灰枣3个优系果实膨大期的光合及荧光特性研究
2016-07-23冯会丽吴正保郭佳欢宋锋惠马合木提阿不来提
冯会丽,吴正保,郭佳欢,宋锋惠,马合木提·阿不来提
(1新疆农业大学林学与园艺学院,乌鲁木齐 830052;2新疆林科院经济林研究所,乌鲁木齐 830000)
灰枣3个优系果实膨大期的光合及荧光特性研究
冯会丽1,2,吴正保2,郭佳欢1,宋锋惠2,马合木提·阿不来提2
(1新疆农业大学林学与园艺学院,乌鲁木齐830052;2新疆林科院经济林研究所,乌鲁木齐830000)
摘要:【目的】研究灰枣3个优系(‘灰实2’、‘灰实7’和‘灰实8’)果实膨大期的光合及叶绿素荧光特性。【方法】以灰枣的3个优系为研究对象,灰枣为CK,采用单因素完全随机区组设计,测定灰枣及其3个优系叶片的光合-光响应生理参数及叶绿素荧光日变化。【结果】‘灰实2’和‘灰实7’的LCP较低而LSP较高、Rd值较高;‘灰实8’的LCP较高而LSP较低、Rd值较低;‘灰实2’的Pmax和AQE均最大,其Rubisco活性和电子传递速率较高。灰枣及其3个优系在强光和高温胁迫下,都会出现光抑制现象;‘灰实2’和‘灰实7’受到光抑制后恢复快且耐受性良好,二者的表观光合电子传递速率优于‘灰实8’;‘灰实7’的光化学反应启动速率和光能利用率最高,‘灰实2’次之,‘灰实8’最低;‘灰实8’受到高温强光胁迫后的生理调节机制和自我保护机制启动较慢。【结论】‘灰实2’对弱光的利用能力最强,对强光的适应性最好,具备丰产潜力;‘灰实7’的光合能力略次于‘灰实2’,优于CK和‘灰实8’。‘灰实2’叶片中PSII天线色素内的最初原子密度及叶绿素含量高于CK和其他优系,具有很好的光能利用潜力;‘灰实2’和‘灰实7’对强光及高温胁迫的反应较‘灰实8’和CK要灵敏;‘灰实8’对弱光的利用能力较弱且对强光适应性差。
关键词:灰枣;优系;光响应;叶绿素荧光;日变化
0引 言
【研究意义】光合作用是绿色植物将光能转变为有机物化学能的能量转换过程,是植物体内最重要的化学反应,同时也是影响植物生长快慢和农作物产量的重要因素,长久以来成为品种选育工作者备受青睐的指标之一[1]。通过光合作用的光响应曲线,运用相关的光合参数,反映植物在不同光照条件下生存能力以及对不断变化的环境条件的适应能力。叶绿素荧光信号能实时反映植物自身的生理状态及其对外界环境因素的响应,其灵敏度高、可靠性强,现已被广泛用于植物生态和植物抗逆性、高光效或抗逆品种筛选及光抑制和光破坏防御机制等方面的研究[2-3]。灰枣(Zizyphusjujuba‘Huizao’)质地密合、含水量低、糖分高、风味佳,含有多种微量元素和较多的药用成分,具有很高的食疗价值和多种保健功效[4-6]。新疆灰枣在20世纪70年代初自河南新郑引种以来,得天独厚的光热资源,各项经济性状均优于原产区,尤其在南疆各地得到大面积栽培[7]。目前,灰枣已成为阿克苏地区主栽品种之一[8]。随着规模化的发展,灰枣在生产中品质和风味出现了问题,灰枣优系选育面临新挑战。【前人研究进展】已有研究表明:决定灰枣品质和经济效益的关键期在果实膨大期,该期内枣果发育最活跃,是细胞加速分裂和迅速生长期,也是枣树生物量及产量形成的关键时期,此时需要消耗大量的营养物质[9-10]。王森[11]、王振亮等[12-13]研究选育‘金粟枣’、‘沧冬1号’和‘沧冬2号’芽变品种。目前,有关灰枣的良种选育研究尚集中在植物学特性、生长结果习性和果实经济性状等方面,关于灰枣及其优系光合-光响应特征及叶绿素荧光参数日变化方面的比较研究未见报道。【本研究切入点】通过对灰枣及其3个优系果实膨大期的光合-光响应及叶绿素荧光参数日变化测定与分析,结合光合有效辐射及气温日变化参数,研究不同优系的光合及荧光生理生态特性。【拟解决的关键问题】为灰枣的优系选育和推广应用提供技术参考及理论依据。
1材料与方法
1.1试验地概况
试验地点位于新疆阿克苏地区实验林场二队,地理坐标41°10′49″N、80°20′22″E,海拔1 108.3 m,属暖温带大陆性气候,降雨量少,蒸发量大,气候干燥、昼夜温差大,年平均降水量42.4~94.4 mm,年蒸发量达2 000~2 900 mm,年平均气温在9.9~11.5℃,无霜期169~247 d,光热资源丰富,年平均≥10℃积温为3 953℃,年日照时数达2 800~3 831.35 h,年太阳总辐射量为5 340~6 220 MJ/(m2·a),风沙浮尘天气较多,主要集中在春、夏两季。
1.2材 料
供试材料为灰枣及新疆林科院初选的3个灰枣优系,暂命名‘灰实2’、‘灰实7’和‘灰实8’,树龄8 a,树势中强,株行距2 m×3 m,东西行向栽植,未间作,漫灌。
1.3方 法
1.3.1试验设计
试验于2014年4月~2015年9月进行,以枣树品种(系)为处理,设‘灰实2’、‘灰实7’和‘灰实8’及CK共4个样本,每处理选取3株调查样株,于果实膨大期(8月中旬)测定光合有效辐射(photosynthetically active radiation,PAR)日变化、气温日变化及叶片的光合生理参数、叶绿素荧光日变化。
1.3.2测定项目
1.3.2.1光合有效辐射及气温日变化
采用美国Li-COR公司生产的LI-191型线状量子传感器测定当地PAR。选择晴朗天气,于08:00~20:00测定光合有效辐射值;采用广州公司生产的JM222型便携式手持气温计记录试验地气温,两项指标同步测定,2 h一次,3个重复,取平均值。
1.3.2.2光合-光响应生理参数
采用Ciras-2便携式光合仪及自带的红蓝光源测定光合生理参数。分别选取灰枣及其3个优系的南向2年生枝枣股的枣吊上健康叶片进行测定。选择晴朗天气,于10:30~13:30测定在光合光量子通量密度(photosynthetic photo flux density,PPFD)、17个梯度下的净光合速率Pn(net photosynthetic rates),绘制Pn-PPFD曲线。测定时,叶片温度、叶室内部CO2浓度、空气相对湿度分别维持在(30±1)℃、80%±1%和420 μmol/(m2·s)。不同品种(系)每次测定选择3片叶,取平均值。
光响应采用目前常用的非直角双曲线模型[14-15],如式(1)。
Pn(I)=
式中,Pn为净光合速率,Pmax为最大净光合速率,α为表观量子效率(μmol·CO2/μmol·PPFD),I为叶片接受到的PPFD(mol·CO2/μmol),Rd为暗呼吸速率,θ为光响应曲线弯曲程度的凸度。根据该模型用SPSS 17.0非线性回归,得到Pn-PPFD的拟合曲线,计算Pmax、Rd及θ。当I为0~200 μmol/(m2·s)时,对Pn-PPFD进行直线回归,得出线性回归方程Pmax=AQE×LSP-Rd,并据此方程得到表观量子效率(apparent quantum efficiency,AQE)、光补偿点(light compensation point,LCP)和光饱和点(light saturation point,LSP)。线性直线的斜率为表观量子效率;当Pn=0时,光合有效辐射为光合作用的光补偿点;当Pn=Pmax时,光合有效辐射为光合作用的光饱和点。
1.3.2.3叶绿素荧光生理参数
采用FMS-2便携式荧光仪测定叶绿素荧光日变化,测定对象及重复的设定同光合生理参数。选择晴朗天气,于08:00~20:00测定叶片的荧光生理参数,每2 h测定一次。测定时,叶片需经过12~16 h暗适应,再在荧光暗反应夹下适应20 min,然后测定叶片的初始荧光(initial fluorescence,F0)和最大荧光(maximum fluorescence,Fm);同一叶片在持续的自然光下适应20 min,测得稳态荧光(steady-state fluorescence,Fs)和光下最大荧光(maximum fluorescence in the light,Fm’),仪器自动记录得出其PSII最大光化学产量(maximal photochemical of PSII in the dark,Fv/Fm)、实际光化学效率(actual photochemical efficiency,ФPSII)、表观光合电子传递效率(the apparent electron transport rate,ETR)。
1.4数据统计
采用Excel 2007对数据整理和基本处理,用SPSS 17.0统计软件进行方差分析、回归分析,采用Origin 8.0作图软件制图。
2结果与分析
2.1光合有效辐射及气温日变化
研究表明,试验地的光合有效辐射及气温日变化曲线表现一致,均呈单峰曲线,自08:00起不断上升,14:00达到峰值,之后不断下降。表明随着太阳高度的变化,光合有效辐射和气温也随之不断发生变化。图1
2.2灰枣及其优系光合生理参数
3个优系及CK的光照利用能力各异,但对PPFD的响应趋势表现一致。当PPFD小于或等于300 μmol/(m2·s)时,净光合速率Pn上升趋势明显,先呈直线上升,之后上升趋势变缓,在1 000~1 100 μmol/(m2·s)处接近最大净光合速率。其中‘灰实8’与CK的光响应曲线几乎重合,‘灰实7’的曲线略低于CK,‘灰实2’明显高于CK。图2
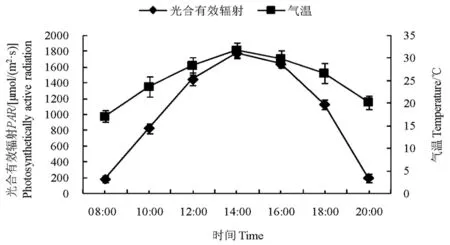
图1 光合有效辐射及气温日变化
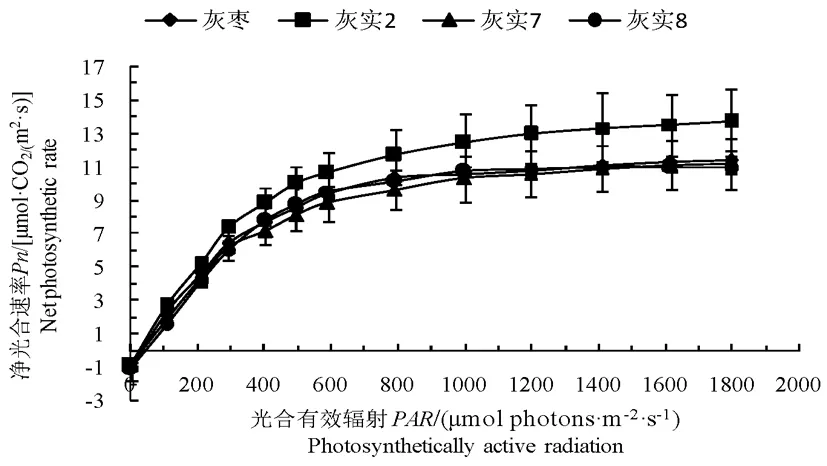
图2 灰枣及其优系果实膨大期的光响应曲线

品种SpeciesLSPμmol/(m2·s)LCPμmol/(m2·s)Pmaxμmol/(m2·s)AQE(μmol·CO2/μmol·PPFD)Rdμmol/(m2·s)灰枣Zizyphusjujuba‘Huizao’1000.13±71.09ABb34.57±13.62Aab11.78±0.46Ab0.027±0.003ABb0.949±0.487Aa‘灰实2’Thegrayexperiment21185.51±30.17Aa28.75±4.41Ab14.32±1.66Aa0.032±0.002Aa0.921±0.154Aa‘灰实7’Thegrayexperiment71066.80±88.07ABab31.78±3.17Aab11.57±1.51Ab0.026±0.003ABb0.821±0.171Aa‘灰实8’Thegrayexperiment8944.76±115.75Bb44.20±0.96Aa11.57±0.43Ab0.025±0.002Bb1.091±0.088Aa
注:表中数值为平均值±标准差, 同列不同大小写字母表示在0.01和0.05水平上差异显著
Note:Values is means±S.D. Different capital and small letters in a column indicate significant different at 0.01 or 0.05 levels
研究表明,LCP由小到大为:‘灰实2’<‘灰实7’
2.3灰枣及其优系叶绿素荧光生理参数
2.3.1初始荧光F0日变化
F0是PSII反应中心全部开放时的荧光水平,反映了PSII的电子受体最大程度氧化态时的荧光[16-17]。PSII天线的热耗散增加常导致F0的降低,而PSII反应中心的破坏或可逆失活则引起F0的增加,可根据F0的变化推测反应中心的状况和可能的光保护机制[18-19]。
研究表明,3个优系与CK的叶片F0日变化趋势表现一致,均呈双峰曲线,都在12:00达第1个峰值,之后显著下降,14:00到谷底后又继续上升,16:00出现第2个峰值,然后开始下降。说明在一天内,12:00和16:00时,PSII反应中心的开放程度及天线色素到PSII反应中心的激发能传递速率最大,生理活性最强;14:00时可能受到光抑制或者高温胁迫,导致生理活性下降。F0整体排序为:‘灰实2’>‘灰实7’>‘灰实8’>CK。表明3个优系叶片中PSII天线色素内的最初原子密度及叶绿素含量均高于CK,且最初原子密度及叶绿素含量的排序为:‘灰实2’>‘灰实7’>‘灰实8’。图3
2.3.2最大荧光Fm日变化
Fm是PSII反应中心处于完全关闭时的荧光产量,可以反映通过PSII的电子传递情况。Fm日变化呈现波动下降趋势,3个优系均在10:00和14:00 出现波谷,CK的波谷只出现在14:00。整体看,3个优系的Fm值全天高于CK,其中‘灰实8’的Fm降幅最大,光抑制现象最明显,‘灰实2’受到的光抑制现象最不明显。3个优系中对强光的耐受力为:‘灰实2’>‘灰实7’>‘灰实8’>CK。图4
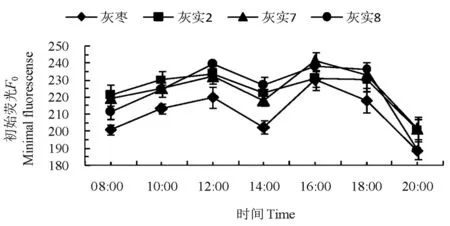
图3 灰枣及其优系果实膨大期的初始荧光日变化
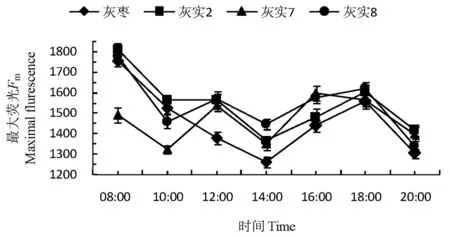
图4 灰枣及其优系果实膨大期的最大荧光日变化
2.3.3最大光化学效率Fv/Fm日变化
灰枣及其优系的Fv/Fm日变化整体呈现出先下降后上升的趋势。‘灰实2’和‘灰实7’的最大光化学效率14:00达最低值、‘灰实8’和CK在16:00达到最低值。当最大光化学效率到达最低值时,表明受到了外界环境因子的胁迫,叶片的PSII反应中心受到威胁,光化学效率下降,光合作用受到抑制。研究表明,灰枣及其优系的最大光化学效率达到最低值后的恢复程度各异,20:00时‘灰实2’、‘灰实7’、‘灰实8’和CK分别恢复到08:00时的98.74%、100.08%、99.72%和99.29%。16:00以后,随着光强和温度的下降,灰枣及其优系叶片PSII反应中心内原初光能转化效率上升趋势明显。表明‘灰实2’和‘灰实7’对强光及高温胁迫的反应比‘灰实8’和CK灵敏,‘灰实8’和CK在外界胁迫后的生理调节机制和自我保护机制启动较慢,容易遭受高温强光伤害。图5
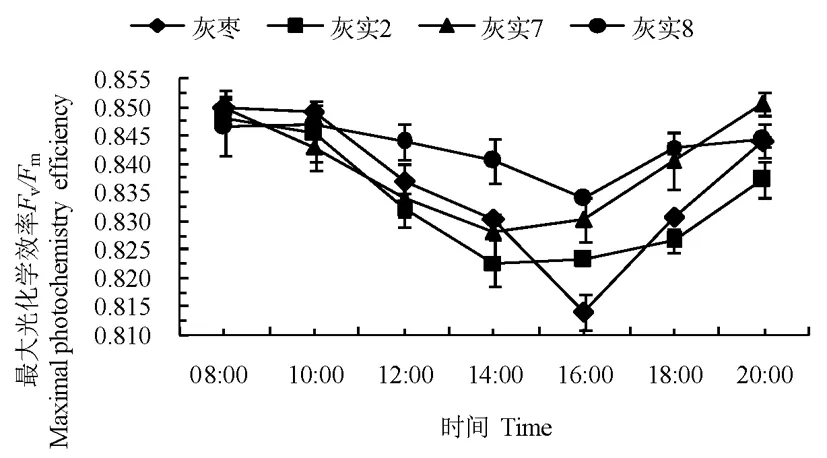
图5 灰枣及其优系果实膨大期最大光化学效率日变化
2.3.4实际光化学效率ФPSII日变化
ФPSII反映叶片用于光合电子传递的能量占所吸收光能的比例,是PSII反应中心部分关闭时的光化学效率,其值大小可以反映PSII反应中心的开放程度,可作为植物叶片光合电子传递速率快慢的相对指标。
3个优系与CK叶片的ФPSII日变化呈现先下降后增长的趋势。‘灰实7’、‘灰实8’和CK均在12:00时达到最低值,且‘灰实7’的降幅最小,‘灰实2’在14:00时达到最低值,降幅最大。3个优系与CK的实际光化学效率整体排序为:‘灰实2’>CK>‘灰实7’>‘灰实8’。表明‘灰实2’的光合电子传递能力强、吸收的光能被用于光化学反应的份额大,对强光的耐受力强,有利于提高光能转化效率,为暗反应的光合碳同化积累更多能量,以促进碳同化的高效运转和有机物的积累,为高产奠定物质基础。图6
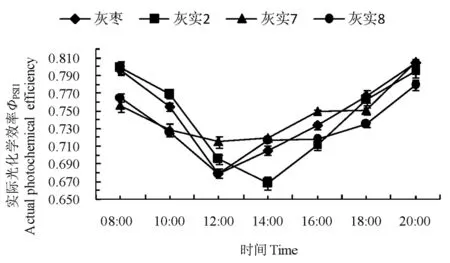
图6 灰枣及其优系果实膨大期实际光化学效率日变化
2.3.5表观光合电子传递速率ETR日变化
ETR反映实际光强条件下的表观电子传递速率。3个优系与CK的ETR值日变化呈现单峰变化趋势,08:00~12:00都处于平缓增长阶段,说明12:00之前均受到弱光或低温的胁迫,导致光合电子传递过程受到抑制,12:00以后,随着光合有效辐射和气温的上升,其表观电子传递速率显著上升,至16:00时达到峰值,之后随着光合有效辐射和气温的下降而降低。整体来看,ETR值由大到小为:CK>‘灰实7’>‘灰实2’>‘灰实8’。表明ETR值是随着光合有效辐射和气温的变化而变化的,‘灰实7’的光化学反应启动速率和光能的利用率高,‘灰实2’次之,‘灰实8’最低。图7
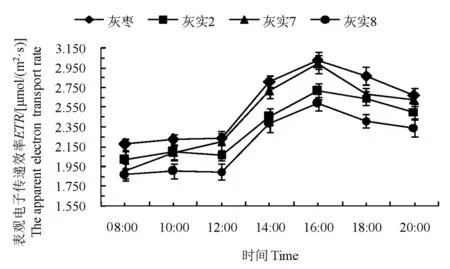
图7 灰枣及其优系果实膨大期表观电子传递效率日变化
3讨 论
3.1灰枣及其优系的光合特征的差异
LCP越低,说明植物利用弱光能力越强;LCP越高,说明植物对光照强度的要求越高,对强光的耐受能力越强。LCP和LSP同时较低的植物耐阴性很强;LCP低而LSP较高的植物能适应多种光照环境;LCP较高而LSP较低的植物,应栽植于侧方遮阴或部分时段阴蔽的环境;LCP、LSP均较高的植物则为喜光的阳性植物[20-22]。研究中,相对于CK,‘灰实2’、‘灰实7’的LCP较低且LSP较高,说明二者对光能的利用区间较宽,能适应多种光照环境;‘灰实8’的LCP较高而LSP较低,说明其对强光和弱光的耐受能力均较差。Pmax反映植物对强光的利用能力,它决定于Rubisco活性和电子传递速率。‘灰实2’有较高的Rubisco活性和电子传递速率,说明‘灰实2’光合干物质生产效率高,为产量的提高奠定了良好的物质基础,丰产潜力较大。AQE反映植物对弱光的利用能力,值越大,植物吸收与转换光能的色素蛋白复合体可能越多,利用弱光的能力越强。该研究发现,AQE值排序为‘灰实2’>‘灰实7’>‘灰实8’,表明‘灰实2’具有生长快、适应性广、推广应用潜力大等生态生物学特性。Rd反映植物消耗光合产物的速率,值越大,生理活性越高。有报道指出,暗呼吸速率的降低并不是由于光强的抑制,而是由于暗呼吸CO2在光下被重新回收利用,其回收利用率随着光强的增加而变化,从而导致暗呼吸速率的变化[23]。‘灰实7’和‘灰实2’的Rd值较高,生理活性也较高。据此推断,‘灰实8’为灰枣优系中不喜光、也不耐阴的植物,这与其具有较高光补偿点和较低光饱和点的结果相一致。
3.2 灰枣及其优系的叶绿素荧光特征的差异
叶绿素是捕获光能的物质基础,植物叶片叶绿素荧光可反映其自身的光合能力。Fv/Fm是一个快速而灵敏的光抑制指标,其值越低说明植物发生光抑制的程度越高[24-25]。魏亦农等[26]研究认为,Fv/Fm下降的同时F0增加,说明光抑制现象是PSII反应中心的可逆性失活造成的。而徐崇志等[27]研究发现,Fv/Fm下降的同时F0随之下降,说明光抑制现象主要是热耗散造成的。研究发现,所有材料叶片的Fv/Fm日变化均呈“V”型变化趋势。‘灰实2’和‘灰实7’对强光及高温胁迫的反应灵敏,自我保护能力强;‘灰实8’和CK在外界胁迫下的生理调节机制和自我保护机制启动较慢,更容易遭受光抑制带来的伤害。研究还发现,灰枣3个优系的叶片中PSII天线色素内的最初原子密度及叶绿素含量均高于灰枣,说明对光合光量子的吸收和捕获能力强。其中‘灰实2’比其他2个优系的光能转化率高及对强光的耐受力强,为丰产栽培奠定了物质基础。
目前在叶绿素荧光参数日变化中,研究最多的是关于叶片的光抑制[28]。试验所研究的3个灰枣优系的耐光抑制能力不同,这与张守仁[29]对不同杨树优系和徐德聪[30]对于不同美国山核桃优系的研究结果一致。通过研究灰枣优系的光合及叶绿素荧光特性,对其的光合能力及生长适应性进行一个综合评估,为灰枣的良种选育及栽培推广提供科学依据。但在研究中,强光与高温并存,这两个因素对于不同优系间叶片的影响及作用机制没有区分,这在研究中有一定的局限性,需在今后进一步探讨。
4结 论
4.1灰枣的3个优系中,‘灰实2’对弱光的利用能力最强,对强光的适应性最好,整体光合能力较优,适应性广,具备丰产潜力,‘灰实7’次之。
4.2‘灰实2’叶片中PSII天线色素内的最初原子密度及叶绿素含量高于‘灰实7’、‘灰实8’和CK,具有很好的光能利用潜力。
4.3‘灰实2’和‘灰实7’对强光及高温胁迫的反应较‘灰实8’和CK灵敏。
4.4‘灰实7’的光化学反应启动速率高,‘灰实2’次之,‘灰实8’最低,而对光能的利用率则为‘灰实7’最高,‘灰实2’次之,‘灰实8’最低。
参考文献(References)
[1]李泽,谭晓风,袁军,等.4个油桐品种光合特性的日变化研究[J].中国农学通报,2013,29(25):12-15.
LI Ze, TAN Xiao-feng, YUAN Jun. (2013). The study on diurnal changes of photosynthetic characteristics of 4 cultivars of tung tree (Vernicia fordii) [J].ChineseAgriculturalScienceBulletin, 29(25):12-15.(in Chinese)
[2]彭方仁,朱振贤,谭鹏鹏,等.NaCl胁迫对5个树种幼苗叶片叶绿素荧光参数的影响[J].植物资源与环境学报,2010,19(3):42-47.
PENG Fang-ren, ZHU Zhen-xian, TAN Peng-peng, et al. (2010). Effect of NaCl stress on chloropliyll fluorescence parameter in leaf of five tree species seeding [J].JournalofPlantResourcesandEnvironment, 19(3):42-47. (in Chinese)
[3]温国胜,田海涛,张明如,等.叶绿素荧光分析技术在林木培育中的应用[J].经济林研究,2006,17(10):1 973-1 977.
WEN Guo-sheng, TIAN Hai-tao, ZHANG Ming-ru, et al. (2006). Applocation of chlorophyll fluorescence analysis in forest tree cultivation [J].ChineseJournalofAppliedEcology, 17(10):1,973-1,977.(in Chinese)
[4]Chen, C. F., Lee, J. F., Wang, D., Shen, C. Y., Shen, K. L., & Lin, M. H. (2010). Water extract of zizyphus jujube attenuates ischemia/reperfusion-induced liver injury in rats (pp106).TransplantationProceedings, 42(3):741-743.
[5]Li J W, Ding S D, Ding X L. (2005). Comparison of antioxidant capacities of extracts from five cultivars of Chinese jujube.ProcessBiochemistry, 40(11):3,607-3,613.
[6]Shen, X., Tang, Y., Yang, R., Yu, L., Fang, T., & Duan, J. A. (2009). The protective effect of zizyphus jujube fruit on carbon tetrachloride-induced hepatic injury in mice by anti-oxidative activities.JournalofEthnopharmacology, 122(3):555-560.
[7]张萍,史彦江,宋锋惠,等.南疆灰枣主要营养品质性状的变异及相关性研究[J].果树学报,2011,28(1):77-81.
ZHANG Ping, SHI Yan-jiang, SONG Feng-hui, et al. (2011). Investigation on variation and correlation of the main nutrition quality traits of Ziziphus Jujuba cv. Huizao from south of Xinjiang [J].JournalofFruitScience, 28(1):77-81. (in Chinese)
[8]李欢,李建贵,秦韵婷,等.微气候因子对南疆CK坐果和果实品质的影响 [J].果树学报,2015,32(6):1 161-1 169.
LI Huan, LI Jian-gui, QIN Yun-ting, et al. (2015). Effects of orchard microclimatic conditions on the fruit setting and quality of Ziziphus jujuba Mill. 'Huizao' in south Xinjiang [J].JournalofFruitScience, 32(6):1,161-1,169.(in Chinese)
[9]彭勇,田福忠,张小燕,等.冬枣果实膨大期追施尿素对叶片氮代谢的影响[J].西北农业学报,2007,16(5):163-166.
PENG Yong, TIAN Fu-zhong, ZHANG Xiao-yan, et al. (2007). Effects of topdressing urea on nitrogen metabolism of leaf during fruit swelling of winter jujube [J].ActaAgriculturaeBoreali-occidentalisSinica, 16(5):163-166.(in Chinese)
[10]李晓彬,汪有科,赵春红,等.水分调控对梨枣果实品质与投入产出效益的影响分析[J].中国生态农业学报,2011,19(4):818-822.
LI Xiao-bin, WANG You-ke, ZHAO Chun-hong, et al. (2011). Effect of regulated irrigation on input-output benefits of pear jujube [J].ChineseJournalofEco-Agriculture, 19(4):818-822.(in Chinese)
[11]王森,谢碧霞,钟秋平,等.枣新品种'中秋酥脆枣'[J].园艺学报,2009,36(5):771-771.
WANG Sen, XIE Bi-xia, ZHONG Qiu-ping, et al. (2009). A new Zizyphus jujube cultivar 'Zhongqiu Sucui' [J].ActaHorticulturaeSinica, 36(5):771-771.(in Chinese)
[12]王振亮,肖家良,韩会智,等.短枝型芽变枣新品种-'沧冬1号' [J].园艺学报,2012,39(6):1 213-1 214.
WANG Zhen-liang, XIAO Jia-liang, HAN Hui-zhi, et al. (2012). A new Chinese jujube cultivar 'Cangdong1' with bud mutation of short branch [J].ActaHorticulturaeSinica, 39(6):1,213-1,214.(in Chinese)
[13]王振亮,韩会智,肖家良,等.短枝型芽变枣新品种-'沧冬2号'的选育[J].果树学报,2012,29(5):956-957,712.
WANG Zhen-liang, HAN Hui-zhi, XIAO Jia-liang, et al. (2012). A new Chinese jujube cultivar with short branch-'Cangdong2' [J].JournalofFruitScience, 29(5): 956-957,712.(in Chinese)
[14]Thornley JHM. (1976).MathematicalModelsinPlantPhysiology. London: Academic Press:86-110.
[15]EUGENE I. RABINOWITCH. (1951). Photosynthesis and related processes.SoilScience, 72(6):831-1,191.
[16]Strasser, B.J, Strasser RJ (1995). Measuring fast fluorescence transients to address environmental questions: the JIP-Test. In: Mathis P (ed) Photosynthesis: from light to biosphere. Kluwer Academic Publishers, The Netherlands, pp 977-980.
[17]Krause, G. H., & Weis, E. (1991). Chlorophyll Fluorescence and Photosynthesis: The Basics.AnnualReviewofPlantPhysiology, 42(4):313-349.
[18]卞佃侠,朱建军,柏新富.脱水方式对单叶蔓荆叶绿素荧光特性的影响[J].安徽农业科学,2010,38(6):3 214-3 216.
BIAN Dian-xia, ZHU Jian-jun, BAI Xin-fu. (2010). Effect of dehydration method on chlorophyll fluorescence characteristics in Vitex trifolia [J].AnhuiAgric.Sci., 38(6): 3,214-3,216.(in Chinese)
[19]段仁燕,吴甘霖,黄振波.草莓叶绿素荧光参数日变化的研究[J].生物学杂志,2009,26(5):69-70,73.
DUAN Ren-yan, WU Gan-lin, HUANG Zhen-bo. (2009). Daily change of chlorophyll fluorescence of Fragaria ananassa Duch [J].JournalofBiology, 26(5):69-70,73.(in Chinese)
[20]王建华,任士福,史宝胜,等.遮荫对连翘光合特性和叶绿素荧光参数的影响[J]. 生态学报,2011,31(7):1 811-1 817.
WANG Jian-hua, REN Shi-fu, SHI Bao-sheng, et al. (2011). Effects of shades on the photosynthetic characteristics and chlorophyll fluorescence parameters of Forsythia suspense [J].ActaEcologicaSinica, 31(7):1,811-1,817.(in Chinese)
[21]梁文斌,聂东伶,吴思政,等.遮荫对短梗大参苗木光合作用及生长的影响[J]. 生态学杂志,2015,34(2):413-419.
LIANG Wen-bin, NIE Dong-lin, WU Si-zheng, et al. (2015). Effects of shading on the growth and photosynthesis of Macropanax rosthornii seedings [J].ChineseJournalofEcology, 34(2):413-419. (in Chinese)
[22]朱万泽,王金锡,薛建辉,等.四川桤木光合生理特性研究[J].西南林学院学报,2001,21(4):196-204.
ZHU Wan-ze, WANG Jin-xi, XUE Jian-hui, et al. (2001). Studies on the Physiological Characteristics of Photosynthesis of Alnus cremastogyme [J].JournalofSouthwestForestryCollege, 21(4):196-204.(in Chinese)
[23]冯玉龙,曹坤芳,冯志立,等.四种热带雨林树种幼苗比叶重、光合特性和暗呼吸对生长光环境的适应[J].生态学报,2002,22(6):901-910.
FENG Yu-long, CAO Kun-fang, FENG Zhi-li, et al. (2002). Acclination of Lan in a Mass Per Unit Area, Photosynthetic Characteristics and Dark Respiration to Growth Light Regines in Four Tropical Rainforest Species [J].ActaEcologicaSinica, 22(6): 901-910. (in Chinese)
[24]温国胜,田海涛,张明如,等.叶绿素荧光分析技术在林木培育中的应用[J].应用生态学报,2006,17(10):1 973-1 977.
WEN Guo-sheng, TIAN Hai-tao, ZHANG Ming-ru, et al. (2006). Application of chlorophyll fluorescence analysis in forest tree cultivation [J].ChineseJournalofAppliedEcology, 17(10): 1,973-1,977.(in Chinese)
[25]张永霞,李国旗,闰伟兄.红麻和自麻叶绿素荧光日变化研究初探[J].中国野生植物资源,2007,26(5):50-53.
ZHANG Yong-xia, LI Guo-qi, YAN Wei-xiong. (2007). Primary research on diurnal changes of the chlorophyll fluorescence parameters of Apocyrrum verwtum L.and Apocyrrum henderserri Hookf [J].ChineseWildPlantResources, 26(5): 50-53.(in Chinese)
[26]魏亦农,孔广超,曹连莆.新小黑麦1号光合速率及叶绿素荧光特性的研究[J].麦类作物学报,2002,22(4):91-93.
WEI Yi-nong, KONG Guang-chao, CAO Lian-pu. (2002). Study on photosynthetic rate and chlorophyll fluorescence character in new triticale [J].JournalofTriticeaeCrops, 22(4):91-93. (in Chinese)
[27]徐崇志,全绍文,朱玲,等.不同核桃品种叶绿素荧光动力学参数比较研究[J].中国农学通报,2015,31(7):6-12.
XV Chong-zhi, QUAN Shao-wen, ZHU Ling, et al. (2015). Studies on the Diurnal Variation of Chlorophyll Fluorescence Parameters in Different Walnut Cultivars [J].ChineseAgriculturalScienceBulletin, 31(7):6-12.(in Chinese)
[28]焦念元,李亚辉,李法鹏,等.间作玉米穗位叶的光合和荧光特性[J].植物生理学报,2015,51(7):1 029-1 037.
JIAO Nian-yuan, LI Ya-hui, LI Fa-peng, et al. (2015). Photosynthesis and chlorophyll fluorescence characteristics in ear leaves of intercropped maize [J].PlantPhysiologyJournal, 51(7):1,029-1,037. (in Chinese )
[29]张守仁,高荣孚.光胁迫下杂种杨优系光合生理生态特性的研究[J].植物生态学报,2000,24(5):528-533.
ZHANG Shou-ren, GAO Rong-fu. (2000). Ecophysiological characteristics of photosynthesis of hybrid polar clones under light stress [J].ActaPhytoecologicaSinica, 24(5):528-533. (in Chinese)
[30]徐德聪,吕芳德,潘晓杰.叶绿素荧光分析技术在果树研究中的应用[J].经济林研究,2003,21(3):88-91.
XU De-cong, LV Fang-de, PAN Xiao-jie. (2003). Application of chlorophyll fluorescence analysis technique to research of fruit trees [J].EconomicForestResearches, 21(3):88-91. (in Chinese)
Fund project:Supported by special funds for national forestry public welfare industry research (201304701-1) and the basic science and technology research support funds of non-profit research institutions of Xinjiang Uygur Autonomous Region public welfare scientific research institutes for basic scientific research funding projects "Research into jujube germplasm resources introduction and growth adaptability"
doi:10.6048/j.issn.1001-4330.2016.06.006
收稿日期(Received):2016-01-05
基金项目:国家林业公益行业科研专项(201304701-1);自治区公益性科研院所基本科研业务经费资助项目“枣优良种质资源引进及生长适应性研究”
作者简介:冯会丽(1988-),女,山西晋城人,硕士研究生,研究方向为生态学,(E-mail)550112219@qq.com 通讯作者(Cotresponding author):宋锋惠(1967-),男,山东聊城人,研究员,博士,研究方向为果实栽培、林木遗传育种,(E-mail)sfh1111@126.com
中图分类号:S665.1,S722.5
文献标识码:A
文章编号:1001-4330(2016)06-1014-09
Research of Photosynthetic and Fluorescence Characteristics at Fruit Expanding Period of Three Excellent Strains ofZizyphusjujuba'Huizao'
FENG Hui-li1,2, WU Zheng-bao2, GUO Jia-huan1, SONG Feng-hui2, Mahemuti Abulaiti2
(1.CollegeofForestryandHorticulture,XinjiangAgriculturalUniversity,Urumqi830052,China;2.ForestResearchInstituteofXinjiangAcademyofForestry,Urumqi830000,China)
Abstract:【Objective】 In order to observe the photosynthetic and fluorescence characteristics at fruit expanding period of three excellent strains ('gray experiment 2', 'gray experiment 7',and 'gray experiment 8') of Zizyphus jujuba 'Huizao'.【Method】By taking three excellent strains of Zizyphus jujuba 'Huizao' as the research object, Zizyphus jujuba 'Huizao' as control, single factors completely randomly block experiment was designed. The photosynthetic-light response physiological parameters and daily variation of chlorophyll fluorescence of leaves of Zizyphus jujuba 'Huizao' and its three excellent strains were measured.【Result】The results showed that the light compensation point was lower and light saturation point was higher of 'gray experiment 2' and 'gray experiment 7', both of which had a higher dark respiration rate. 'Gray experiment 8' had a higher light compensation point, but the light saturation point and dark respiration rate was lower. The maximum net photosynthetic rate and apparent quantum efficiency were the largest in 'gray experiment 2', the Rubisco activity and electron transfer rate were relatively high. Zizyphus jujuba 'Huizao' and its three excellent strains presented photo inhibition phenomenon under the stress of strong light and high temperature. After the photo inhibition, 'gray experiment 2' and 'gray experiment 7' recovered faster and well tolerated, and the apparent photosynthetic electron transport rate was better than that of 'gray experiment 8'. 'Gray experiment 7' had the highest photochemical reaction starting rate and light energy utilization, followed by 'gray experiment 2', and 'gray experiment 8' was the lowest. The physiological regulatory mechanism and self-protection mechanism starting rate of 'gray experiment 8' was slower, when suffering high temperature and strong light stress.【Conclusion】'Gray experiment 2'had the strongest ability to use weak light, and the best adaptability to strong light with high yield potential. The photosynthetic ability of 'gray experiment 7' was slightly inferior to 'gray experiment 2', which was better than that of CK and 'gray experiment 8'. The initial atomic density in PSII antenna pigment and chlorophyll content of 'gray experiment 2' leaves were higher than those of the CK and other excellent strains and it had very good light utilization potential. The reaction of 'gray experiment 2' and 'gray experiment 7' to strong light and high temperature stress was more sensitive than 'gray experiment 8' and CK. The ability of 'gray experiment 8' to use weak light was feeble, and the strong light adaptability was poor.
Key words:Zizyphus jujuba 'Huizao'; excellent strains; light response; chlorophyll fluorescence; diurnal variation
