蜜蜂嗅觉研究进展
2016-07-19谢翠琴聂红毅苏松坤福建农林大学蜂学学院福州350000
谢翠琴 聂红毅 苏松坤(福建农林大学蜂学学院,福州350000)
蜜蜂嗅觉研究进展
谢翠琴聂红毅苏松坤
(福建农林大学蜂学学院,福州350000)
摘要:蜜蜂嗅觉影响蜜蜂交尾、哺育和采集等行为,对蜂群的繁衍和发展具有重要作用。蜜蜂触角上分布着大量的嗅觉感觉器,能感知挥发性分子、气味和激素等,因此是蜜蜂最重要的嗅觉器官,也是蜜蜂参与嗅觉交流的重要组成部分。综述蜜蜂触角、嗅觉感受器、嗅觉相关蛋白及嗅觉传导机制等,能为蜜蜂后续的研究和蜜蜂触嗅觉的应用提供参考。
关键词:蜜蜂;触角;嗅觉
自然界中昆虫众多,它们依靠化学、视觉、嗅觉、听觉和触觉等方式进行信息交流,适应环境的选择;信息交流可以促进昆虫个体相互协作,也能提高昆虫求偶、觅食和抵御天敌的能力,从而增强昆虫个体与群体的生存和繁衍能力。嗅觉在昆虫中具有重要作用,参与昆虫择偶、选择寄主和寻找食物源等。蜜蜂是社会性昆虫,嗅觉与蜜蜂生命活动的各个方面息息相关,如:工蜂通过嗅觉感知蜂王物质有助于维持蜂群的稳定,还能通过嗅觉识别植物分子等;雄蜂利用灵敏的嗅觉追寻处女王,即嗅觉参与蜜蜂交尾,表明嗅觉对蜜蜂适应环境和种群繁衍具有重要意义。而蜜蜂触角上分布着众多的嗅觉感觉器,因此是蜜蜂最重要的嗅觉器官,也是蜜蜂参与嗅觉交流的重要组成部分。对蜜蜂嗅觉进行综述,能为蜜蜂后续的研究和蜜蜂触嗅觉的应用提供参考。
1 蜜蜂触角
自然界中昆虫众多,它们依靠化学、视觉、嗅觉、听觉和触觉等方式进行信息交流,适应环境的选择。触角上分布着众多的感受器,是昆虫重要的感觉器官,不仅拥有触觉、嗅觉和味觉的功能,还能感受CO2、温度和湿度等。嗅觉感受器主要分布于昆虫的触角,其次是在下颚须和下颚唇上,因此在昆虫触角的众多功能中,嗅觉是昆虫最重要的功能之一。
蜜蜂头部有1对膝状触角,由柄节、梗节和鞭节组成;柄节是触角基部的第一节,梗节是第二节,鞭节是由若干鞭小节组成的,蜂王和工蜂的鞭节有10节,而雄蜂的有11节。在蜜蜂触角上有板形、坛形、锥形、钟形、腔形和毛形等多种感觉器,其中的板形感觉器与嗅觉最相关;每个板形感觉器中含有多条感觉神经元(图1-1)。雄蜂、工蜂和蜂王触角上的板形感受器的数目分别为16685、2232和1505,所以雄蜂触角上的板形感受器最多,大约是工蜂的7.5倍,说明不同级型的蜜蜂,其嗅觉能力有所差异[1]。中华蜜蜂雄蜂触角上的板形感受器比意大利蜜蜂的少,而中华蜜蜂和意大利蜜蜂工蜂触角上的板形感觉器没有显著差异[2]。
探索蜜蜂触角是一个复杂的过程,早期主要观察触角的基本结构和形态,然后利用扫描电镜对蜜蜂触角上感觉器进行分类并研究其功能,随后运用触角电位技术研究感觉器中电信号的变化。随着分子技术的发展,蜜蜂触角中嗅觉相关基因和蛋白逐步被研究,同时分析它们各自的结构和功能。随着科学技术的发展与蛋白质组学的成熟,蛋白质组越来越多应用于蜜蜂触角蛋白质组的差异鉴定。
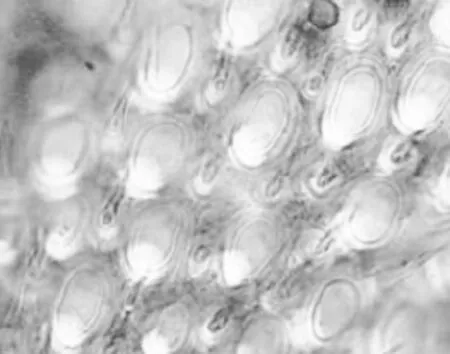
图1-1(A)光学显微镜下蜜蜂板形感觉器[3]
图1-1(B)蜜蜂板形感觉器的横切面[3]
2 蜜蜂嗅觉系统
蜜蜂在长期进化中演化出复杂的嗅觉系统,其主要功能由感觉器、嗅觉感受蛋白、嗅觉受体、感觉神经元膜蛋白和气味降解酶等相互协调运行产生。蜜蜂能通过嗅觉识别植物气味分子、主要蜂王物质成分(9-氧代-癸二烯酸、和9-羟基-癸二烯酸)、脂肪酸幼虫信息素成分、报警信息素成分和其它信息素进行化学交流,这对蜂群的繁殖和稳定具有重要作用。昆虫嗅觉感受传递是非常复杂的过程,目前尚未完全研究清楚;对蜜蜂嗅觉系统的研究主要是从感受器、感受蛋白、嗅觉受体、感觉神经元膜蛋白和气味降解酶分块逐步研究的,目前尚未深入研究蜜蜂的嗅觉系统。
2.1嗅觉感觉蛋白
蜜蜂嗅觉感觉蛋白分为气味结合蛋白(odorantbinding proteins,OBPs)和化学感觉蛋白(chemosensory proteins,CSPs)两种,都是球形的水溶性α-螺旋小蛋白(10~30 kDa),有2个或3个二硫键。OBP和CSP基因不仅在嗅觉器官中表达,还在其它组织中表达,说明除嗅觉功能外,它们可能还具有其它功能。
2.1.1气味结合蛋白
昆虫中的OBPs的分子量通常在13~16 kDa范围,是α-螺旋水溶性酸性小蛋白,在辅细胞中合成后被运送到感觉器淋巴液中,识别与结合气味分子,然后将气味分子运输至嗅觉神经元树突膜的周围。
OBPs家族根据基因序列可以分为6个亚家族Classic,Minus-C,Plus-C,Dimer,Doble和Double-Minus-C。也可以根据蛋白功能将OBPs分为五类:(1)信息素结合蛋白(pheromone binding proteins,PBPs),此类蛋白在触角中表达丰富,能与主要信息素结合;(2)普通气味结合蛋白(general odorant binding proteins,GOBPs),能结合花香化合物[4];(3)触角结合蛋白(Antennal Binding Protein,ABP);(4)化学感觉相关亲脂配体结合蛋白(Chemical-sense-Related Lipophilic-ligand-Binding Protein);(5)双翅目唾液蛋白家族(member of the Diptera salivary protein family,D7)。在蜜蜂中,根据蛋白功能把OBPs分为PBPs、GOBPs、ASP(触角特异性蛋白antennal-specific protein,ASP)。又根据基因序列分为两类:一类是传统的OBPs(1-13),含6个半胱氨酸,与其它昆虫相同,以Cys(I)-Cys(III),Cys(II)-Cys(V),Cys (IV)-Cys(VI)成对交联;另一类是C-minus OBPs(缺少第二或第五个半胱氨酸以及它们之间相连的二硫键),其中的OBP15-21含有4个半胱氨酸,而OBP14具有5个半胱氨酸[3,5]。
在1998年Danty等人通过质谱和N端序列分析,推测在西方蜜蜂中有25个OBPs类似蛋白,并将划分出在触角中特异表达蛋白;后来经过基因组测序、原位杂交等方法鉴定出西方蜜蜂中有21个OBP基因[6,7];与许多昆虫相比,蜜蜂含有较少的OBP基因。西方蜜蜂的21个OBP基因进行neighbour-joining tree分析表明,OBP13和C-minus OBPs组成最大一单系,它们都在15号染色体上,并且它们的序列相似度高达48%,因此推断OBP13和C-minus OBP基因都是由含6个半胱氨酸共同的祖先演化而来;相反的C-minus组外的OBP基因的序列相似度低,难以发现它们的起源[8]。
对东方蜜蜂OBPs的研究比较零散,陆续克隆表达研 究 了 ASP1,ASP2,OBP3,OBP5,OBP9,OBP11和OBP12,其中的ASP1属于PBP;OBP11能与工蜂信息素法呢醇结合,同时也能与植物芳香物质结合[9-16]。
西方蜜蜂的 OBP3(除触角)、OBP16、OBP17、OBP18、OBP19、OBP20和OBP21在成年蜜蜂各个部分都表达;OBP7和OBP9仅在蜂王的卵巢中表达;OBP14和OBP15在幼虫期表达,在蛹期不表达;OBP10在蛹期表达,并在刚出房时表达达到最高峰;蜜蜂OBP基因在嗅觉以外的器官或组织中表达,说明OBPs可作为发育和其它生理过程的普通载体;而OBP1、OBP2、OBP4、OBP5、OBP6、OBP8、OBP11、OBP15和OBP12这7个基因在触角中高表达,在成蜂其它部位表达量低[3];而OBP1,即是PBP基因,在工蜂和小于2周的雄蜂触角中高表达[17]。
已研究的东方蜜蜂OBP基因序列一般长达400~ 500 bp,编码140~150个氨基酸。Ac-ASP1和Ac-ASP2的基因都在工蜂触角中高表达,Ac-ASP1属于PBP,其基因在幼虫和蛹期也表达且表达量丰富,而Ac-ASP2属于GOBP,其基因不在这两个发育阶段表达[9,10]。Ac-OBP3在蜜蜂触角和足的表达量显著高于在头和胸[14]。2010年张小辉等对Ac-OBP5、Ac-OBP9、Ac-OBP11和Ac-OBP12这几个基因进行克隆,分析表明它们的同源性低[12,13]。
昆虫OBPs在嗅觉中的功能机制备受研究者关注,其中对PBP功能机制的研究表明,昆虫中的PBP分为三类:长链PBP(约140个氨基酸长),中链PBP(约120个氨基酸长)和短链PBP(约100个氨基酸长)。刚开始,研究者发现长链PBP和短链PBP具有相似的功能机制,都如家蚕一样,在中性环境中PBP与配体结合,在酸性条件下PBP的构象发生变化,使之与配体解离[18]。但在飞蝗(Locusta migratoria)中研究表明:pH的改变对LmigOBP1的结构影响很小,说明昆虫间相似的PBP功能机制有其特异性[19]。不同种昆虫的GOBPs能作为载体运输各类气味分子,还能协同气味降解酶使气味分子失去活性。
西方蜜蜂PBP(OBP1)具有6个半胱氨酸,其中的4个半胱氨酸形成疏水性的信息素结合位点,能与主要蜂王信息素成分结合[17]。西方蜜蜂PBP与棉铃虫的PBP的功能机制相近,在酸性环境中与配体结合,在中性环境其构象改变后释放配体[20,21]。西方蜜蜂中的OBP2(ASP2)在中性条件中形成二聚体结构,能与多种气味结合;OBP13能与油脂和类似物质结合;OBP14与柠檬腈和丁香酚具有高亲和力,此外还能与报警信息素成分,即2-庚酮结合;OBP21也能与9-氧代-癸二烯酸结合,另外还与追踪信息素成分法尼醇具有较高的亲和性[22-24]。C-minus OBPs中OBP15-21缺少第二和第五个半胱氨酸,OBP14缺少第5个半胱氨酸,使得C-minus OBPs与OBP1(ASP1)即PBP在三级折叠结构有很大的区别,可使它们结合不同的挥发性物质[24]。
2.1.2化学感受蛋白
昆虫CSPs的主要功能是识别结合运输非挥发性分子,其分子量约为12~14 kDa,含100~120个氨基酸,具有4个保守半胱氨酸连接形成两个二硫键,三维结构大体上为球形,内含疏水结合区域。这类蛋白较为保守,含N-端的、中心区的和C-端区的3个保守区域。
1998年,Danty等在意大利蜜蜂触角中发现CSP基因,并将它们命名为ASP3,ASP3含3个亚型—ASP3a,ASP3b和ASP3c,经鉴定ASP3b与果蝇的CSP相似性高,属于CSP基因[25];后来ASP3c经Briand用实验证明也是CSP基因[26];在2006年Forêt在意大利蜜蜂中鉴定出6个CSP基因,并将它们命名为CSP1-6,其中的CSP1、CSP2即分别为ASP3b和ASP3c基因[3]。随着研究的进行,在中华蜜蜂中也陆续克隆表达出6 个Ac-CSPs基因。无论是意大利蜜蜂还是中华蜜蜂的CSPs都与其它昆虫的CSPs经典特征相似。
在昆虫中,CSP基因在触角、头、足、腹部、腺体、下颚须和下唇须等各个器官和组织都表达。CSP1,CSP2,CSP4在意大利蜜蜂和中华蜜蜂触角中都表达,Am-CSP4只在触角中表达;Ac-CSP3在蜜蜂翅中的表达量最高,而Am-CSP3在触角中表达丰富[27-30]。CSP5和CSP6都在意大利蜜蜂卵和卵巢中表达,但CSP6也在其幼虫期、蛹期和成蜂头中表达;而在中华蜜蜂中,CSP5在各个发育期都表达且表达量较为稳定,CSP6只在蛹期表达[29]。
昆虫CSPs具有多种功能,如嗅觉、调节发育、免疫、信号传导等。Maleszka等通过RNAi技术发现CSP5对意大利蜜蜂外胚层的发育有重要作用,CSP5的缺失会导致卵不能孵化,说明CSP5具有调节发育的功能[31]。Am-CSP2与芳香族化合物,说明其具有嗅觉功能。Ac-CSP3能与植物花和叶的挥发性次生代谢物结合,并且Am-CSP3能与16~18个碳的幼虫信息素结合,说明CSP3也具有一定的嗅觉功能[30]。
2.2嗅觉受体
昆虫的嗅觉受体(olfactory receptors,Ors)是含7个跨膜区域的跨膜蛋白,广泛存在于嗅觉感觉神经树突膜上;与脊椎动物G-蛋白偶联受体不同,它们是N端在细胞膜内,C端在细胞膜外。昆虫的Ors一般分为两类:一类是典型嗅觉受体(atypical odorant receptor,Ors),这些Ors基因高度变异同源性低;另一类其基因在不同昆虫间较为保守,同源性可高达70%以上,是传统嗅觉受体(conventional Ors即olfactory receptor coreceptor family,Orco;在果蝇中被命名为Or83b)。
Krieger等在2003年扩增出第一个西方蜜蜂Or基因AmelR2,这个基因在大多数触角嗅觉感觉神经中表达[32]。西方蜜蜂中有170个Ors基因,其中有7个假基因,并且大多数蜜蜂Ors基因是串联排列的,其中最大的串联阵列有60个Ors的序列,并且在2号染色体上;对其进行系统进化分析发现这些基因可分为5个亚家系,其中的4个亚家系都很小,第5个亚家系很大,包含157个Ors基因[33]。经序列分析表明AmOr2(即AmelR2)与DmOr83b的直系同源基因,属于Orco基因。Wanner等在2007年研究表明,Or10、Or11、Or18和Or170这4个基因在西方蜜蜂雄蜂中高表达,并且Or11对蜂王物质的主要成分9-氧代-癸二烯酸(9-ODA)有反应[34]。
中华蜜蜂中克隆表达定位了Orco、Or1、Or2、Or3 和Or11基因,这5个Ors基因都在触角中表达[35];Orco在内勤蜂触角表达量高,在采集蜂各个部位丰富表达;Or2在雄蜂触角中显著表达;Or11在采集蜂中成对地大量表达[11,36]。
昆虫ORs的功能是与气味分子结合后改变细胞膜上的离子通道产生动作电位。昆虫ORco能与OR结合成二聚体,功能之一是协助OR到达神经元树突膜上,例如在果蝇中敲掉OR83b,OR无法抵达神经元树突膜上[37];功能之二是可作为OR的分子伴侣能增强与OR的识别能力,如在家蚕BmOR2能使BmOR56对顺-茉莉酮的敏感度增高[38]。
2.3感觉神经元膜蛋白
感觉神经元膜蛋白(sensory neuron membrane proteins,SNMPs)是含有2个跨膜区域的膜结合蛋白,其基因属于人类脂肪酸转蛋白(FAT)CD36家族基因。最早在鳞翅目昆虫的嗅觉受体神经中发现SNMPs,研究表明这类蛋白可能在识别气味分子上具有重要作用,如果蝇的一个SNMP(即CG700)在信息素检测中具有重要作用,但其功能尚未完全研究清楚[39-41]。西方蜜蜂中有8个CD36同源的候选基因[42]。2013年胡颖颖等克隆表达出中华蜜蜂神经元膜蛋白AccSNMP1编码区,该基因1563 bp编码520个氨基酸,并且AccSNMP1在各个部位都表达,但在触角中表达最为丰富[43]。
2.4气味降解酶
气味降解酶(odorant degradation enzyme,ODEs)能使信号迅速失活,这对昆虫嗅觉具有重要作用,因为ODEs能避免持续信号刺激和保证后续嗅觉感知的传递。ODEs主要包含酯酶(esterases,EST)、细胞色素P450氧化酶(cytochrome P450s,P450)、谷胱甘肽-s-转移酶(glutathione S-transferases,GST)和醛氧化酶(aldehyde oxidase,AOX),并且这些酶中很多都具有解毒功能。
酯酶是胆碱酯酶、羧酸酯酶和磷酸酯酶等多种酯酶的统称,根据功能的特异性,羧酸酯酶又可分为α-酯酶、β-酯酶、乙酰胆碱酯酶、保幼激素酯酶、神经趋化蛋白、神经连接蛋白、gliotactins和glutactin共8个亚家族[44]。酯酶具备降解气味的功能,如雄性天蚕蛾触角中的酯酶ApolSE能降解信息素中乙酸酯成分;海灰翅夜蛾成虫触角中的羧酸酯酶SICXE10和甜菜夜蛾中的酯酶SexiCXE4能降解植物绿叶气味成分的(Z)-3-己烯醋酸盐[45-47]。细胞色素P450氧化酶参与信息素的代谢,如金龟子触角特异性P450能降解性外激素成分,此外,金龟子的CYP4AW1信息素、甘蓝夜蛾中的CYP4L4和CYP4S4能检测气味和信息素[48-50]。UGTs和AOX在昆虫触角中都有生成,也参与信息素的分解,并且UGTs能催化亲脂性糖基混合物与AOX能分解醛类化合物,有助于触角中有毒物质的代谢[51-53]。
2.5嗅觉系统机制
昆虫嗅觉系统机制有两种假说:其一是气味分子与感受蛋白结合后被运输到嗅觉神经元的树突上,因气味分子带有负电荷气味分子的积累,使得嗅觉神经元的树突周围的pH发生变化,从而导致气味分子与感受蛋白分离后与嗅觉受体结合产生动作电位[18,54];其二是气味分子与感受蛋白结合后,与感受神经元膜蛋白(SNMP)相互作用激发OR,OR与G-蛋白偶联后,由效应器激发第二信使,随着第二信使的增加激活离子通道[3];最后气味被气味降解酶降解。
3展望
嗅觉交流对蜜蜂具有重要作用,但目前尚未完全研究清楚,可以用RNA-Seq技术研究不同行为表现下蜜蜂触角几乎所有基因的表达情况;可以利用信息素成分或气味分子与OBP或者CSP是如何结合与释放的,以及它们与受体结合后是怎样产生电信号的,以及用信息素成分或气味分子刺激蜜蜂,分析嗅觉候选基因的表达是否受到影响;也可以利用western blot和免疫荧光等技术,对嗅觉相关基因进行定位,以及研究它们之间是否存在互作关系,逐步探明蜜蜂嗅觉机制与行为发育的关系。蜜蜂触角上分布着成千上万个感觉器,能感受环境中各种挥发性气味分子,赋予了蜜蜂灵敏的嗅觉,而且蜜蜂可以通过学习识别新的气味,具有可塑性。因此可深入研究蜜蜂触角,推进触角仿生的研究,发明识别特定气味灵敏的元器件,这样的元器件能用于军事和医学,如检测炸药、寻找尸体和诊断疾病等。
参考文献
[1]Fang Y,Song F,Zhang L,et al.Differential antennal proteome comparison of adult honeybee drone,worker and queen(Apis mellifera L.)[J].Journal of proteomics,2012,75(3):756-773.
[2]Feng M,Song F,Aleku D W,et al.Antennal proteome comparison of sexually mature drone and forager honeybees[J].Journal of proteome research,2011,10(7):3246-3260.
[3]Foret S.Function and evolution of putative odorant carriers in the honey bee(Apis mellifera)[J].2006.
[4]Vogt R,Lerner M.Two groups of odorant binding proteins in insects suggest specific and general olfactory pathways[J].Neurosci Abst,1989,15:1290.
[5]赵红霞,曾鑫年,梁勤,等.蜜蜂气味结合蛋白的生化特性、系统进化、表达部位和生理功能的研究[J].环境昆虫学报,2015,37 (2):420-425.
[6]Danty E,Arnold G,Huet J-C,et al.Separation,characterization and sexual heterogeneity of multiple putative odorant-binding proteins in the honeybee Apis mellifera L.(Hymenoptera:Apidea)[J]. Chemical senses,1998,23(1):83-91.
[7]Foret S M,R.Function and evolution of a gene family encoding odorant binding-like proteins in a social insect,the honey bee (Apis mellifera)[J].Genome research,2006,16(11):1404-1416.
[8]For T S,Maleszka R.Function and evolution of a gene family encoding odorant binding-like proteins in a social insect,the honey bee(Apis mellifera)[J].Genome research,2006,16(11):1404-1413.
[9]Li H,Nie W,Gao Q,et al.Cloning of cDNA encoding odorant binding protein ASP2 in working bee’s antenna of Apis cerana cerana and its prokaryotic expression[J].Scientia Agricultura Sinica, 2008,41(3):933-938.
[10]Li H-L,Gao Q-K,Cheng J-A.Cloning and spatio-temporal expression of cDNA encoding pheromone binding protein ASPl in Apis cerana cerana(Hymenoptera:Apidae)[J].Acta Entomologica Sinica,2008,51(7):689-693.
[11]张林雅.中华蜜蜂两种嗅觉相关蛋白的基因克隆与功能研究[D].中国计量学院,2013.
[12]张小辉,朱忠珂,祁艳霞.中华蜜蜂气味结合蛋白5,9,11, 12基因克隆及生物信息学分 [J].黑龙江畜牧兽医,2010,(4): 140-141.
[13]曹联飞,胡福良.蜜蜂属气味结合蛋白OBP5基因的进化[J].中国蜂业中旬刊(学术),2012,63(3):17-20.
[14]吉挺,沈芳,梁勤,等.中华蜜蜂OBP3基因的克隆,原核表达及组织表达谱[J].昆虫学报,2014,57(8):897-904.
[15]李红亮,聂文敏,高其康,等.中华蜜蜂气味结合蛋白ASP2 cDNA的克隆及原核表达 [J].中国农业科学,2008,41(3):933-938.
[16]李红亮,高其康,程家安.中华蜜蜂信息素结合蛋白ASP1 cDNA的克隆及时空表达[J].昆虫学报,2008,51(7):689-693.
[17]Danty E,Briand L C,Michard-Vanh E C,et al.Cloning and expression of a queen pheromone-binding protein in the honeybee: an olfactory-specific,developmentally regulated protein[J].The Journal of neuroscience,1999,19(17):7468-7475.
[18]Laughlin J D,Ha T S,Jones D N,et al.Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein[J].Cell,2008,133(7):1255-1265.
[19]Zheng J,Li J,Han L,et al.Crystal structure of the Locusta migratoria odorant binding protein[J].Biochemical and biophysical research communications,2015,456(3):737-742.
[20]Pesenti M E,Spinelli S,Bezirard V,et al.Queen bee pheromone binding protein pH-induced domain swapping favors pheromone release[J].Journal of molecular biology,2009,390(5): 981-990.
[21]Li Z-Q,Zhang S,Luo J-Y,et al.Two Minus-C odorant binding proteins from Helicoverpa armigera display higher ligand binding affinity at acidic pH than neutral pH[J].Journal of insect physiology,2013,59(3):263-272.
[22]Briand L,Nespoulous C,Huet J C,et al.Ligand binding and physico‐chemical properties of ASP2,a recombinant odorant‐binding protein from honeybee(Apis mellifera L.)[J].European Journal of Biochemistry,2001,268(3):752-760.
[23]Spinelli S,Lagarde A,Iovinella I,et al.Crystal structure of Apis mellifera OBP14,a C-minus odorant-binding protein,and its complexes with odorant molecules[J].Insect biochemistry and molecular biology,2012,42(1):41-50.
[24]Iovinella I,Dani F R,Niccolini A,et al.Differential expression of odorant-binding proteins in the mandibular glands of the honey bee according to caste and age[J].Journal of proteome research, 2011,10(8):3439-3449.
[25]E Danty G A,J-C Huetl,D Huet,et,al.Separation,Characterization and Sexual Heterogeneity of Multiple Putative Odorant-binding Proteins in the Honeybee Apis mellifera L(Hymenoptera:Apidea)[J].Chemical senses,1988,(23):83-91.
[26]Briand L,Swasdipan N,Nespoulous C,et al.Characterization of a chemosensory protein(ASP3c)from honeybee(Apis mellifera L.)as a brood pheromone carrier[J].European Journal of Biochemistry,2002,269(18):4586-4596.
[27]Liu Q,Wang H,Li H,et al.Impedance sensing and molecular modeling of an olfactory biosensor based on chemosensory proteins of honeybee[J].Biosensors and Bioelectronics,2013,40(1):174-179.
[28]李红亮,倪翠侠,姚瑞,等.中华蜜蜂化学感受蛋白基因Acer-CSP1克隆与表达特征分析[J].昆虫学报,2010,53(9):962-968.
[29]倪翠侠.中华蜜蜂化学感受蛋白基因家族克隆,分析及其功能鉴定[D].中国计量学院,2013.
[30]李红亮,张林雅,倪翠侠,等.中华蜜蜂化学感受蛋白AcerCSP3的配基结合功能分析 [J].昆虫学报,2011,54(3):259-264.
[31]Maleszka J,For T S,Saint R,et al.RNAi-induced phenotypessuggest a novel role for a chemosensory protein CSP5 in the development of embryonic integument in the honeybee(Apis mellifera) [J].Development genes and evolution,2007,217(3):189-196.
[32]Riddiford R Gvalm.Two groups of odorant binding proteins in insects suggest specific and general olfactory pathways[J].Neurosci Abst 1989,15:1290.
[33]Robertson H M,Wanner K W.The chemoreceptor superfamily in the honey bee,Apis mellifera:expansion of the odorant,but not gustatory,receptor family[J].Genome research,2006,16(11): 1395-403.
[34]Wanner K W,Nichols A S,Walden K K,et al.A honey bee odorant receptor for the queen substance 9-oxo-2-decenoic acid[J]. Proceedings of the National Academy of Sciences,2007,104(36): 14383-14388.
[35]赵慧婷.中华蜜蜂气味受体基因Or1,Or2,Or3的鉴定,表达及定位分析[D].山西农业大学,2013.
[36]张林雅,谢冰花,倪翠侠,等.中华蜜蜂Orco嗅觉受体基因的克隆,表达及亚细胞定位 [J].昆虫学报,2012,55(11):1246-1254.
[37]Smart R,Kiely A,Beale M,et al.Drosophila odorant receptors are novel seven transmembrane domain proteins that can signal independently of heterotrimeric G proteins[J].Insect biochemistry and molecular biology,2008,38(8):770-780.
[38]Tanaka K,Uda Y,Ono Y,et al.Highly selective tuning of a silkworm olfactory receptor to a key mulberry leaf volatile[J].Current Biology,2009,19(11):881-890.
[39]Benton R,Vannice K S,Vosshall L B.An essential role for a CD36-related receptor in pheromone detection in Drosophila[J]. Nature,2007,450(7167):289-293.
[40]Rogers M E,Sun M,Lerner M R,et al.Snmp-1,a novel membrane protein of olfactory neurons of the silk moth Antheraea polyphemus with homology to the CD36 family of membrane proteins[J].Journal of Biological Chemistry,1997,272(23):14792-14799.
[41]Rogers M E,Krieger J,Vogt R G.Antennal SNMPs(sensory neuron membrane proteins)of lepidoptera define a unique family of invertebrate CD36‐like proteins[J].Journal of neurobiology,2001, 49(1):47-61.
[42]Nichols Z,Vogt R G.The SNMP/CD36 gene family in Diptera, Hymenoptera and Coleoptera:Drosophila melanogaster,D.pseudoobscura,Anopheles gambiae,Aedes aegypti,Apis mellifera,and Tribolium castaneum[J].Insect biochemistry and molecular biology, 2008,38(4):398-415.
[43]Hu Y-Y,Xu S-F,Wubie A J,et al.Molecular characterization and tissue localization of sensory neuron membrane protein from Chinese honey bee,Apis cerana cerana(Hymenoptera:Apidae)[J]. Applied Entomology and Zoology,2013,48(4):533-545.
[44]Ranson H,Claudianos C,Ortelli F,et al.Evolution of supergene families associated with insecticide resistance[J].Science, 2002,298(5591):179-181.
[45]Vogt R G,Riddiford L M.Pheromone binding and inactivation by moth antennae[J].1981,293:161-163.
[46]Durand N,Carot-Sans G,Chertemps T,et al.Characterization of an antennal carboxylesterase from the pest moth Spodoptera littoralis degrading a host plant odorant[J].PloS one,2010,5(11): e15026.
[47]Merlin C,Rosell G,Carot Sans G,et al.Antennal esterase cDNAs from two pest moths,Spodoptera littoralis and Sesamia nonagrioides,potentially involved in odourant degradation[J].Insect molecular biology,2007,16(1):73-81.
[48]Wojtasek H,Leal W S.Degradation of an alkaloid pheromone from the pale-brown chafer,Phyllopertha diversa(Coleoptera: Scarabaeidae),by an insect olfactory cytochrome P450[J].FEBS letters,1999,458(3):333-336.
[49]Ma B Che Coisne M,Jacquin Joly E,Francois M,et al.cDNA cloning of biotransformation enzymes belonging to the cytochrome P450 family in the antennae of the noctuid moth Mamestra brassicae[J].Insect molecular biology,2002,11(3):273-281.
[50]Ma B Che-Coisne M,Nikonov A A,Ishida Y,et al.Pheromone anosmia in a scarab beetle induced by in vivo inhibition of a pheromone-degrading enzyme[J].Proceedings of the National A-cademy of Sciences of the United States of America,2004,101 (31):11459-11464.
[51]Rybczynski R,Reagan J,Lerner M R.A pheromone-degrading aldehyde oxidase in the antennae of the moth Manduca sexta[J]. The Journal of neuroscience,1989,9(4):1341-1353.
[52]Prestwich G D,Graham S M,Handley M,et al.Enzymatic processing of pheromones and pheromone analogs[J].Experientia, 1989,45(3):263-270.
[53]Ahn S-J,Vogel H,Heckel D G.Comparative analysis of the UDP-glycosyltransferase multigene family in insects[J].Insect biochemistry and molecular biology,2012,42(2):133-147.
[54]Mohanty S,Zubkov S,Gronenborn A M.The solution NMR structure of Antheraea polyphemus PBP provides new insight into pheromone recognition by pheromone-binding proteins[J].Journal of molecular biology,2004,337(2):443-451.
资助项目:国家自然科学基金(31340061)
作者简介:谢翠琴(1989-),女,硕士,研究方向为蜜蜂饲养与繁育。E-mail:3030068335@qq.com。
通讯作者:苏松坤,研究员。E-mail:susongkun@zju.edu.cn。
Advances in research on honey bee olfactory
Xie Cuiqin,Nie Hongyi,Su Songkun
(College of Bee Science,Fujian Agriculture and Forestry University,Fuzhou 350000)
Abstract:Olfactory system plays an important role in reproduction and development of honey bee,affecting mating,nursing,and foraging.The surface of the honey bee antennae,which is covered with numberous sensillum,has the ability to percept volatile molecules,odors,and hormones.Therefore,antennae are vital sensory organs of honey bees and accomplish an important role for olfactory communication.Therefore,we summarized honey bee antennae, olfactory sensillum,olfactory proteins and olfactory transmission mechanism,etc,which can provide reference for later researches as well as application of antennae in the honey bee.
Key words:honey bee;antennae;olfactory
