背角无齿蚌对水体净化作用的研究
2016-06-05魏小飞关保华刘正文
魏小飞, 关保华, 刘正文,2
1. 华中农业大学水产学院, 武汉430070
2. 中国科学院南京地理与湖泊研究所, 湖泊科学与环境国家重点实验室, 南京 210008
背角无齿蚌对水体净化作用的研究
魏小飞1, 关保华2,*, 刘正文1,2
1. 华中农业大学水产学院, 武汉430070
2. 中国科学院南京地理与湖泊研究所, 湖泊科学与环境国家重点实验室, 南京 210008
通过设置不同生物量的背角无齿蚌, 研究其对再悬浮水体总悬浮质(TSS)、叶绿素 a(Chla)及水体营养盐含量的影响。结果表明: (1)背角无齿蚌可以快速降低水体中总悬浮质(TSS)和叶绿素a(Chla)的浓度; (2)增大背角无齿蚌生物量可加快水体总悬浮质(TSS)和叶绿素 a(Chla)浓度的下降速度; (3)背角无齿蚌在第 2天可降低水体总氮(TN)、总磷(TP)、总溶解磷(TDP)的含量, 升高总溶解氮(TDN)的含量, 但试验结束时, 空白组和有蚌组总氮(TN)、总溶解氮(TDN)、总磷(TP)、总溶解磷(TDP)均没有显著差异(P>0.05)。因此, 背角无齿蚌可在水体生态修复初期用于快速降低水体悬浮物和浮游藻类含量, 但从长期看, 不能明显改善水体营养盐含量。
滤食性底栖动物; 总悬浮质; 叶绿素a; 营养盐; 透明度
1 前言
湖泊富营养化是当今世界面临的重大环境问题。据统计, 目前全球约有 75%以上的封闭型水体存在富营养化问题[1], 我国绝大多数湖泊也已经富营养化或者正在富营养化中[2]。随着水体富营养化进程的加快, 沉水植物的衰退和消失变成了一个普遍现象[3-4], 进而导致水体内浮游藻类和沉积物再悬浮的增加[5], 水体透明度降低, 最终破坏整个生态系统。沉水植物不仅能够快速吸收水体和沉积物中的营养盐, 分泌产生他感物质抑制浮游植物生长[7],而且在密度较高的情况下, 可以改变湖水流向和流强, 影响湖水与底泥之间物质交换平衡[8], 因此, 沉水植物恢复是浅水富营养化湖泊修复的关键[6]。但沉水植物的良好生长受多方面因素影响, 其中高悬浮物导致的低透明度是沉水植物恢复的主要限制因子[9]。因此在生态修复初期, 我们需要迅速提高水体的透明度, 降低水体中悬浮质的含量, 从而为沉水植物的生长提供有利的条件。国内外已有研究表明,滤食性枝角类可以控制浮游植物, 提高透明度, 促进沉水植物恢复[10], 而淡水双壳类软体动物作为滤食性生物, 也能滤食水中的浮游藻类[11]和有机碎屑[12],提高水体透明度。
背角无齿蚌(Anodonta woodiana) 是我国淡水水体中分布很广的淡水双壳类软体动物之一[13], 是典型的滤食性动物。研究表明, 背角无齿蚌在滤水过程中能形成大量的“假粪”, 产生显著的生物沉降作用, 可加速水体中悬浮颗粒物向底层转移, 有助于提高水体透明度[14-15]。但与此同时, 贝类的排泄作用不断的向水体内排放氨、尿素、尿酸、氨基酸、可溶性磷等营养物质, 且氨氮占总排泄量的60%—70%[16], 从而可能提高水体溶解性营养盐的浓度。因此, 能否利用背角无齿蚌修复富营养化水体生态系统有着十分显著的意义。本研究以苏州太湖东山附近本地的土著背角无齿蚌为研究对象, 比较其不同生物量对水体的净化能力的大小, 以求为今后的富营养化水体生态修复提供科学依据。
2 材料与方法
2.1 试验材料
背角无齿蚌采自太湖, 选取大小一致(50.0 ± 5.0 g)、健康的个体备用。试验用水为河蟹养殖池塘水。底泥采自太湖, 用 40目筛网除去螺类、石子等杂物,混匀后备用。试验装置为150 L的蓝色塑料桶, 规格是底径45 cm、口径60 cm、高75 cm。搅动泵功率为3 W, 流量200 L·h-1。
2.2 试验设计
试验于2013年9月20日至9月30日在中科院南京地理与湖泊研究所位于苏州市东山镇的野外试验站进行。试验共设3个处理, 每个处理设置3个平行(见表1)。每个试验桶铺设约10 cm厚底泥, 加水130 L, 3 W搅动水泵固定于距离底泥10 cm处, 24小时连续工作并保证搅动方向一致。待水桶内生态系统相对稳定, 向各处理组分别放入对应生物量的背角无齿蚌, 河蚌用尼龙网挂于距水面20 cm处, 尼龙网底部置塑料平板, 减少尼龙网对河蚌活动的影响。
试验期间, 每天观察桶内河蚌和水泵搅动情况,确保水泵的正常运行和河蚌的准确位置, 保持桶内水位稳定。
2.3 试验指标及测定方法
试验过程中对所有试验桶水体总悬浮质(TSS)、叶绿素a(Chla)、总氮(TN)、总溶解氮(TDN)、总磷(TP)、总溶解磷(TDP)浓度等指标进行测定, 待搅动水泵工作稳定后, 采集水样作为本底值, 之后每两天采样一次, 用柱状采水器采集1 L水样, 用于理化指标的测定。同时YSI测定实时水体温度、pH等指标。采样后, 补充相同体积的蟹塘水。
相关指标及测定方法: TSS采用重量法; Chla采用丙酮萃取分光光度法; TN和TDN采用碱性过硫酸钾消解紫外分光光度法; TP和TDP采用碱性过硫酸钾消解钼酸铵分光光度法; 具体分析方法参考金相灿和屠清瑛(1990)[17]。
2.4 数据处理及统计分析
所有试验数据均由Microsoft Excel 2010处理,由Origin Pro 8.5生成图形; 所有试验数据的描述性统计及检验统计均采用 SPSS19.0进行, Repeated Measures对重复测量数据进行分析, 多元方差分析使用 Multivariate分析方法, 其中显著性水平为 P<0.05, 极显著性水平位P<0.01。
3 结果与分析

表1 不同处理组背角无齿蚌设置情况Tab. 1 Initial value of Anodonta woodiana in different treatments
3.1 不同处理组水体总悬浮质浓度的变化
水体TSS浓度变化见图1。在整个实验期间, 所有处理组TSS浓度呈逐步下降趋势, 低蚌组TSS浓度低于空白组, 但差异不显著(P>0.05)。高蚌组TSS浓度显著低于空白组 (P<0.05)。有蚌处理组TSS浓度在试验开始 2 d后出现显著的下降, 空白组 TSS浓度与低蚌组存在显著差异(P<0.05), 低蚌组与高蚌组存在显著差异(P<0.05), 空白组与高蚌组存在极显著差异(P<0.01)。试验结束时, 各处理组间TSS浓度差异不显著, 但高蚌组浓度在试验期间一直低于空白组和低蚌组。
3.2 不同处理组水体叶绿素a浓度的变化
水体Chla浓度变化见图2, 空白组呈现先上升后下降的趋势但始终高于其他处理组, 低蚌组变化不大, 高蚌组整体呈现下降趋势, 其中空白组与高蚌组存在显著差异(P<0.05)。试验开始后第2天空白组 Chla浓度与低蚌组存在显著差异(P<0.05), 与高蚌组存在极显著差异(P<0.01), 低蚌组与高蚌组差异不显著(P>0.05)。试验结束时, 各处理组 Chla浓度差异不显著, 但高蚌组<低蚌组<空白组。
3.3 不同处理组氮浓度的变化
各处理组TN浓度变化见图3A。可以看出, 各个处理组 TN浓度均呈现先降低后升高的趋势, 但各个组别间差异不显著(P>0.05)。高蚌组试验第 2天 TN浓度与第 1天相比存在极显著差异(P<0.01),第7天TN浓度与第2天相比存在显著差异(P<0.05)。试验结束时, 低蚌组和高蚌组 TN浓度均略高于空白组, 但差异不显著(P>0.05)。
各处理组TDN浓度变化见图3B。与TN变化趋势不同的是, 各个处理组TDN浓度均呈现逐渐升高的趋势, 但不同处理组之间差异不显著(P>0.05)。第 2天, 空白组 TDN浓度与高蚌组存在显著差异(P<0.05)。试验结束时, 各处理组 TDN浓度关系为空白组<低蚌组<高蚌组, 但差异不显著(P>0.05)。
3.4 不同处理组磷浓度的变化
不同处理组TP浓度变化见图4A。各个组TP浓度均呈现不同程度的下降趋势, 其中空白组TP浓度显著高于高蚌组(P<0.05)。多元方差分析表明, 第2天空白组TP浓度与高蚌组存在显著差异(P<0.05),低蚌组TP浓度也显著高于高蚌组(P<0.05), 但空白组与低蚌组差异不显著(P>0.05)。试验结束时各处理组TP浓度由高到低的顺序为: 空白组>低蚌组>高蚌组,但差异不显著(P>0.05)。

图1 不同处理组总悬浮质浓度变化Fig. 1 Comparison of total suspended substance in different treatments

图2 不同处理组叶绿素a浓度变化Fig. 2 Comparison of Chla in different treatments

图3 不同处理组总氮及总溶解氮浓度变化Fig. 3 Comparison of TN and TDN in different treatments
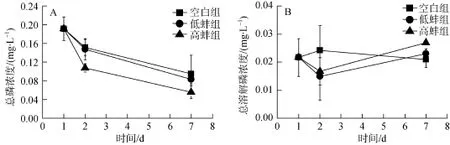
图4 不同处理组总磷及总溶解磷浓度变化Fig. 4 Comparison of TP and TDP in different treatments
不同处理组TDP浓度变化见图4B。其中空白组呈现先升高后降低的趋势, 而低蚌组与高蚌组则呈现先降低后升高的趋势, 但各个组别之间差异不显著(P>0.05)。多元方差分析表明, 第 2天空白组TDP浓度与高蚌组存在显著差异(P<0.05), 其他组别之间差异不显著。试验结束时, 各处理组TDP浓度由低到高的顺序为: 空白组<低蚌组<高蚌组, 但差异不显著(P>0.05)。
4 讨论
贝类可以通过滤食悬浮颗粒提高水体透明度,从而改善生态系统的光照条件[18]。研究表明, 背角无齿蚌能控制浮游藻类的生长[19]。控制悬浮物或浮游植物能力与贝类滤水率有关, 而不同体重的背角无齿蚌的滤水速率有明显区别, 特别是体重小于100 g 的蚌的滤水速率显著增加[13]。本研究显示, 背角无齿蚌的滤食可以快速降低水体TSS和Chla的浓度, 尤其是在悬浮物和浮游藻类较多时, 其滤食作用更为明显; 且加大背角无齿蚌生物量可加快水体TSS和Chla浓度的下降速率。
本试验初期, 水体内悬浮物和浮游藻类浓度较高, 在背角无齿蚌的滤食作用下, 水体悬浮物和浮游藻类快速下降, 总氮、总磷含量也下降; 但随着水体悬浮物和浮游藻类的减少, 水体中总溶解氮和溶解性总磷的含量逐渐上升, 可能与背角无齿蚌本身的营养盐排泄作用有关[22-23], 在试验结束时有蚌组和无蚌组在总磷含量上并没有显著差异。而目前我们普遍采用氮、磷、叶绿素等指标来评定水体的富营养化程度[24]。因此, 背角无齿蚌的存在并不能显著降低营养盐浓度。因此, 贝类的排泄也为浮游藻类的生长提供营养盐, 但当淡水双壳类的滤食强度增加到一定程度并超过藻类的生长时, 就可以在较高的营养水平下抑制浮游藻类的生长, 从而保持较高的水体透明度[21]。因此, 我们可以利用背角无齿蚌的滤食作用来去除水体内的悬浮颗粒物和浮游藻类, 从而提高水体的透明度, 可为沉水植物的生长提供良好的条件, 促进沉水植物的恢复, 从而恢复以沉水植物为主的清水态浅水湖泊生态系统。
5 结论
综上所述, 背角无齿蚌可快速降低水体内悬浮颗粒物和浮游藻类的含量, 提高水体透明度, 且在一定范围内, 增大背角无齿蚌生物量可加快悬浮颗粒物和浮游藻类的下降速度, 但并不能长期明显改善水体内营养盐含量, 因此单纯依靠背角无齿蚌进行富营养化水体的修复工作是不可行的。
在富营养化水体生态修复中, 沉水植物是降低水体内营养盐含量的主要因素, 而沉水植物生长的重要先决条件就是提高水体的透明度, 从而使得沉水植物能够获得充足的光照进行生长。因此, 在富营养化水体生态修复中, 可以利用背角无齿蚌的滤食作用首先降低水体悬浮颗粒物和浮游藻类的含量,提高水体透明度, 为后期沉水植物的生长提供良好的条件, 在整个生态修复过程中存在重要意义。
[1] FREEDMAN B. Environmental Ecology[M]. Sandiego:Academic Press, 2002.
[2] 成小英, 李世杰. 长江中下游典型湖泊富营养化演变过程及其特征分析[J]. 科学通报, 2006, 51:848-855.
[3] JIN Xiangcan, CHU Jianzhou, WANG Shengrui. Effects of Nitrogen Concentration and Form in Water on Photosynthetic Characteristics of Hydrilla verticillata and Myriophyllum verticillatum[J]. Chinese Journal of Applied and Environmental Biology, 2007, 13(2): 200-204.
[4] MELZER A. Aquatic macrophytes as tools for lakemanagement[J]. Hydrobiologia, 1999, 395/396: 181-190.
[5] MOSS B. Engineering and biological approaches to the restoration for eutrophication of shallow lakes in which aquatic plant communities are important components[J]. Hydrobiologia, 1990, 200/201: 367-377.
[6] 王军霞, 张亚娟, 刘存歧, 等. 水生植物在富营养化湖泊生物修复中的作用[J]. 安徽农业科学, 2011, 39(10):6055-6057, 6110.
[7] GOI A. A study of toxicity, biodegradability, and some by-products of ozonized nitrophenols[J]. Advances in Environmental Research, 2004, 8(3): 303-311.
[8] 吴振斌, 邱东茹, 贺锋, 等. 沉水植物重建对富营养水体氮磷营养水平的影响[J]. 应用生态学报, 2003, 14(8):1351-1355.
[9] 王华, 冯勇, 刘申宝, 等. 沉水植物生长影响因子研究进展[J]. 生态学报, 2008, 28(8): 3958-3968.
[10] ERIK J, MARTIN S, TORBEN L L, et al. Chapter 6 -Biomanipulation as a Restoration Tool to Combat Eutrophication: Recent Advances and Future Challenges[M]. Advances in Ecological Research, 2012:411-488.
[11] REEDERS H H, DE VAATE B A. Zebra mussels (Dreissena polymorpha): a new perspective for water quality management[J]. Hydrobiologia, 1990, 200/201: 437-450.
[12] SHPIGEL M, Blaylock R A. The pacific oyster as abiological filter for a marine fish[J]. Aquaculture, 1991, 92(2-3): 187-197.
[13] 吴庆龙, 陈宇炜, 刘正文. 背角无齿蚌对浮游藻类的滤食选择性与滤水率研究[J]. 应用生态学报, 2005, 16(12):2423-2427.
[14] 胡元亮. 实用药用动物养殖技术[M]. 北京: 中国农业出版社, 2001.
[15] 吴桂汉, 陈品健, 江瑞胜. 滤食性贝类在对虾综合养殖生态系统中的作用[J]. 台湾海峡, 2001, 20(4) : 447-452.
[16] 周毅, 杨红生, 张福绥. 四十里湾栉孔扇贝的生长余力和C 、N 、P 元素收支[J]. 中国水产科学, 2002, 9(2):161-166.
[17] 金相灿, 屠清瑛. 湖泊富营养化调查规范[M]. 北京: 中国环境科学出版社, 1990.
[18] ZHU B, FITZGERALD D G, MAYER C M, et al. Alteration of Ecosystem Function by Zebra Mussels in Oneida Lake: Impacts on Submerged Macrophytes[J]. Ecosystems, 2006, 9: 1017-1028.
[19] 杨东妹, 陈宇炜, 刘正文, 等. 背角无齿蚌滤食对营养盐和浮游藻类结构影响的模拟[J]. 湖泊科学, 2008, 20(2):228-234.
[20] 葛长字. 体重与氨氮浓度对河蚌氨氮排泄的影响[J]. 安徽农业科学, 2010, 38(4): 1881-1883, 1887.
[21] 周毅, 杨红生, 何义朝, 等. 四十里湾几种双壳贝类及污损动物的氮、磷排泄及其生态效应[J]. 海洋与湖沼, 2002, 33(4): 424-431.
[22] 秦伯强, 高光, 朱广伟, 等. 湖泊富营养化极其生态系统响应[J]. 科学通报, 2013, 58(10): 855-864.
[23] CARPENTER S R, KITCHELL J F, HODGSON J R. Cascading trophic interactions and lake productivity[J]. Bioscience, 1985, 35: 634-639.
Research on water purification of Anodonta woodiana
WEI Xiaofei1, GUAN Baohua2,*, LIU Zhengwen1,2
1. College of Fisheries, Hua Zhong Agricultural University, Wuhan 430070, China
2. State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China
Our study aims to reveal the influence of Anodonta woodiana on the total suspended substance (TSS), chlorophyll a (Chla) and water nutrients via setting different levels of Anodonta woodiana biomass. The results are as follows. (1) Anodonta woodiana could lower the concentrations of TSS and Chla in the water quickly. (2) The concentrations of TSS and Chla decreased faster in the treatments of high biomass of Anodonta woodiana. (3) Anodonta woodiana decreased the concentrations of TN, TP, TDP and boosted the TDN in the second day, while there was no significant difference (P >0.05) between the blank treatments and the Anodonta woodiana treatments at the end of this experiment. This study indicates that Anodonta woodiana can reduce the TSS concentrations and phytoplankton biomass obviously it the beginning of ecological restoration process, but it does not improve the water qualities in a long term view.
filter-feeding zoobenthos; total suspended substance; chlorophyll a; nutrients; transparency
10.14108/j.cnki.1008-8873.2016.01.008
A
1008-8873(2016)01-056-05
2015-03-17;
2015-05-12
国家自然科学基金(31270409);国家科技重大专项(2013ZX07101014-2)
魏小飞(1989—), 男, 河北保定人, 硕士研究生, 研究方向为浅水湖泊生态修复, E-mail: xiaof_wei@163.com
*通信作者:关保华, 女, 博士, 副研究员, 主要从事水生植物生态学、湖泊生态修复科学与技术的研究, E-mail: bhguan@niglas.ac.cn
魏小飞, 关保华, 刘正文. 背角无齿蚌对水体净化作用的研究[J]. 生态科学, 2016, 35(1): 56-60.
WEI Xiaofei, GUAN Baohua, LIU Zhengwen. Research on water purification of Anodonta woodiana[J]. Ecological Science, 2016, 35(1): 56-60.
