中心组合响应面优化里氏木霉B4菌产纤维素酶培养基组成研究*
2016-06-06屈海峰白殿国于占春王继艳宁艳春李爱力徐友海胡世洋
屈海峰,白殿国,于占春,岳 军,王继艳,宁艳春,李爱力,徐友海,胡世洋
(1.中国石油吉林石化公司 研究院,吉林 吉林 132021;2.吉林燃料乙醇有限责任公司,吉林 吉林 132101;3.钦州出入境检验检疫局,广西 钦州 535000)
化石燃料燃烧所排放的大量二氧化碳,造成了严重的温室效应,同时机动车尾气也是引起雾霾的原因之一。我国是一个农业大国,每年的农作物秸秆量约达7亿t,除少部分被利用外,大部分被焚烧,造成了严重的环境污染。而将秸秆转化为燃料乙醇,替代部分汽油使用,可以减少二氧化碳的排放,同时也可解决因秸秆焚烧所造成的环境问题[1-2]。
将木质纤维素材料转化为乙醇,首先需要将大分子状态的纤维素转化为酵母可以利用的可发酵性糖。主要途径有两条:一是通过酸法水解木质纤维素材料为可发酵性糖,另一条途径是生物转化法。由于酸法存在一系列的问题,生物转化法已成为主要的发展方向[3-4]。
里氏木霉是常用于发酵生产纤维素酶的菌株,能够分泌较多的胞外蛋白,酶系构成中外切酶组分占到蛋白含量的60%~70%,且比较均衡[5-6]。但是受制于较低的酶活性,如何提高里氏木霉的产酶能力仍是研究的重点。一方面可对菌株进行遗传改造[7-8],另一方面可通过发酵控制策略的研究,优化发酵条件,充分发挥菌株的性能[9-10]。碳源、氮源是影响里氏木霉产纤维素酶的两个主要因素,很多文献研究了碳、氮源种类对于酶活的影响,但多使用昂贵的纤维素粉、乳糖、酵母粉、玉米浆、豆饼粉,这些物质的使用增加了纤维素酶的生产成本[11-12],因此有必要研究使用廉价的碳、氮源替代,从而降低成本。
1 实验部分
1.1 材料、试剂与仪器
菌种为里氏木霉B4菌株:由本实验室保存,保存条件为4 ℃,马铃薯葡萄糖琼脂培养基(PDA培养基),保存过程中每月将菌株进行重复培养。
葡萄糖、KH2PO4、(NH4)2SO4、MgSO4、CaCO3、NaNO3,MnSO4·H2O、ZnSO4·7H2O、CoCl2·6H2O、FeSO4·7H2O:均为分析纯,玉米芯粉、麦麸:市售。
电热恒温水浴锅:HHS,上海博讯实业有限公司医疗设备厂;振荡培养箱:BS-1E,金坛市富华仪器有限公司;离心机:Anke TGL-16G,上海安亭科学仪器厂;微孔板光谱仪:Spectra Max Plus 384,美国 Molecular Device;超净工作台:苏州安泰空气技术有限公司。
种子培养基(1 L):20 g葡萄糖、3 g KH2PO4、5 g(NH4)2SO4、0.5 g MgSO4、3 g CaCO3、2 g NaNO3,上述物质加水配制成1 L溶液,pH值自然,500 mL的三角瓶中装入100 mL培养基,以上培养基均匀配制分装后121 ℃灭菌30 min。
产酶培养基(1 L):培养基中含有不同含量的玉米芯粉、麦麸、(NH4)2SO4,其余与种子培养基相同,500 mL的三角瓶中装入100 mL培养基,以上培养基均匀配制分装后121 ℃灭菌30 min。
微量元素浓缩液(1 L):250 mg MnSO4·H2O、360 mg ZnSO4·7H2O、370 mg CoCl2·6H2O、750 mg FeSO4·7H2O,上述物质加水配制成1 L溶液。
1.2 实验方法
1.2.1 种子制备
加入孢子悬液1 mL(孢子数量控制在106~108/mL)到灭菌后的种子培养基中,接种后置于恒温振荡培养箱中28 ℃,180 r/min下培养40 h,作为一级种子备用。
1.2.2 产酶培养
吸取10 mL种子液在无菌操作条件下,加入到产酶培养基中,放入恒温振荡培养箱中于28 ℃,180 r/min下培养96 h,培养过程中,每24 h取样并测定滤纸酶活。
1.2.3 实验设计
采用Minitab 15进行中心组合设计及统计学分析,利用Statistica 6.0软件进行响应面的分析。
1.2.4 滤纸酶活测定
滤纸酶活(FPase)的测定:将Waterman 1号滤纸裁剪成1 cm×6 cm大小的长条形状,质量为49.5~50.5 mg,折叠成M状放入25 mL比色管底部,加入 pH=4.8的醋酸-醋酸钠缓冲液1 mL,然后加入0.5 mL稀释适当倍数的粗酶液并混匀,对照为煮沸10 min灭活的相同稀释倍数的粗酶液。于50 ℃水浴水解60 min,然后用3,5-二硝基水杨酸(DNS)法测定生成的还原糖(以葡萄糖计)。FPase酶活力的定义是以上述条件下每分钟水解底物产生1 μmol还原糖所需的酶量为一个酶活力单位(IU)。
2 结果与讨论
2.1 三因素中心组合优化
碳源、氮源是影响里氏木霉产纤维素酶的主要因素[13-14],其中硫酸铵与豆饼粉是纤维素酶发酵过程中常用的两种氮源,两种氮源中硫酸铵为生理酸性盐,而豆饼粉被利用后,发酵液pH值呈碱性,里氏木霉的最适生长pH值在4~5,pH值过高会造成菌丝生长裂解,产酶量下降。相关研究表明在里氏木霉产纤维素酶的过程中,相比于有机氮源[15],无机氮源硫酸铵已经足够菌丝的生长及酶的分泌,并且采用廉价易得的麦麸及玉米芯代替价格较高的纤维素粉为碳源,可以降低成本。
实验过程中以麦麸、玉米芯、硫酸铵为主要因素,以滤纸酶活为响应值进行了三因素五水平的优化,并使用Statistic 6.0与Minitab 15软件进行响应面及统计学分析,因子及水平设计见表1,实验结果及预测结果对比见表2。

表1 三因素中心组合水平表
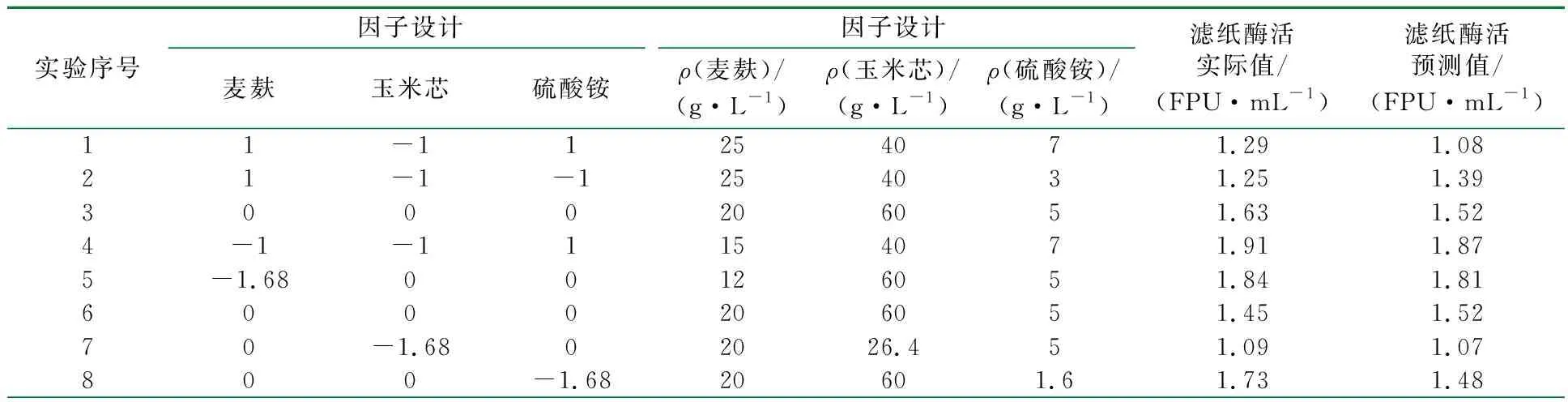
表2 中心组合因子水平设计及实验结果预测结果对比表
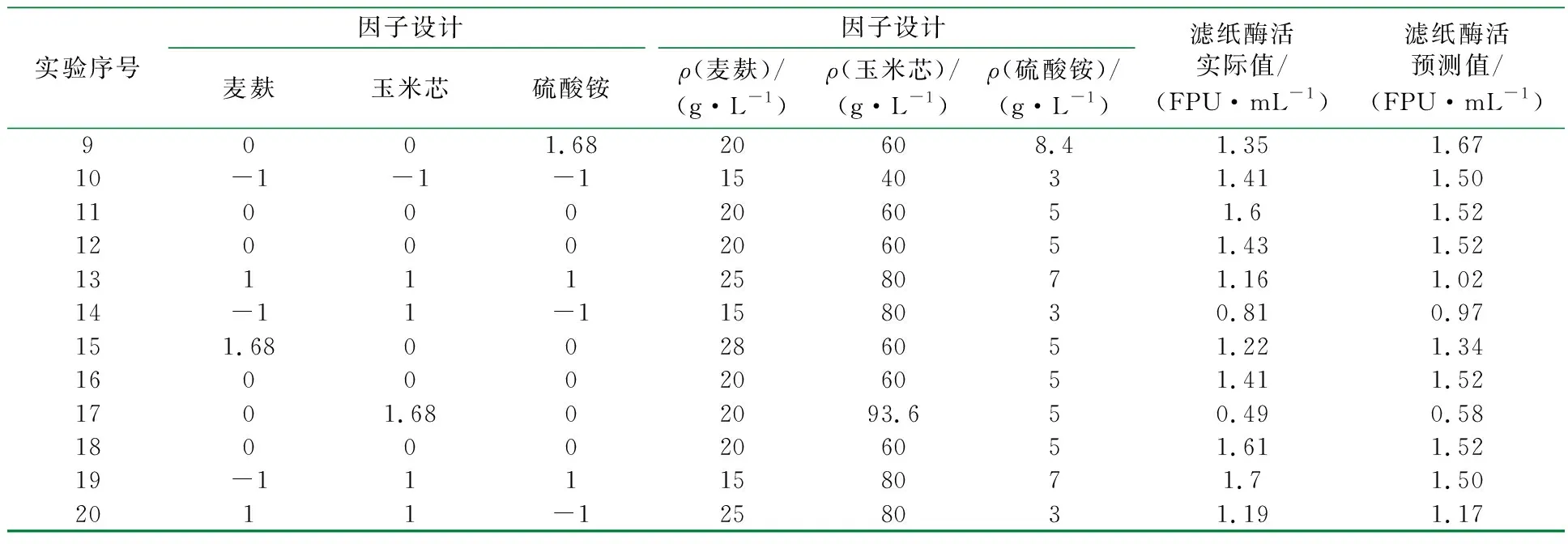
续表
响应面法可以直观看出因素对于响应值的影响,并且可以得出进一步的优化方向或是最佳工艺条件,为了对里氏木霉产纤维素酶的条件进行优化,利用Statistic 6.0软件分析了中心组合三个因素:ρ(麦麸)、ρ(玉米芯)和ρ(硫酸铵)对响应值滤纸酶活的影响,结果见图1~图3。通过对响应曲面和等高线图的分析可以看出ρ(麦麸)、ρ(玉米芯)和ρ(硫酸铵)这三个因素对于滤纸酶活的影响趋势。
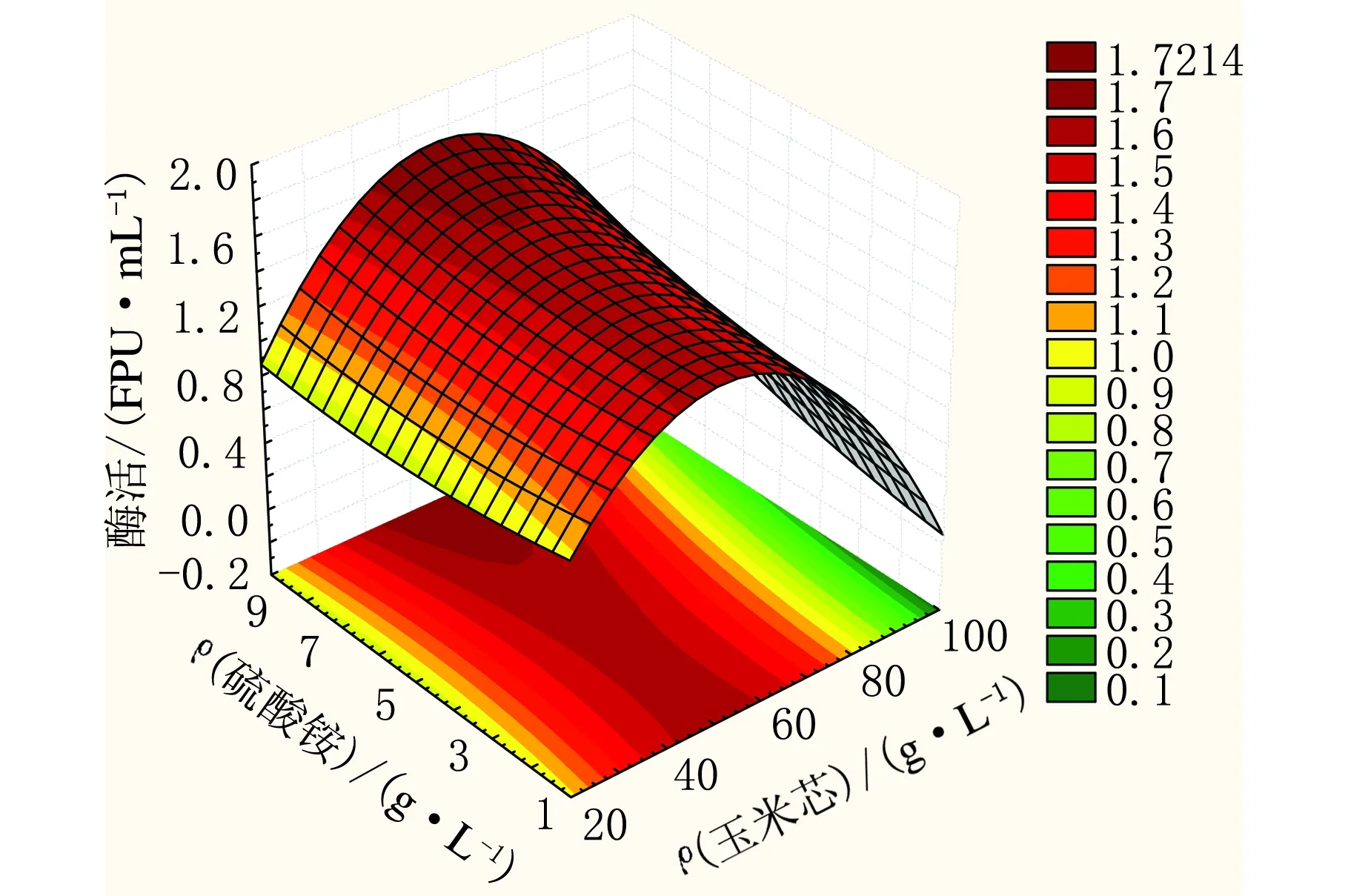
图1 中心组合响应面图(硫酸铵与玉米芯交互作用)
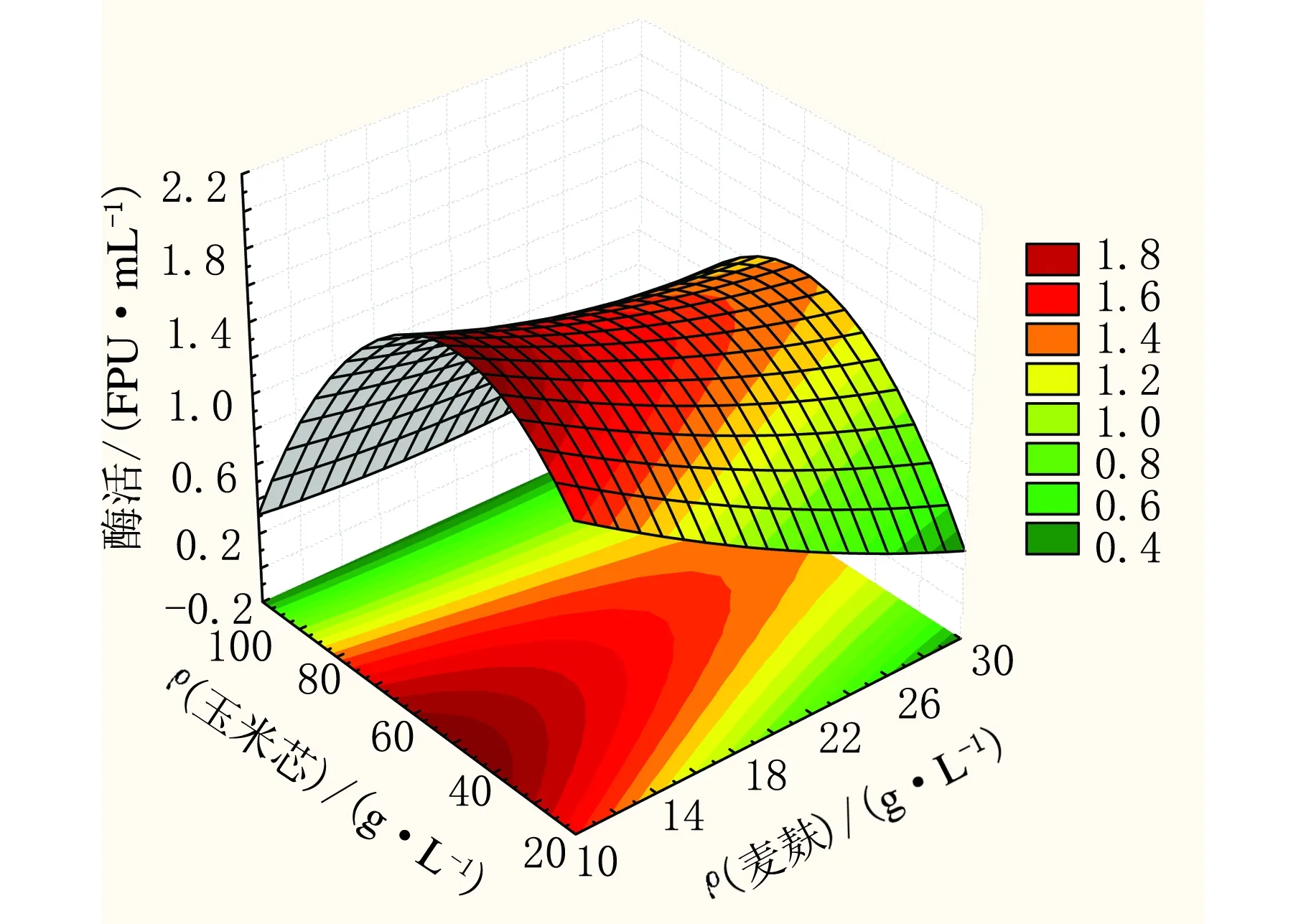
图2 中心组合响应面图(麦麸与玉米芯交互作用)
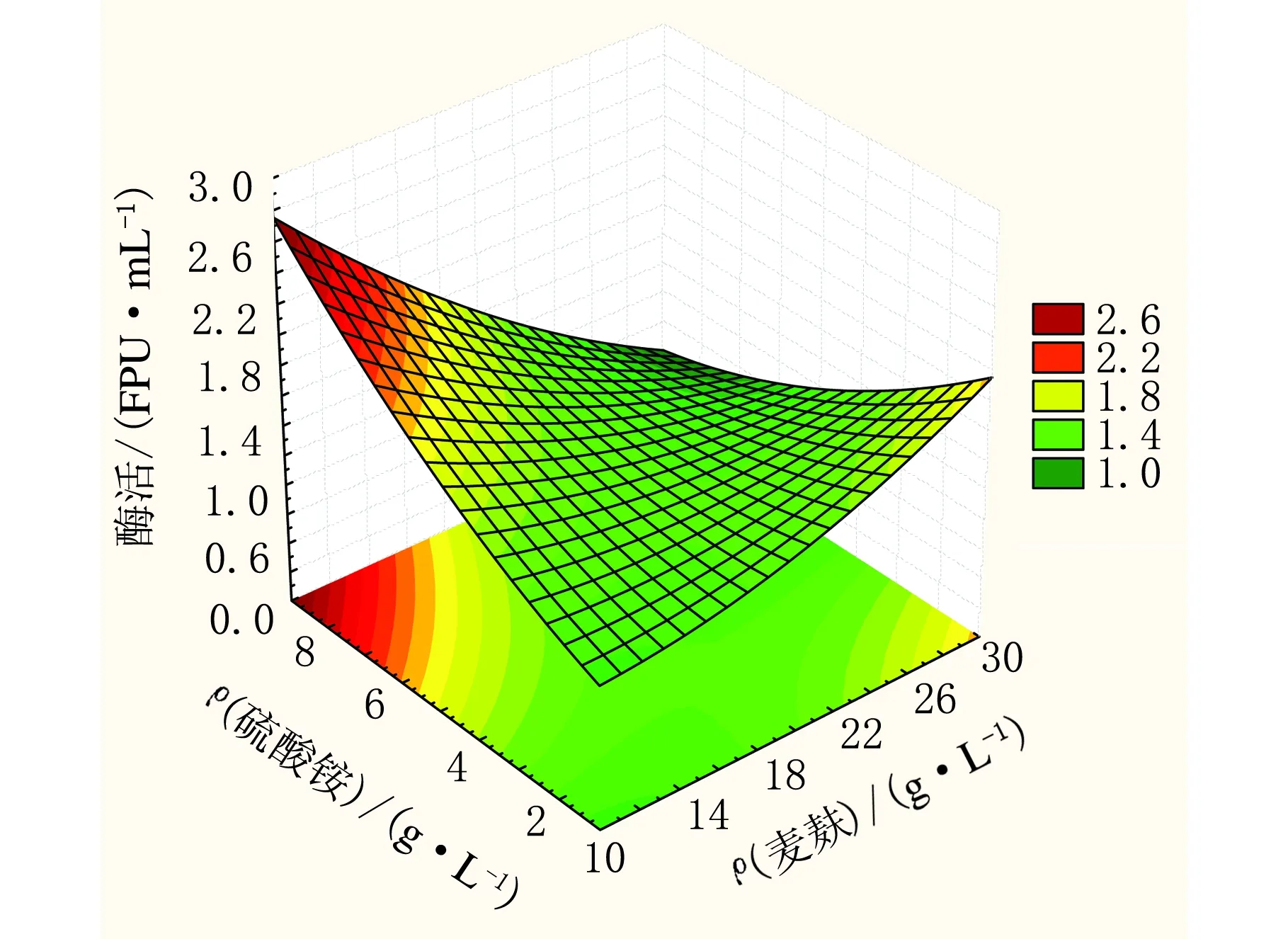
图3 中心组合响应面图(麦麸与硫酸铵交互作用)
由图1可知,当ρ(硫酸铵)由低水平增加到高水平时酶活有增加的趋势,固定硫酸铵用量,ρ(玉米芯)无论在低水平与高水平时都不利于产酶,较高的产酶区间是ρ(玉米芯)=40~60 g/L。
由图2可知,ρ(玉米芯)在高水平和低水平时,酶活都处在较低值,从三维曲面图中可以得出,当ρ(玉米芯)=50 g/L时,响应曲面的颜色最深,表示酶活处于最高值,因此为取得较高的酶产量,ρ(玉米芯)=50 g/L为宜;固定ρ(玉米芯),ρ(麦麸)由低水平增加到高水平时酶活逐渐降低,因此为提高酶的产量应减少麦麸的用量。
由图3可知,当ρ(麦麸)处于高水平而ρ(硫酸铵)处于低水平时酶活处于较高值,ρ(麦麸)处于高水平增加硫酸铵的用量会降低滤纸酶活;ρ(麦麸)与ρ(硫酸铵)同时处于低水平时酶活也处于较低值;当ρ(麦麸)处于低水平而ρ(硫酸铵)处于高水平时,酶活趋于增加的趋势。因此为增加滤纸酶活应增加硫酸铵的用量而相应的降低麦麸的用量。ρ(玉米芯)、ρ(硫酸铵)和ρ(麦麸)三个因素中,ρ(麦麸)对产酶的影响最小,应降低麦麸的用量。
利用Minitab软件对于ρ(玉米芯)、ρ(硫酸铵)和ρ(麦麸)三因素中心组合实验结果进行了统计学分析,结果见表3。
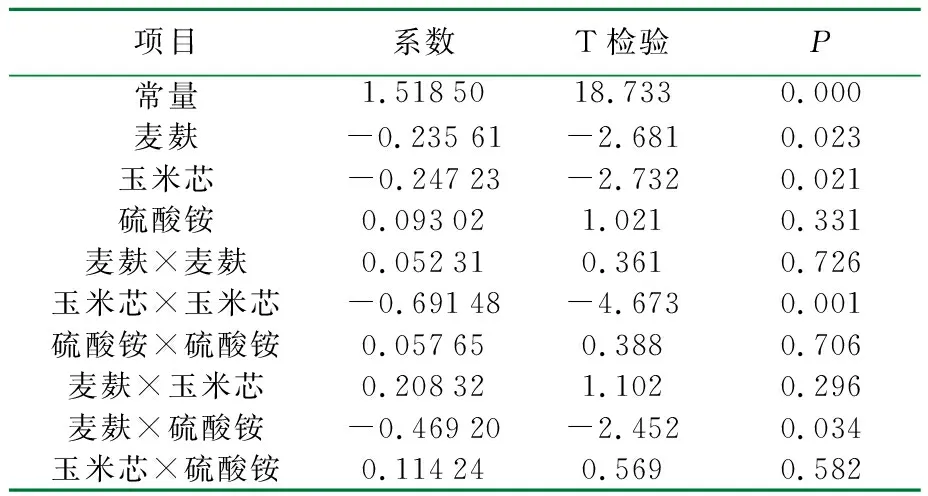
表3 中心组合优化以滤纸酶活为响应值的结果分析
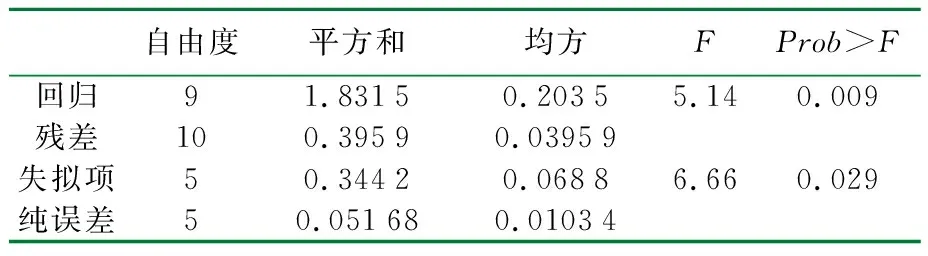
表4 以滤纸酶活为响应值的拟合模型统计学分析
2.2 两因素中心组合优化
在三因素的中心组合优化实验中,发现麦麸的加入量在低水平时对产酶更加有利,因此培养基中不添加麦麸,只对ρ(玉米芯)与ρ(硫酸铵)进行两因素五水平的全因子优化,利用Minitab软件进行实验设计,各水平实验值见表5。实验结果与模型的预测值见表6。

表5 二因素中心组合水平表
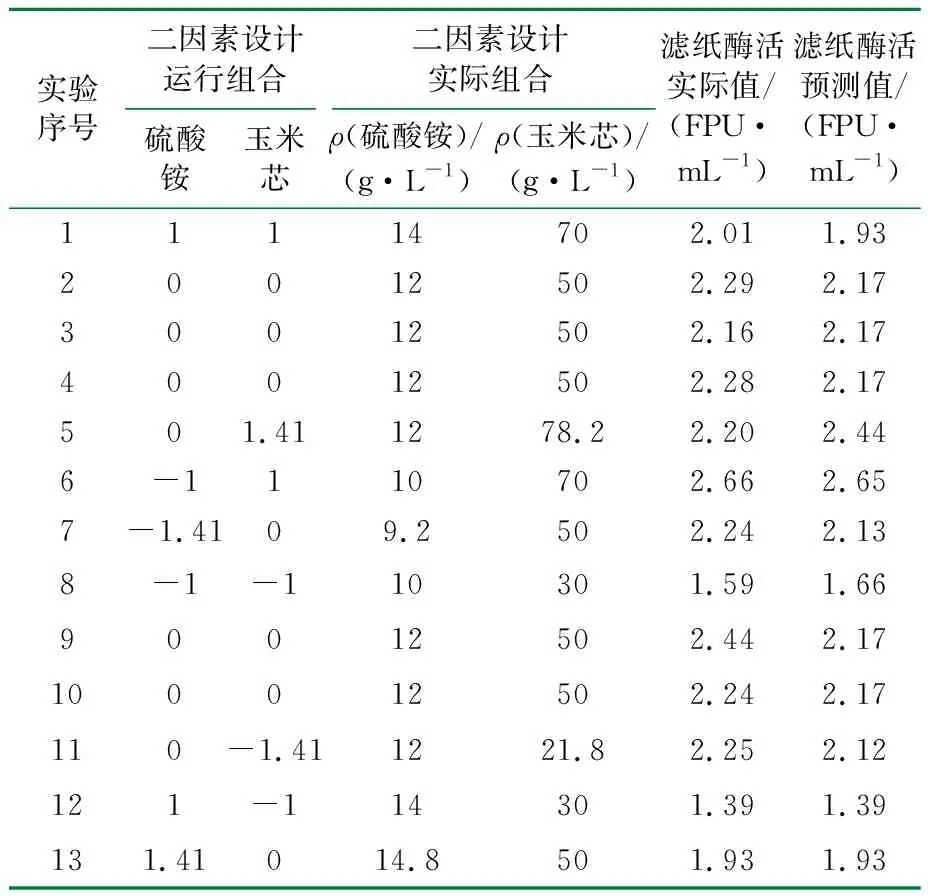
表6 二因素全因子水平设计及实验结果预测结果对比表
利用Statistic 6.0软件对ρ(玉米芯)、ρ(硫酸铵)以及滤纸酶活为响应值时进行了响应面分析(见图4)。通过对响应曲面和等高线图的分析可以看出,当ρ(玉米芯)<55 g/L时,无论硫酸铵在低水平与高水平时酶活都处于较低值,只有ρ(玉米芯)>55 g/L时,滤纸酶活才逐渐增加。较适宜的ρ(硫酸铵)=12 g/L,继续增加硫酸铵的用量导致酶活下降。
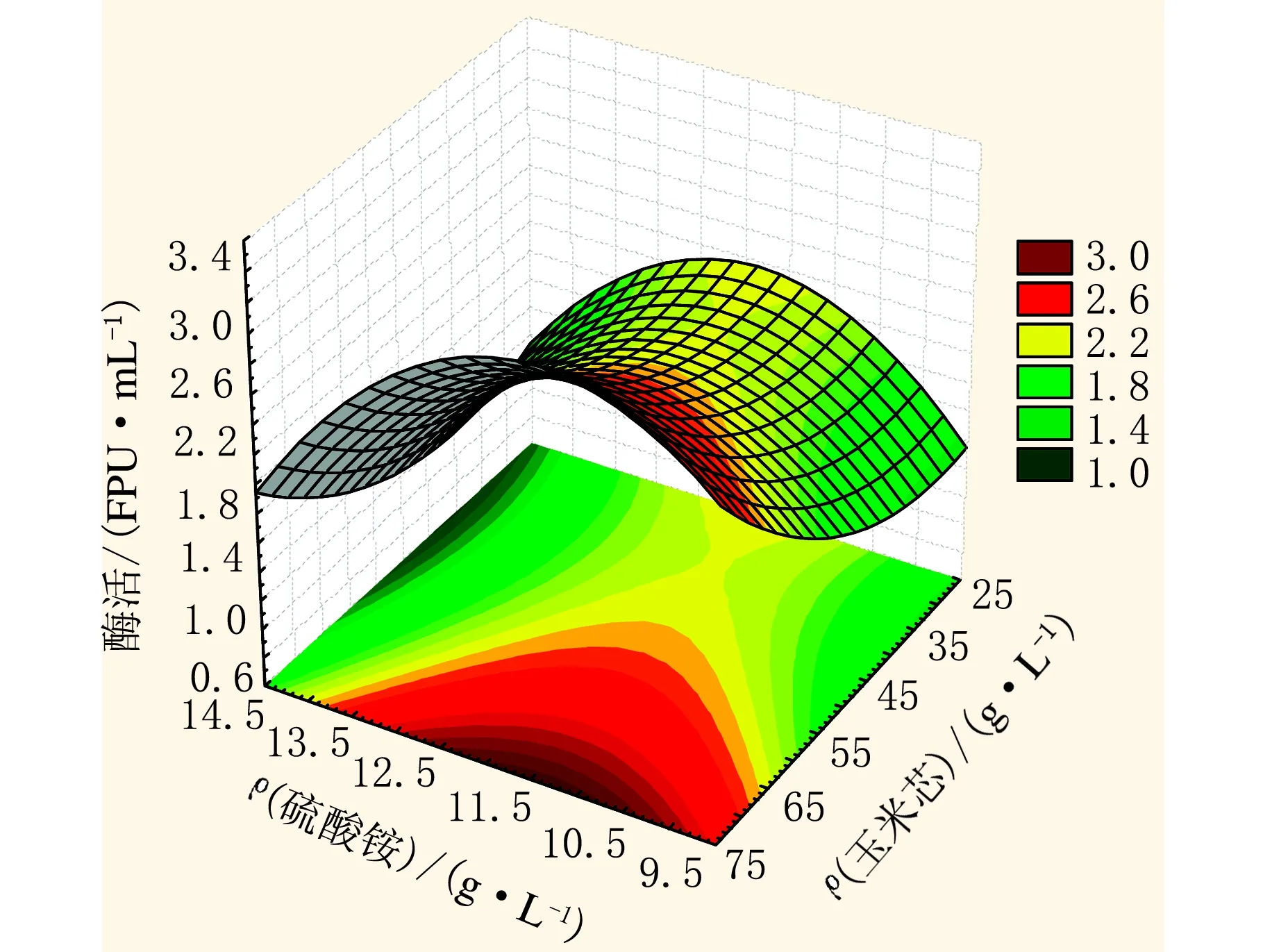
图4 二因素全因子响应面(玉米芯与硫酸铵交互作用)
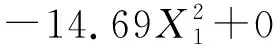
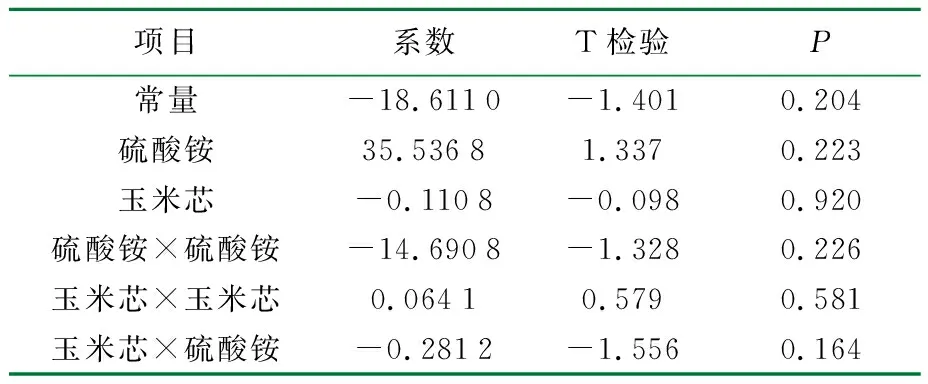
表7 中心组合优化以滤纸酶活为响应值的结果分析
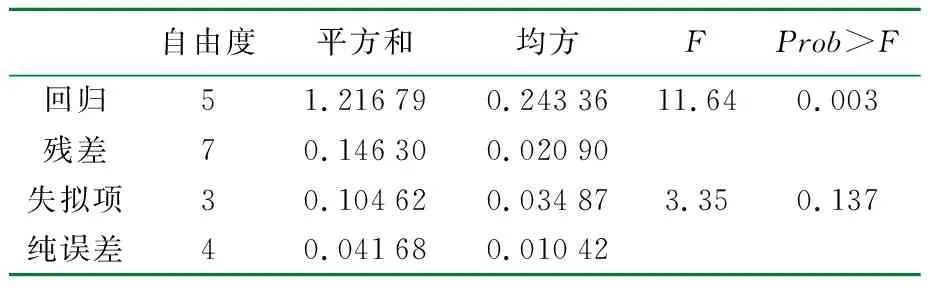
表8 以滤纸酶活为响应值的拟合模型统计学分析
3 结 论
采用廉价易得的碳源玉米芯、氮源硫酸铵,经过三因素五水平及两因素五水平两轮实验,里氏木霉产酶培养条件得到了优化。仅采用硫酸铵做为氮源也可以满足菌体的生长与产酶需求,模型预测的最优组成为ρ(玉米芯)=75 g/L、ρ(硫酸铵)=12 g/L,此条件下滤纸酶活为3.12 FPU/mL,较优化前提高58.38%,回归模型的p<0.05,模型显著,可以用于对实验结果的预测。
参 考 文 献:
[1] 赵建,曲音波.木质纤维素资源生物精炼技术研究进展[J].生命科学,2014(5):489-496.
[2] 方诩,秦玉琪,李雪芝,等.纤维素酶与木质纤维素生物降解转化的研究进展[J].生物工程学报,2010,26(7):864-869.
[3] 岳军,胡世洋,恵继星,等.木质纤维素材料预处理研究进展[J].现代化工,2014,34(10):31-35.
[4] 刘凯,戴莉,方诩,等.纤维素酶生产技术的研究进展[J].生物产业技术,2014(3):59-65.
[5] 王涫,王明钰,穆子铭,等.里氏木霉纤维素酶生产工艺的优化[J].黑龙江农业科学,2012(5):108-112.
[6] ROBYN PETERSON,HELENA NEVALAINEN.TrichodermareeseiRUT C30 thity years of strain improvement[J].Microbiology,2012,158:58-68.
[7] WANG S W,LIU G,WANG J,et al.Enhancing cellulse production inTrichodermareeseiRUT C30 through combined manipulation of activating genes[J].Jurnal of industrial microbiology biotechnology,2013,40:633-641.
[8] WU G H,HE R L,JIA W D,et al.Strain improvement and process optimization ofTrichdermareeseiRUT C30 for enhanced cellulase production[J].Biofuels,2012,2:545-555.
[9] LI C,YANG Z H,HE R L.Effect of pH on cellulase production and morphology ofTrichodermareeseiand the application in cellulosic material hydrolysis[J].Journal of Biotechnology,2013,168:470-477.
[10] MA L J,LI C,YANG Z H.Kinetic studies on batch cultivation ofTrichodermareeseiand application to enhance cellulase production by fed-batch fermentation[J].Journal of Biotechnology,2013,166:192-197.
[11] SARAVANAN P,MUTHUVELAYUDHAM R,RAJESH KANNAN R,et al.Optimization of cellulase production usingTrichodermareeseiby RSM and comparison with genetic algorithm[J].Front Chem Sci Eng,2012,6(4):443-452.
[12] SHAHRIARINOUR,M WAHAB MNA.,MOHAMAD R,et al.Effect of medium composition and cultural conditionon cellulase production byAspergillusterreus[J].African Journal of Biotechnology,2011,10(38):7459-7467.
[13] 伍红,陈秀珍,董志扬,等.碳源对里氏木霉QM9414胞外酶基因表达的影响[J].西南民族大学学报(自然科学版),2011,37(3):411-418.
[14] 勇强,李树炎,陈牧,等.氮源对里氏木霉木聚糖酶和纤维素酶生物合成的影响[J].林产化学与工业,2004,24(3):7-11.
[15] DIVANERY RODRIGUEZ,GOMEZ TIMOTHY,JOHN HOBLEY.Is an organic nitrogen source needed for cellulase production byTrechodermareeseiRUT C30?[J].World J Microbial Biotechnol,2013,29:2157-2165.
