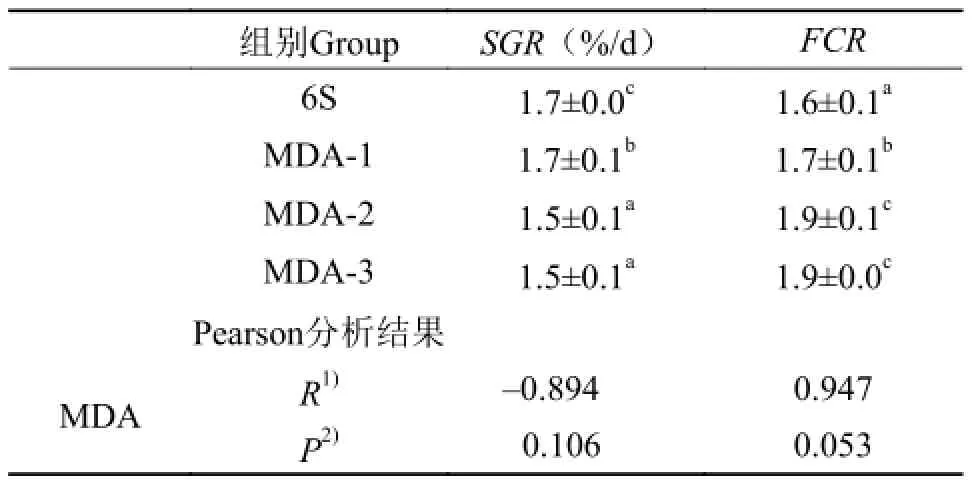
表 3 MDA对草鱼生长的影响及饲料MDA含量与生长指标的相关性分析Tab. 3 Effect of MDA on the growth of Ctenopharyngodon idellus and the correlation analysis of them
由表 3可知, 与对照组相比, 在饲料中添加MDA后,草鱼的SGR都出现显著下降(P<0.05), 而FCR都出现显著升高(P<0.05)。
在相关性分析中, 饲料MDA含量与SGR显示负相关关系的变化趋势, 而与FCR显示正相关关系的变化趋势。
2.2 草鱼血清内毒素、D-乳酸含量和DAO活性显著增加
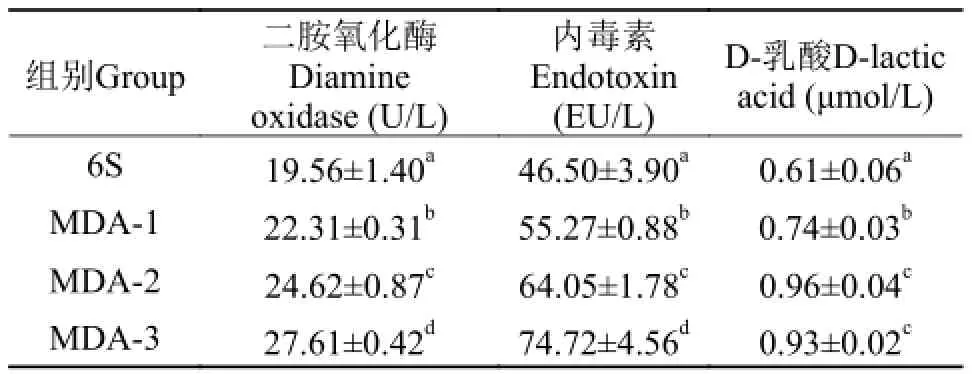
表 4 MDA对草鱼肠道通透性的影响Tab. 4 Effect of MDA on the permeability of grass carp intestine
由表 4可知, 与对照组相比, 在饲料中添加MDA后,血清DAO活性、内毒素含量和D-乳酸含量都出现显著增加(P<0.05), 其中DAO、内毒素含量在MDA-3组达到最大值, 与MDA-1、MDA-2组差异显著。
2.3 MDA使草鱼肠黏膜细胞间紧密连接结构严重损伤
经过72d养殖试验后, 各个试验组草鱼肠黏膜细胞间紧密连接结构的透射电镜图见图版I。图版I-A-D分别为对照组、MDA-1、MDA-2和MDA-3组(图中箭头所示为草鱼肠黏膜细胞间紧密连接结构), 在透射电镜下, 紧密连接结构表现为一条条黑色的致密电子带, 起始于上皮顶端, 从绒毛根部向基底层延伸。由图版I-A可见, 对照组紧密连接结构没有缝隙, 图版I-B-D箭头所示处可以发现紧密连接结构出现缝隙, 并且逐步扩大, MDA-3组紧密连接结构严重受损, 缝隙达到最大。结果显示肠黏膜细胞间紧密连接结构受到严重的损伤。
2.4 MDA诱导草鱼肠黏膜细胞间紧密连接蛋白基因表达显著下调及相关性分析
对组成肠道黏膜细胞间紧密连接结构蛋白质基因表达的检测结果见图 1。由图 1可知, 与对照组相比, 在饲料中添加MDA后, 闭合蛋白Claudin-3、Claudin-15a, 闭锁蛋白Occludin和胞浆蛋白ZO-1、ZO-2、ZO-3基因表达都显著下调(P<0.05)。
由表 5可知, 饲料MDA含量与闭合蛋白Claudin-3、Claudin-15a, 胞浆蛋白ZO-1、ZO-2、ZO-3和闭锁蛋白Occludin基因表达均显示负相关关系的变化趋势。
2.5 MDA对血清、肝胰脏、肠道TC、TBA含量和血清HDL/LDL的影响
MDA对血清TC、TBA、HDL/LDL的影响 由表6可知, 与6S组相比, 在添加MDA后, 血清TC含量都出现不同程度的增加, 在MDA-2和MDA-3组显著增加(P<0.05), 而TBA含量都出现显著下降(P<0.05)。
与6S组相比, 在添加MDA后, 血清HDL/LDL比值都出现显著下降(P<0.05)。
MDA对肝胰脏TC、TBA含量的影响 由表 6可知, 与6S组相比, 在添加MDA后, 肝胰脏TC含量都出现不同程度的增加, 在MDA-2和MDA-3组显著增加(P<0.05)。在MDA组中, 肝胰脏TC含量在4OF组达到最大值, 与MDA-1组差异显著 (P<0.05)。
与6S组相比, 在添加MDA后, 肝胰脏TBA含量都出现不同程度的下降, 但差异不显著(P>0.05)。
MDA对肠道TC、TBA含量的影响 由表 6可知,与6S组相比, 在添加MDA后, 肠道TC含量除在MDA-3组出现下降外, 在MDA-1和MDA-2组显著增加(P<0.05)。
与6S组相比, 在添加MDA后, 肠道TBA含量都出现不同程度的下降, 差异显著(P<0.05)。
2.6 MDA对肝胰脏、肠道HMGCR、CYP7A1基因表达活性的影响及相关性分析
由图 2可知, 在肝胰脏中, 与6S组相比, 在添加MDA后, HMGCR基因表达活性显著上调(P<0.05), 而CYP7A1基因表达活性显著下调(P<0.05)并且HMGCR基因表达活性在MDA-2组达到最大值, 与MDA-1、MDA-3组差异显著(P<0.05)。在肠道中, 与6S组相比, 在添加MDA后, HMGCR、CYP7A1基因表达活性都显著下调(P<0.05)。
由表 7可知, 在肝胰脏中, 饲料MDA含量与HMGCR基因表达活性显示正相关关系的变化趋势, 而与CYP7A1基因表达活性显示负相关关系的变化趋势。在肠道中,饲料MDA含量与HMGCR、CYP7A1基因表达活性均显示负相关关系的变化趋势。

图 1 MDA对草鱼肠黏膜细胞间紧密连接蛋白基因表达的影响Fig. 1 Tight junction protein gene expression of intestine in grass carp under MDA
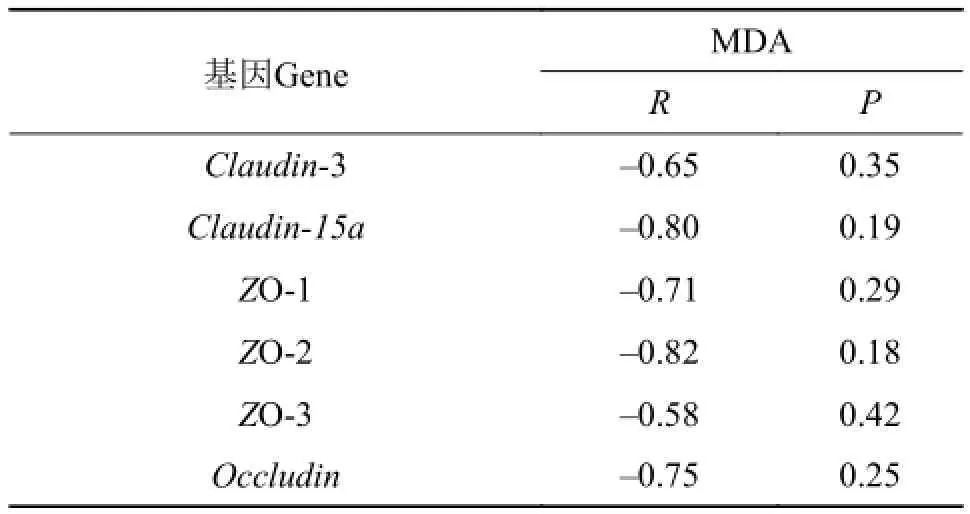
表 5 饲料MDA含量与草鱼肠黏膜细胞间紧密连接蛋白基因表达的相关性分析Tab. 5 Correlation analysis between the content of MDA in diets and tight junction protein gene expression of intestine in grass carp
3 讨论
3.1 MDA对草鱼肠道黏膜结构屏障损伤
MDA使草鱼肠黏膜细胞间紧密连接蛋白基因表达显著下调 如图 3所示, 闭锁蛋白Occludin和闭合蛋白Claudins是构成紧密连接结构的主要跨膜蛋白, 相邻肠黏膜细胞间通过跨膜蛋白Occludin、Claudins的胞外环以“拉链”状相连接, 形成“锁扣”结构, 从而封闭细胞旁间隙, 在维持紧密连接的屏障功能和通透性上起着关键作用[17, 18]。胞浆蛋白ZOs是一类外周膜蛋白, 有三种异构体(ZO-1、ZO-2、ZO-3), 它们一端可以与跨膜蛋白Occludin、Claudins的胞内域相连, 另一端可以与肌动蛋白相结合, 从而将跨膜蛋白与细胞内骨架系统连接起来,构成稳定的紧密连接结构[19]。胞浆蛋白ZOs可以将不同的信号传递到跨膜蛋白Claudins、Occludin, 对紧密连接结构的“开启”与“闭合”进行调控[20]。研究表明, MDA对离体草鱼肠道黏膜细胞膜具有损伤作用, 作用途径可能是促使细胞膜脂质过氧化, 导致细胞凋亡[21]。在氧化豆油水溶物对离体草鱼肠道黏膜细胞损伤的研究中提到,氧化豆油水溶物中的MDA可能是对细胞产生损伤的重要物质之一[22]。在本试验中, 如图 1所示, 在饲料添加MDA后, 闭锁蛋白Occludin、闭合蛋白Claudin-3、Claudin-15a基因表达都出现不同程度的下调, 且差异显著(P<0.05), 胞浆蛋白ZO-1、ZO-2、ZO-3基因表达显著下调(P<0.05), 同时表 5结果显示, 饲料MDA含量与闭锁蛋白Occludin, 闭合蛋白Claudin-3、Claudin-15a, 胞浆蛋白ZO-1、ZO-2、ZO-3基因表达均显示负相关关系的变化趋势。上述结果表明, 饲料MDA减少了闭锁蛋白Occludin、闭合蛋白Claudin-3、Claudin-15a的生成能力,削弱了胞浆蛋白ZOs对紧密连接“锁扣”结构的“闭合”调控, 通过打开紧密连接“锁扣”结构的方式, 导致肠黏膜细胞间紧密连接结构的破坏和肠黏膜通透性的增加, 损伤肠黏膜屏障。
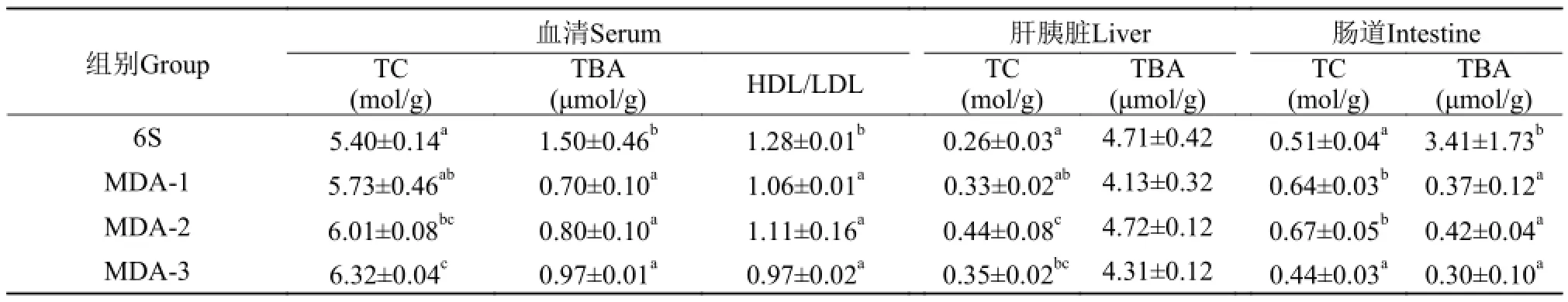
表 6 草鱼血清、肝胰脏、肠道TC、TBA含量和血清HDL/LDL的变化Tab. 6 The effect of contents of TC, TBA in serum, liver and intestine and the value of HDL/LDL in serum
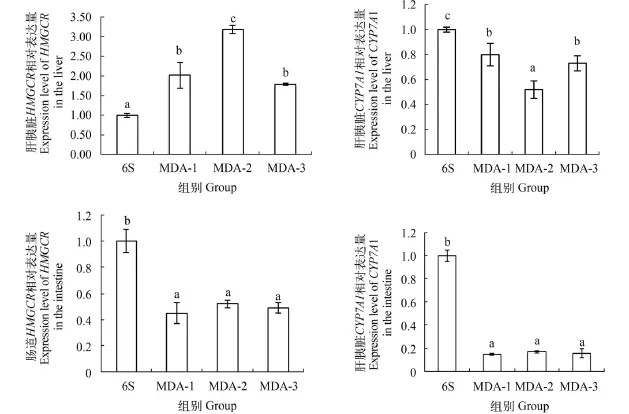
图 2 MDA对草鱼肝胰脏和肠道HMGCR、CYP7A1基因表达活性的影响Fig. 2 The relative gene expression of HMGCR, CYP7A1 of liver and intestine in grass carp under MDA
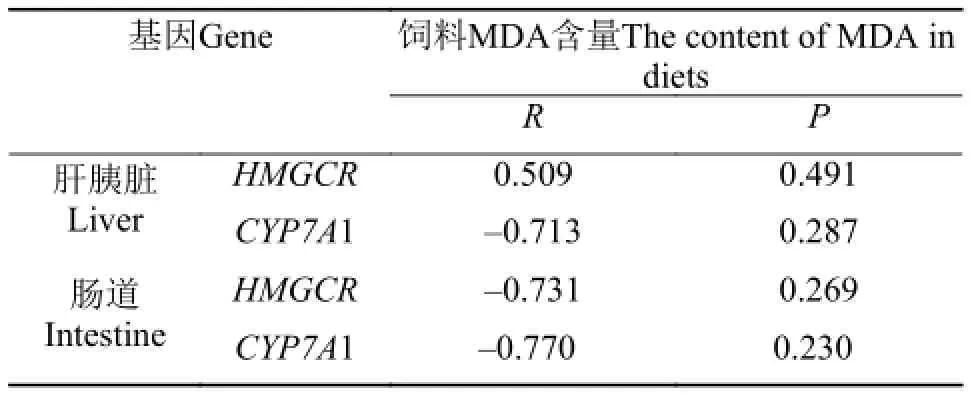
表 7 肝胰脏、肠道HMGCR、CYP7A1基因表达活性相关性分析Tab. 7 Correlation analysis of HMGCR and CYP7A1 in the liver and intestine
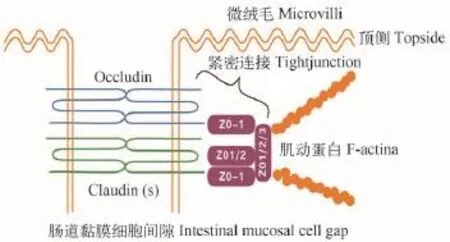
图 3 肠上皮细胞间的紧密连接示意图Fig. 3 The tight junctions of intestinal epithelial intercellular
MDA使草鱼肠黏膜通透性显著增加 肠黏膜通透性的升高主要通过肠黏膜细胞通路和肠黏膜细胞间通路通透性增加来实现[23]。其中, DAO活性、D-乳酸和内毒素含量经常作为判断肠黏膜通透性和肠黏膜屏障功能的指标。
DAO是具有高度活性的细胞内酶, 该酶在小肠黏膜上层绒毛含量高, 活性强, 在其他组织含量少, 活性低。当肠黏膜细胞受损、肠黏膜通透性增加后, 胞内释放大量的DAO会通过肠黏膜屏障而进入血液, 使血浆DAO活性升高[24]。D-乳酸主要是细菌发酵的代谢产物, 正常情况下很少被吸收。当肠黏膜细胞受损时, 肠黏膜通透性增加, 肠道中细菌产生大量D-乳酸会通过受损黏膜细胞进入血液, 使血浆D-乳酸水平升高[25]。所以血浆DAO活性和D-乳酸含量可作为反映肠道黏膜损害程度和通透性变化的重要指标。当肠道屏障被破坏时, 肠黏膜通透性增加, 大量的内毒素可通过肠黏膜细胞间通路、肠黏膜细胞微绒毛的细胞膜通路, 进入血液引发内毒素血症。因此, 内毒素的通透性可反映肠黏膜屏障的功能[26]。有研究表明, 腹泻、感染和手术等多种应激状态均可导致暂时或长时间的肠黏膜屏障损伤, 表现为肠黏膜通透性增加、细菌和毒素移位等[27]。表 4结果显示, 在饲料中添加MDA后, 血清DAO活性、D-乳酸和内毒素含量都出现显著增加(P<0.05), 表明饲料MDA破坏了肠黏膜细胞和肠黏膜细胞间紧密连接结构, 即肠黏膜屏障遭到严重损伤, 肠黏膜通透性显著增加。
MDA使紧密连接结构遭到破坏 作为肠黏膜细胞间通路的关键结构, 紧密连接结构只允许离子和可溶性的小分子通过, 大分子物质及微生物难以通过, 通过紧密连接结构的“开启”与“闭合”实现肠黏膜细胞间通路的“开启”与“闭合” 。Schmitz等[28]研究发现, 紧密连接发生变化会使肠黏膜屏障受损。如图版I所示, 对照组紧密连接结构没有出现缝隙, MDA-1、MDA-2组紧密连接结构开始出现缝隙, 并且逐步扩大, MDA-3组紧密连接结构严重受损, 缝隙达到最大。这些结果表明, 在饲料中添加MDA后, 紧密连接的“锁扣”结构会被打开, 紧密连接结构遭到破坏, 导致肠黏膜通透性增加。这也为肠黏膜细胞间紧密连接蛋白基因表达显著下调导致肠道紧密连接结构的破坏和肠黏膜通透性的增加提供了很好的证据。
3.2 MDA对草鱼生长性能的影响
肠道作为鱼体消化和吸收的主要场所, 承担着从外界获取物质、能量的重任。当鱼类的肠道屏障结构遭到破坏时, 必然会引起其摄食饲料的利用率降低, 从而阻碍其正常的生长、发育[29, 30]。上述结果表明, 饲料MDA会减少闭锁蛋白Occludin、闭合蛋白Claudin-3、Claudin-15a的生成能力, 削弱胞浆蛋白ZOs对紧密连接“锁扣”结构的“闭合”调控, 使紧密连接结构遭到破坏, 导致肠黏膜通透性增加和肠黏膜屏障损伤, 最终破坏肠道屏障功能的完整性, 其结果发展的趋势应该会降低草鱼的生长速度。由表 3结果显示, 在饲料中添加MDA后, 草鱼SGR显著下降(P<0.05), 而FCR显著升高(P<0.05), 同时在相关性分析中, 饲料MDA含量与SGR显示负相关关系的变化趋势, 而与FCR显示正相关关系的变化趋势, 表明MDA降低了草鱼的生长性能, 这也为上述分析提供了有力的证据。
3.3 MDA对草鱼肝胰脏、肠道胆固醇、胆汁酸合成影响
MDA对草鱼肝胰脏胆固醇、胆汁酸合成影响肝胰脏不仅是鱼类重要的代谢和解毒器官, 也是机体合成胆固醇、胆汁酸的主要场所。Manwaring等[31]在小鼠体内首次发现MDA会与肝脏中的蛋白质发生交联, 形成一种高分子量、水溶性的荧光物质。Klamerth等[32]用MDA饲喂的大鼠肝脏DNA模板活性也明显下降。本试验中, 在饲料添加MDA后, 肝胰脏胆固醇合成关键酶HMGCR显著上调(P<0.05), 而胆汁酸合成关键酶CYP7A1基因表达活性显著下调(P<0.05), 同时由表 7结果可知, 饲料MDA含量与HMGCR基因表达活性显示正相关关系的变化趋势, 而与CYP7A1基因表达活性显示负相关关系的变化趋势, 表明MDA会导致肝细胞以乙酰辅酶A为原料合成胆固醇的能力增强和以胆固醇为原料合成胆汁酸的能力减弱, 由表 6结果显示, 血清 HDL/ LDL的比值在丙二醛组都出现显著下降(P<0.05), 也暗示着外周组织胆固醇转入肝细胞的能力在增强, 此时肝胰脏胆固醇含量应该会出现增加, 而胆汁酸含量会出现减少的趋势。Horton等[33]研究表明, MDA可以与大鼠肝脏线粒体中的乙醛脱氢酶高效且不可逆的结合从而影响线粒体的功能, 进而损伤肝脏。由表 6结果显示, 在饲料中添加MDA后, 血清、肝胰脏胆固醇含量都出现显著增加(P<0.05), 而胆汁酸含量都出现不同程度的减少, 这不仅为上述分析提供了很好的证据, 也暗示在MDA的损伤作用下, 肝胰脏可能需要更多的胆固醇以满足其生理代谢的需要, 而胆汁酸的需求可能会出现供给不足的局面。
本试验还发现, 肝胰脏HMGCR基因表达活性在MDA-2组达到最大值, 与MDA-1、MDA-3组差异显著(P<0.05), 肝胰脏胆固醇含量在MDA-2组达到最大值, 与MDA-1组差异显著 (P<0.05), 表明在摄食含有0.12 g/kg MDA的饲料后, MDA对肝胰脏胆固醇合成的影响最大。
MDA对草鱼肠道胆固醇、胆汁酸合成影响 肠道作为机体与外界接触的最大界面, 在选择性吸收营养物质和防御有毒有害物质入侵等方面发挥着屏障功能。Saito等[34]的研究发现, 脂质过氧化产物会改变肠道细胞膜表面的酶活性甚至使其活性丧失。在本试验中, 在饲料中添加MDA后, 草鱼肠道胆固醇合成关键酶HMGCR、胆汁酸合成关键酶CYP7A1基因表达活性都显著下调(P<0.05), 同时表 7结果显示, 饲料MDA含量与HMGCR、CYP7A1基因表达活性均显示负相关关系的变化趋势, 表明MDA会导致肠道细胞的胆固醇和胆汁酸合成能力减弱。有研究表明, MDA对离体草鱼肠道黏膜细胞膜具有损伤作用, 作用途径可能是促使细胞膜脂质过氧化, 导致细胞凋亡[21]。在氧化豆油水溶物对离体草鱼肠道黏膜细胞损伤的研究中提到, 氧化豆油水溶物中的MDA可能是对细胞产生损伤的重要物质之一[22]。在本试验中, 由表6结果显示, 肠道胆固醇含量显著增加(P<0.05), 而肠道胆汁酸含量显著减少(P<0.05), 暗示在MDA损伤作用下,肠道可能需要更多的胆固醇以满足其生理代谢的需要,胆汁酸的需求可能会出现供给不足的局面。
综上所述, MDA会引起草鱼肠道黏膜结构屏障遭到破坏和肝胰脏、肠道胆固醇、胆汁酸合成能力受到影响, 而MDA作为鱼油氧化的最主要产物, 是否会通过损伤草鱼肠道黏膜结构屏障的方式, 引起肝胰脏、肠道胆固醇、胆汁酸合成能力的变化呢, 这有待后面进一步研究。
4 结论
饲料MDA会减少闭锁蛋白Occludin、闭合蛋白Claudin-3、Claudin-15a的生成能力, 削弱胞浆蛋白ZOs对紧密连接“锁扣”结构的“闭合”调控, 使紧密连接结构遭到破坏, 导致肠黏膜通透性增加和肠黏膜屏障损伤, 最终破坏肠道屏障功能的完整性, 从而降低草鱼的生长性能。
随着饲料MDA含量的增加, 肝胰脏胆固醇合成能力在增强, 其中以含有0.12 g/kg MDA的饲料对肝胰脏胆固醇合成的影响最大, 而胆汁酸合成能力在减弱, 致使肝胰脏、血清胆固醇含量增加和胆汁酸含量减少, 预示着肝胰脏可能需要更多的胆固醇以满足生理代谢的需要, 而胆汁酸可能出现供给不足的局面。
随着饲料MDA含量的增加, 肠道胆固醇、胆汁酸合成能力都在减弱, 而此时肠道胆固醇含量增加和胆汁酸含量减少, 预示着肠道同样需要更多的胆固醇以满足生理代谢的需要, 而胆汁酸可能出现供给不足的局面。
[1]Om A D, Umino T, Nakagawa H, et al. The effects of dietary EPA and DHA fortification on lipolysis activity and physiological function in juvenile black sea bream (Bleeker) [J]. Aquaculture Research, 2001, 32(suppl.):255—262
[2]Cao J M, Tian L X, Chen Z, et al. Effect of dietary fatty acias on growth and tissue chemical composition of grass carp, Ctenopharyngodon idellus [J]. Journal of South China University of Technology (Natural Science), 1996,12(Suppl): 149—154 [曹俊明, 田丽霞, 陈竹, 等. 饲料中不同脂肪酸对草鱼生长和组织营养成分组成的影响.华南理工大学学报(自然科学版), 1996, 12(增刊):149—154]
[3]Esterbauer H, Schauur J S, Zollner H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes [J]. Free Radical Biology and Medicine,1991, 11(1): 81—128
[4]Chen Q, Le G W, Shi Y H, et al. Advance in the damage of reactive oxygen species on the gastrointestinal tract in animal [J]. China Animal Husbandry & Veterinary Medicine, 2006, 33(11): 106—108 [陈群, 乐国伟, 施用晖, 等.氧自由基对动物消化道损伤及干预研究进展. 中国畜牧兽医, 2006, 33(11): 106—108]
[5]Monahan F J, Gray J I, Asghar A, et al. Effect of dietary lipid and vitamin E supplementation on free radical production and lipid oxidation in porcine muscle miscrosomal fractions [J]. Food Chemistry, 1993, 46(1): 1—6
[6]Epstein M D, Tchervenkov J I, Alexander J W, et al. Increased gut permeability following burn trauma [J]. Archives of Surgery, 1991, 126(2): 198—200
[7]Tsukita S, Furuse M. Occludin and claudins in tight junction strands: leading or supporting players [J]? Trends Cell Biology, 1999, 9(7): 268—273
[8]Laura L M, Christina M, Van I, et al. Molecular physiology and pathophysiology of tight junctions I. tight junction structure and function: lessons from mutant animals and proteins [J]. American Journal of Physiology Gastrointest Liver Physiology, 2000, 279(2): G250—G254
[9]Furuse M, Sasaki H, Fujimoto K, et al. A single gene product, claudin-1 or -2, reconstitutes tight junction strands and recruits occludin in fibroblasts [J]. Journal of Cell Biology, 1998, 143(2): 391—401
[10]Keno B H, Schafer S, Kuhn C, et al. Symplekin, a novel type of tight junction plaque protein [J]. Cell Biology,1996, 134(4): 1003—1018
[11]Weber E, Berta G, Tousson A, et al. Expression and polarized targeting of a rab3 isoform in epithetial cells [J]. Journal of Cell Biology, 1994, 125(3): 583—594
[12]Gao Z G, Qin H L. The biological functions of intestinal epithelial tight junctions and its role in intestinal barrier [J]. Parenteral & Enteral Nutrition, 2005, 12(5): 299—302 [高志光, 秦环龙. 肠上皮细胞紧密连接的生物学功能及在肠屏障中的作用. 肠外与肠内营养, 2005, 12(5):299—302]
[13]Huang L, Jiang Z Y, Lin Y C, et al. Influence of oxidized fish oil on intestinal mucosal immune response in neonatal piglets and the use of soybean isoflavones as an intervention measure [J]. Acta Zoonutrimenta Sinica, 2011,23(5): 799—806 [黄琳, 蒋宗勇, 林映才, 等. 饲喂氧化鱼油对新生仔猪肠道黏膜免疫应答的影响及大豆异黄酮的干预作用. 动物营养学报, 2011, 23(5): 799—806]
[14]Chen Q, Yue G W, Shi Y H, et al. Advance in the damage of reactive oxygen species on the gastrointestinal tract in animal [J]. China Animal Husbandry & Veterinary Medicine, 2006, 33(11): 106—108 [陈群, 乐国伟,施用晖, 等. 氧自由基对动物消化道损伤及干预研究进展. 中国畜牧兽医, 2006, 33(11): 106—108]
[15]Huang Y W, Ye Y T, Cai C F, et al. The effect of the gene expression in metabolism of cholesterol synthesis pathway after intestine injuried on Ctenopharyngodon idellus [J]. Journal of Nanjing Agricultural University,2015, 38(5): 497—503 [黄雨薇, 叶元土, 蔡春芳, 等. 肠道损伤对草鱼胆固醇代谢通路基因表达的影响. 南京农业大学学报, 2015, 38(5): 497—503]
[16]Ye Y T, Cai C F, Xu F, et al. Feeding grass carp (Ctenopharyngodon idellus) with oxidized fish oil upregulates the gene expression in the cholesterol and bile acid synthesis pathway in intestinal mucosa [J]. Acta Hydrobiologica Sinica, 2015, 39(1): 94—104 [叶元土, 蔡春芳, 许凡, 等. 灌喂氧化鱼油使草鱼肠道黏膜胆固醇胆汁酸合成基因通路表达上调. 水生生物学报, 2015, 39(1):94—104]
[17]Sakakibara A, Furuse M, Saitou M, et al. Possible involvement of phosphorylateion of occludin in tight junction formation [J]. The Journal of Cell Biology, 1997,137(6): 1393—1401
[18]Itoh M, Furuse M, Morita K, et al. Direct binding of three tight junction associated MAGUKs, ZO-1, ZO-2, and ZO-3, with the COOH termini of claudins [J]. Journal of Cell Biology, 1999, 147(6): 1351—1363
[19]Keon B H, Schafer S, Kuhn C, et al. Symplekin, a novel type of tight junction plaque protein [J]. Cell Biology,1996, 134(4): 1003—1018
[20]Zhong Y, Saitoh T, Minase T, et al. Monoclonal antibody 7H6 reacts with a novel tight junction associated protein distinct from ZO-1, cingulin and ZO-2 [J]. Journal of Cell Biology, 1993, 120(2): 477—483
[21]Yao S B, Ye Y T, Cai C F, et al. Damage of MDA on intestinal epithelial cells in vitro of grass carp (Ctenopharyngodon idellus) [J]. Acta Hydrobiologica Sinica, 2015,39(1): 137—146 [姚仕彬, 叶元土, 蔡春芳, 等. 丙二醛对离体草鱼肠道黏膜细胞的损伤作用. 水生生物学报,2015, 39(1): 137—146]
[22]Yao S B, Ye Y T, Cai C F, et al. The water soluble matter from oxidized soybean oil damages dissociated intestinal epithelial cells of grass carp (Ctenopharyngodon idellus) [J]. Acta Hydrobiologica Sinica, 2014, 38(4):690—698 [姚仕彬, 叶元土, 蔡春芳, 等. 氧化豆油水溶物对离体草鱼肠道黏膜细胞的损伤作用. 水生生物学报, 2014, 38(4): 690—698]
[23]Lu L, Walker W A. Pathologic and physiclogic interactions of bacteria with the gastrointestinal epithlium [J]. American Journal of Clinical Nutrition, 2001, 73(suppl):1124S—1130S
[24]Li J Y, Lü Y. The significance of changes and in diamine oxidase activity in intestinal injury after trauma [J]. Chinese Critical Care Medicine, 2000, 12(8): 482—484[黎君友, 吕艺. 二胺氧化酶在创伤后肠道损伤中变化及意义. 中国危重病急救医学, 2000, 12(8): 482—484]
[25]Sun X Q, Fu X B, Zhang R, et al. The effect of intestinal ischemia - reperfusion injury on intestinal permeability [J]. Journal of Traumatic Surgery, 1999, 1(4): 208—210[孙晓庆, 付小兵, 张蓉, 等. 大鼠肠缺血-再灌流损伤对肠黏膜通透性的影响. 创伤外科杂志, 1999, 1(4):208—210]
[26]Lenz A, Franklin G A, Cheadle W G. Systemic inflammation aftertrauma [J]. Injury, 2007, 38(12): 1336—1345
[27]Liu H P, Hu C H, Xu Y. Effects of early weaning on intestinal permeability and tight junction protein occludin mRNA expression levels of piglets [J]. Chinese Journal of Animal Nutrition, 2008, 20(4): 442—446 [刘海萍, 胡彩虹, 徐勇. 早期断奶对仔猪肠通透性和肠上皮紧密连接蛋白Occludin mRNA表达的影响. 动物营养学报,2008, 20(4): 442—446]
[28]Schmitz H, Bamteyer C, Fromm M, et al. Utered fight junction structure contributes to the impaired epithetial barrier function in ulcerative colitiss [J]. Gastroenterology, 1999, 116(2): 301—309
[29]Qiu Y, Ye Y T, Cai C F, et al. Effect of raffinose on growth performance and intestinal morphology of grass carp (Ctenopharyngodon idellus) [J]. Cereal & Feed Industry, 2012, (1): 53—56 [邱燕, 叶元土, 蔡春芳等. 棉子糖对草鱼生长性能与肠道黏膜形态的影响. 粮食与饲料工业, 2012, (1): 53—56]
[30]Zhang J X. The effects of soybean protein source on growth performance and intestinalimmune of juvenile Jian carp (Cyprinus carpio Var. Jian) [D]. Sichuan Agricultural University, 2003 [张锦秀. 大豆蛋白源对幼建鲤生长性能及肠道免疫的影响. 四川农业大学, 2003]
[31]Manwaring J D, Csallany A S. Malondialdehyde-containing proteins and their relationship to vitamin E [J]. Lipids,1988, 23(7): 651—655
[32]Klamerth O L, Levinsky H. Template activity in liver DNA from rats fed with malondialdehyde [J]. FEBS Letters, 1969, 3(3): 205—207
[33]Horton A A, Packer L. Interactions between malondialdehyde and rat liver mitochondria [J]. Journal of Gerontology, 1970, 25(3): 199—204
[34]Saito M, Nakatsugaw K. Incrteased susceptibility of liver to lipidperoxidation after ingestion of a high fish oil diet [J]. International Journal for Vitamin and Nutrition Research, 1994, 64(2): 144—151
THE DAMAGE OF INTESTINAL MUCOSA BARRIER STRUCTURE AND THE EFFECT OF CHOLESTEROL AND BILE ACID SYNTHESIS PATHWAY IN THE LIVER AND INTESTINE UNDER MDA IN GRASS CARP (CTENOPHARYNGODON IDELLA)
HUANG Yu-Wei, YE Yuan-Tu, CAI Chun-Fang, WU Ping, CHEN Ke-Quan, WU Tao, XU Deng-Hui, PENG Kan,LIN Xiu-Xiu and LUO Qi-Gang
(Key Laboratory of Aquatic Nutrition of Jiangsu Province, Suzhou University, Suzhou 215123, China)
丙二醛; 肝胰脏; 肠道; 胆固醇; 胆汁酸; 紧密连接
MDA; Liver; Intestine; Cholesterol; Bile acid; Tight junction
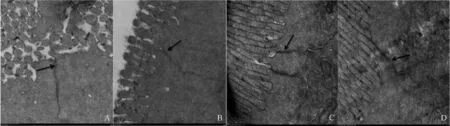
图版 I 草鱼肠道紧密连接的透射电镜图(×12000)Plate I TEM micrographs of the junction structures in grass carp
Q959; S965.1
A
1000-3207(2016)04-0869-10
10.7541/2016.112
2015-02-16;
2015-08-12
国家自然科学基金资助项目(31172417); 苏州市应用基础研究项目(SYN201316)资助 [Supported by the National Natural Science Foundation Project of China (31172417); Applied Basic Research Project of Suzhou (SYN201316)]
黄雨薇(1989—), 女, 四川宜宾; 硕士研究生; 研究方向为水产动物营养与饲料。E-mail: 1041372540@qq.com
叶元土, 教授; Tel: 0512-65880179; E-mail: yeyt@suda.edu.cn
