超级稻品种中嘉早17产量相关性状QTL定位研究
2016-04-29圣忠华朱子亮马宁李威贺记外魏祥进邵高能王建龙胡培松唐绍清
圣忠华 朱子亮 马宁 李威 贺记外 魏祥进 邵高能 王建龙 胡培松 唐绍清,*
(1中国水稻研究所, 杭州 310006; 2湖南农业大学 农学院, 长沙 410128; #共同第一作者; *通讯联系人,E-mail: sqtang@126.com)
超级稻品种中嘉早17产量相关性状QTL定位研究
圣忠华1,#朱子亮2, #马宁1李威2贺记外1魏祥进1邵高能1王建龙2胡培松1唐绍清1,*
(1中国水稻研究所, 杭州 310006;2湖南农业大学 农学院, 长沙 410128;#共同第一作者;*通讯联系人,E-mail: sqtang@126.com)
圣忠华, 朱子亮, 马宁, 等. 超级稻品种中嘉早17产量相关性状QTL定位研究. 中国水稻科学, 2016, 30(1): 35-43.
摘要:以中嘉早17/D50 F2和F(2∶3)群体为材料,考查各群体株系产量相关农艺性状,同时构建其遗传连锁图谱,最后定位到与产量性状相关的66个QTL,其贡献率变幅为0.08% ~ 20.45%,分布于除第9染色体外的其他各染色体上。两个群体在第3、7、10染色体某区段同时定位到多个与产量性状相关的QTL, 这些QTL的加性效应来自中嘉早17。在定位到的66个QTL中,F2和F(2∶3)群体均检测到,且贡献率较大的QTL qPH-10(株高)、qFLW-4(剑叶宽)、qTGW-2(千粒重) 加性效应均来自中嘉早17。初步检测到超级稻品种中嘉早17携带多个高产QTL,为今后将中嘉早17的优良基因导入其他水稻品种进而培育产量更高、综合性状更优良的超级稻新品种提供理论和技术支持。
关键词:QTL 定位; 产量性状; 超级稻
水稻是世界上主要粮食作物,一半以上世界人口以大米为主食。近年来,全球每年对稻米的需求以1.2%幅度增长,然而,全球水稻单产每年增幅却不足1%[1-2]。在我国,随着城镇化的不断推进,全国水稻种植面积正逐年递减,因而提高水稻单产是解决我国粮食安全问题的最有效最重要的途径。提高水稻单产的措施主要有两方面:1)栽培措施改良提高水稻单产;2)培育高产水稻品种。通过栽培措施改良一般需要投入较大的物力财力,培育高产水稻新品种则相对经济高效[3-4]。水稻产量性状主要包括有效穗数、每穗粒数、结实率、千粒重等[5-6]。由于水稻产量性状大多属于数量性状,故目前对其研究应用较多的是QTL分析法:分析其产量相关性状的QTL位点,之后构建次级分离群体以及近等基因系精细定位产量相关性状基因,最后通过图位克隆的方法克隆目的基因[7-8]。
截至目前,已被精细定位甚至克隆的水稻产量相关性状基因包括株高、生育期、粒形、千粒重等相 关基因。其中Ghd7是一个能同时控制水稻每穗粒数、株高和抽穗期的主效QTL,利用珍汕97和明恢63构建的F2∶3和重组自交系群体将Ghd7最初定位于水稻第7染色体上标记R1440和C1023 之间;进一步,Ghd7被精细定位到RM5436和RM2256之间的79 kb区间内。Ghd7的CDS全长1013 bp,编码一个由257个氨基酸组成的核蛋白,该产物是一个CCT(CO, CO-LIKE 和 TIMING OF CAB1)结构蛋白。长日照条件下,Ghd7的增强表达能使抽穗期延迟并增加株高和每穗粒数,而功能减弱的自然变异品系能够种植到温带甚至更冷的地区。因此,Ghd7在增加全球水稻产量潜力和提高适应性方面有着非常重要的作用[9]。GS3是控制水稻粒重和粒长的主效QTL,被定位于水稻第3染色体着丝粒区域,与标记GS09和MRG5881连锁。同时,GS3也是控制水稻粒宽和籽粒充实度的微效QTL,在调节籽粒和器官大小中发挥负调节子的功能[10-11]。另外,影响稻谷粒宽和粒重的GW2,其等位基因显著增加谷粒粒宽和粒重。GW2的CDS包含有8个外显子,全长1634 bp,编码425个氨基酸的蛋白产物。由于WY3第4外显子上一个碱基的缺失,引起GW2等位基因在转录过程中提前终止[12]。水稻产量性状受多基因共同调节,其代谢调控网络极其复杂,对控制水稻产量性状基因的挖掘有利于丰富和完善其代谢调控网络,进而从分子水平阐明水稻高产机理。
中嘉早17是中国水稻研究所培育的超级稻新品种,2008年通过浙江省品种审定,2009年通过国家品种审定,2012年通过湖北省品种审定;平均单产7500 kg/hm2左右。中嘉早17是目前我国年推广面积最大的籼稻品种。为了挖掘和分析中嘉早17高产相关基因,阐明其高产机理,本研究以中嘉早17与热带粳稻品系D50为亲本,构建中嘉早17/D50 F2及F2∶3群体;并于2013年早季和2014年早季分别考查各群体株系的产量相关性状,构建F2群体的遗传连锁图谱,分析中嘉早17高产相关性状QTL,以期阐明中嘉早17高产机理。
1材料与方法
1.1供试材料
2012年将超级稻品种中嘉早17与热带粳稻品种D50杂交,2012年冬季在海南陵水种植中嘉早17/D50 F1。 2013年早季种植中嘉早17/D50 F2群体及亲本。2013年秋季随机收取中嘉早17/D50 F2群体及亲本单株300个,2014年早季种植中嘉早17/D50 F2∶3群体共225个株系。
1.2材料种植与农艺性状考查
中嘉早17、D50及中嘉早17/D50 F2群体于2013年3月26日播种,4月27日移栽,株行距为16.5 cm×22.1 cm,单本栽插。中嘉早17、D50及中嘉早17/D50 F2∶3群体于2014年3月28日播种,4月30日移栽,每个株系种植6行,每行8株,株行距为16.5 cm×22.1 cm,单本栽插。田间水肥管理及病虫害防治同常规大田。
成熟时,亲本中嘉早17、D50及中嘉早17/D50 F2,F2∶3群体取中间单株考查农艺性状,其中亲本中嘉早17与D50各选取整齐一致的5个单株考查农艺性状。 F2群体在中间选取300个单株考查各单株农艺性状,在F2群体300个单株的基础上,衍生出300个F2∶3株系,即F2∶3群体;F2∶3群体各株系选中间8个单株考查各农艺性状。按照考种标准对各单株的株高、剑叶长、剑叶宽、有效穗数、穗长、单株总粒数、结实率及千粒重共8个农艺性状进行考查,利用SPSS 19.0进行数据统计分析。
1.3SSR分子标记连锁图谱构建及QTL定位分析
利用本实验室556对公共SSR引物对中嘉早17和D50两亲本进行多态性筛选[13];所有的SSR引物由上海英骏生物技术有限公司合成。DNA提取方法采用SDS法[14]。
将筛选出的在亲本中嘉早17与D50之间有多态性的引物标记用于中嘉早17与D50 F2群体各单株的基因型分型,利用Mapmaker 3.0进行连锁图谱的构建,标记分组(group)取LOD≥2.5,采用最大似然值算法进行作图,利用Kosambi函数将重组率转化为图距[15]。利用构建好的遗传连锁图谱,结合表型农艺性状统计值,采用Win QTL cartographer 2.5软件进行QTL分析[16];采用McCouch 的命名系统对定位到的QTL进行命名[17]。
2结果与分析
2.1F2、F2∶3群体及其亲本与产量相关农艺性状的表型值分析
2013年中嘉早17/D50 F2群体及其亲本、2014年中嘉早17/D50 F2∶3群体及其亲本的8个与产量性状相关的农艺性状的表型统计参数及分布见表1与图1。与双亲的各性状表型值比较分析结果表明:中嘉早17/D50 F2群体与中嘉早17/D50 F2∶3群体的各种性状分离明显,具有双亲的各种重组类型;而且大部分性状表现出双向超亲分离现象,并呈连续分布状态。峰值和偏值在F2群体中表现超过1的性状为有效穗数、单株总粒数、结实率。峰值和偏值在F2∶3群体中表现超过1的性状有剑叶宽与结实率。在这8个与产量相关的性状表型值中,除了株高、剑叶长、穗长、单株总粒数的高值表现为亲本D50之外;其他性状的高值表现为亲本中嘉早17。图1结果分析表明,除单株总粒数之外其他性状无论在F2群体还是F2∶3群体均呈单峰分布,表明这些性状呈正态分布,受数量性状遗传控制;另外除有效穗数、穗长、单株总粒数及千粒重之外其他性状在F2与F2∶3群体两世代表现出较好的重复性,表明这些性状受环境影响较小在世代间能稳定遗传。
2.2F2、F2∶3群体不同产量相关性状间的相关系数分析
对中嘉早17/D50 F2和F2∶3群体产量性状间相关分析表明,部分产量相关性状间存在显著甚至极显著的相关性(表2)。其中,株高与剑叶长、株高与剑叶宽、剑叶长与剑叶宽、有效穗数与穗长、有效穗数与单株总粒数、单株总粒数与穗长、单株总粒数与结实率等性状间在F2和F2∶3群体均检测到显著甚至极显著的相关性,且方向一致。其他产量相关性状间在F2群体或F2∶3群体也检测到显著或极显著的相关性。如:F2群体中穗长与株高(r=0.268)、剑叶长(r=0.153)存在极显著相关;有效穗数与千粒重(r= - 0.136)显著相关。在F2∶3群体,结实率与千粒重(r=0.118)显著相关。表明水稻产量是一个复杂的农艺性状,构成产量的各因素间关系密切,共同影响水稻产量的形成。
表1中嘉早17/D50 F2与F2∶3群体及亲本的农艺性状表型
Table 1. Agronomic traits performances of F2and F2∶3populations of Zhongjiazao 17 × D50.

性状与世代TraitandgenerationF2/F2∶3分离群体Segregationpopulations变幅VariationX±SD峰值Peak偏值Partial亲本ParentD50中嘉早17Zhongjiazao17株高PH/cm F267.13~123.0092.52±0.660.180.1590.5077.71 F2∶371.76~130.70101.92±0.710.37-0.2691.2079.30剑叶长LFL/cm F220.14~53.8934.04±0.340.830.3031.4030.80 F2∶319.10~44.5431.76±0.32-0.100.1633.1030.30剑叶宽WFL/cm F21.06~2.231.65±0.010.200.011.311.79 F2∶31.18~3.771.66±0.022.263.521.351.81有效穗数NTPP F25.50~36.0014.12±0.351.981.0513.6015.10 F2∶34.38~12.007.62±0.10-0.130.5212.8015.60穗长PL/cm F218.56~33.2525.54±0.170.52-0.1726.8620.74 F2∶317.21~26.4422.00±0.110.10-0.3027.1019.80单株总粒数NFGP F2897.00~4295.001654.35±33.245.281.692188.431586.63 F2∶3594.88~1747.881076.50±15.30-0.010.521785.301521.36结实率SSR F20.09~0.880.57±0.011.24-0.5662.2971.30 F2∶30.20~0.870.63±0.011.14-0.6465.3078.10千粒重TGW/g F215.17~29.0022.97±0.150.440.0622.0025.60 F2∶318.25~33.2527.09±0.170.23-0.1722.5026.20
PH, Plant height; LFL, Length of flag leaf; WFL, Width of flag leaf; NTPP, Number of panicles per plant; PL, Panicle length; NFGP, Number of filled grains per plant; SSR, Seed-setting rate; TGW, 1000-grain weight. The same as in tables below.

YK17-中嘉早17。
YK17, Zhongjiazao 17.
图1中嘉早17/D50 F2与F2∶3群体农艺性状表型频率分布
Fig. 1. Phenotypic frequency distributions of agronomic traits in F2and F2∶3populations of Zhongjiazao 17 × D50.
2.3F2、F2∶3群体与产量相关性状的QTL分析
利用Win QTL cartographer 2.5软件进行QTL区间作图分析(表3和图2),结果表明在这8个与产量相关的性状中都检测到QTL的存在,共有66个QTL,其中,检测到QTL最多的性状是剑叶长和穗长,都检测到12个QTL位点;检测到QTL数最少的性状是单株总粒数和千粒重,都检测到5个QTL。在所有检测到的QTL中,F2和F2∶3群体均检测到的QTL数为13个,其中株高6个:qPH-1、qPH-3-2、qPH-5、qPH-6、qPH-7、qPH-10;剑叶长2个:qFLL-2和qFLL-7-2;剑叶宽1个:qFLW-4;穗长1个:qPL-3-3;千粒重3个:qTGW-2、qTGW-3和qTGW-6。在F2和F2∶3群体均检测到的QTL中,除部分株高QTL的加性效应来自D50之外,其他均来自中嘉早17。另外,有效穗数和千粒重所定位到的QTL的加性效应基本都来自中嘉早17。
所有检测到的QTL贡献率变幅为0.08% ~ 20.45%,其中,在F2和F2∶3群体均能检测到且贡献率均大于9%的QTL有qPH-10(株高)、qFLW-4(剑叶宽)、qTGW-2(千粒重)。所有检测到的QTL除第9染色体外,在其他染色体上均有分布;其中,在第1染色体上分布最多,为11个;在第8染色体上分布最少,为1个;另外,第3、7、10染色体某些区域内同时定位到多个与产量相关的QTLs,如在第3染色体SSR标记RM1373与RM7389之间同时定位到控制株高、剑叶长、穗长的QTL;在第7染色体标记RM118与RM248之间也同时定位到控制株高、剑叶长、穗长的QTL;在第10染色体标记RM1375与RM25664之间同时检测到控制株高、剑叶长、剑叶宽、穗长和有效穗数的QTL。
表2中嘉早17/D50 F2与F2∶3群体不同性状间的相关系数
Table 2. Correlation coefficients among agronomic traits in F2and F2∶3populations of Zhongjiazao 17 × D50.

性状Trait(F2)株高PH剑叶长LFL剑叶宽WFL有效穗数NTPP穗长PL单株总粒数NFGP结实率SSR千粒重TGW性状Trait(F2∶3)株高PH0.322**0.242**0.0040.066-0.009-0.0080.080株高PH剑叶长LFL0.323**0.211**-0.0090.0490.003-0.035-0.038剑叶长LFL剑叶宽WFL0.407**0.271**-0.0660.0700.023-0.119*0.036剑叶宽WFL有效穗数NTPP-0.099-0.014-0.107-0.184**0.671**-0.0020.111有效穗数NTPP穗长PL0.268**0.153**0.087-0.158**0.205**-0.151**-0.073穗长PL单株总粒数NFGP-0.0100.003-0.152**0.312**0.123*-0.119*-0.013单株总粒数NFGP结实率SSR0.0140.026-0.022-0.213**0.050-0.260**0.118*结实率SSR千粒重TGW0.084-0.0770.081-0.136*-0.008-0.055-0.070千粒重TGW
*,**分别表示P<0.05、P<0.01显著水平。
*and**mean significant levels atP<0.05 andP<0.01,respectively.
表3中嘉早17/D50 F2与F2∶3群体检测到的产量相关性状QTL分布情况
Table 3. QTL distributions of yield related traits detected in F2and F2∶3populations of Zhongjiazao 17 × D50.
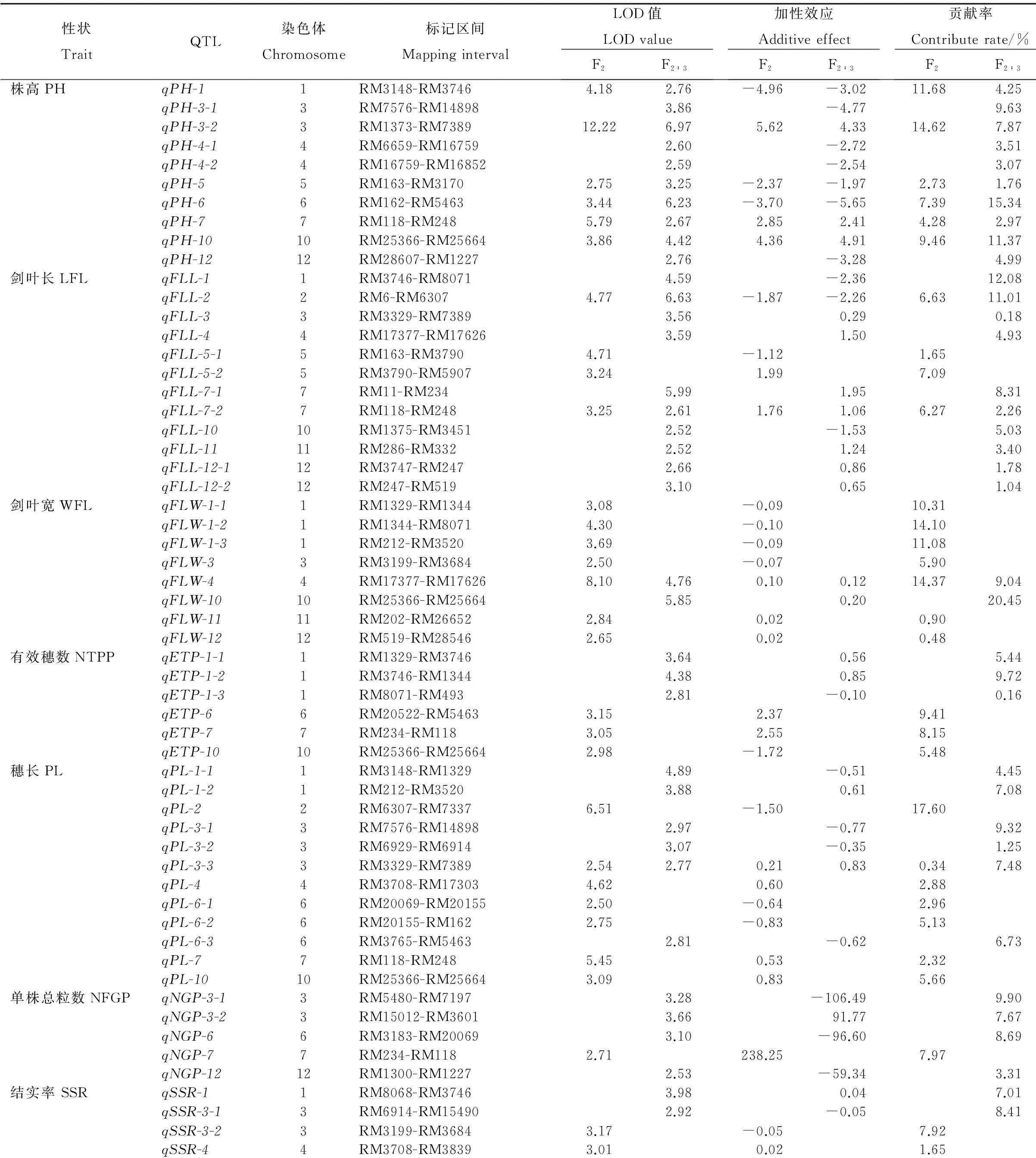
性状TraitQTL染色体Chromosome标记区间MappingintervalLOD值LODvalueF2F2∶3加性效应AdditiveeffectF2F2∶3贡献率Contributerate/%F2F2∶3株高PHqPH-11RM3148-RM37464.182.76-4.96-3.0211.684.25qPH-3-13RM7576-RM148983.86-4.779.63qPH-3-23RM1373-RM738912.226.975.624.3314.627.87qPH-4-14RM6659-RM167592.60-2.723.51qPH-4-24RM16759-RM168522.59-2.543.07qPH-55RM163-RM31702.753.25-2.37-1.972.731.76qPH-66RM162-RM54633.446.23-3.70-5.657.3915.34qPH-77RM118-RM2485.792.672.852.414.282.97qPH-1010RM25366-RM256643.864.424.364.919.4611.37qPH-1212RM28607-RM12272.76-3.284.99剑叶长LFLqFLL-11RM3746-RM80714.59-2.3612.08qFLL-22RM6-RM63074.776.63-1.87-2.266.6311.01qFLL-33RM3329-RM73893.560.290.18qFLL-44RM17377-RM176263.591.504.93qFLL-5-15RM163-RM37904.71-1.121.65qFLL-5-25RM3790-RM59073.241.997.09qFLL-7-17RM11-RM2345.991.958.31qFLL-7-27RM118-RM2483.252.611.761.066.272.26qFLL-1010RM1375-RM34512.52-1.535.03qFLL-1111RM286-RM3322.521.243.40qFLL-12-112RM3747-RM2472.660.861.78qFLL-12-212RM247-RM5193.100.651.04剑叶宽WFLqFLW-1-11RM1329-RM13443.08-0.0910.31qFLW-1-21RM1344-RM80714.30-0.1014.10qFLW-1-31RM212-RM35203.69-0.0911.08qFLW-33RM3199-RM36842.50-0.075.90qFLW-44RM17377-RM176268.104.760.100.1214.379.04qFLW-1010RM25366-RM256645.850.2020.45qFLW-1111RM202-RM266522.840.020.90qFLW-1212RM519-RM285462.650.020.48有效穗数NTPPqETP-1-11RM1329-RM37463.640.565.44qETP-1-21RM3746-RM13444.380.859.72qETP-1-31RM8071-RM4932.81-0.100.16qETP-66RM20522-RM54633.152.379.41qETP-77RM234-RM1183.052.558.15qETP-1010RM25366-RM256642.98-1.725.48穗长PLqPL-1-11RM3148-RM13294.89-0.514.45qPL-1-21RM212-RM35203.880.617.08qPL-22RM6307-RM73376.51-1.5017.60qPL-3-13RM7576-RM148982.97-0.779.32qPL-3-23RM6929-RM69143.07-0.351.25qPL-3-33RM3329-RM73892.542.770.210.830.347.48qPL-44RM3708-RM173034.620.602.88qPL-6-16RM20069-RM201552.50-0.642.96qPL-6-26RM20155-RM1622.75-0.835.13qPL-6-36RM3765-RM54632.81-0.626.73qPL-77RM118-RM2485.450.532.32qPL-1010RM25366-RM256643.090.835.66单株总粒数NFGPqNGP-3-13RM5480-RM71973.28-106.499.90qNGP-3-23RM15012-RM36013.6691.777.67qNGP-66RM3183-RM200693.10-96.608.69qNGP-77RM234-RM1182.71238.257.97qNGP-1212RM1300-RM12272.53-59.343.31结实率SSRqSSR-11RM8068-RM37463.980.047.01qSSR-3-13RM6914-RM154902.92-0.058.41qSSR-3-23RM3199-RM36843.17-0.057.92qSSR-44RM3708-RM38393.010.021.65

图2中嘉早17/D50 F2与F2∶3群体均检测到的产量相关性状QTL及其所包含的已被精细定位和克隆的产量相关基因在染色体上的分布情况
Fig. 2. QTLs distribution of yield related traits on chromosomes detected both in F2and F2∶3populations of Zhongjiazao 17× D50, and some fine mapped or cloned yield related genes in the mapped QTL intervals.
中嘉早17/D50 F2及F2∶3群体检测到的与产量相关QTL具体分析如下:
1)株高。本研究在中嘉早17/D50 F2及F2∶3群体共检测到控制株高QTL有10个,分布于第1、3、4、5、6、7、10、12染色体上,其对表型的贡献率从1.76%至15.34%不等。在中嘉早17/D50 F2及F2∶3群体中均能检测到的株高QTL有6个,分别位于第1、3、5、6、7、10染色体上,加性效应来自亲本中嘉早17的有3个,来自亲本D50的有3个。
2)剑叶长。在中嘉早17/D50 F2及F2∶3群体共检测到剑叶长QTL有12个,分布于第1、2、3、4、5、7、10、11、12染色体,其对表型的贡献率从0.18%至12.08%不等。剑叶长在中嘉早17/D50 F2及F2∶3群体中均检测到的QTL有2个,qFLL-2和qFLL-7-2,分别位于第2和第7染色体;位于第2染色体的QTL-qFLL-2,其等位基因加性效应来自D50,位于第7染色体的QTL-qFLL-7-2,来自亲本中嘉早17的等位基因使其剑叶长度增加。
3)剑叶宽。在中嘉早17/D50 F2及F2∶3群体共检测到剑叶宽的QTL有8个,分别位于第1、3、4、10、11和12染色体上,其对表型的贡献率从0.48%至20.45%不等。剑叶宽在中嘉早17/D50 F2及F2∶3群体中均检测到的QTL为1个,位于第4染色体,对表型的贡献率在F2和F2∶3群体分别为14.37%和9.04%,来自亲本中嘉早17的等位基因能使其剑叶宽增加。
4)有效穗数。在中嘉早17/D50 F2及F2∶3群体共检测到有效穗数的QTL有6个,分别位于第1、6、7、10染色体上,其对表型的贡献率从0.16%至9.72%不等;其加性效应大多数来自亲本中嘉早17。
5)穗长。在中嘉早17/D50 F2及F2∶3群体共检测到穗长的QTL有12个,其贡献率变幅为0.34%~17.6%,在F2和F2∶3群体中均能检测到的穗长QTL为1个,位于第3染色体,其贡献率分别为0.34%和7.48%;来自亲本中嘉早17的等位基因能增加其穗长。
6)单株总粒数。在中嘉早17/D50 F2及F2∶3群体共检测到单株总粒数QTL有5个,分别位于第3、6、7、12染色体上,其对表型贡献率变幅为3.31%~9.90%。
7)结实率。在中嘉早17/D50 F2及F2∶3群体共检测到控制结实率的QTL有8个,分别位于第1、3、4、5、6、8、11染色体上,其对表型的贡献率变幅为1.65%~13.25%。
8)千粒重。在中嘉早17/D50 F2及F2∶3群体共检测到控制千粒重的QTL有5个,分别位于第2、3、6、10、12染色体上,其对表型的贡献率变幅为0.08%~17.33%。在中嘉早17/D50 F2与F2∶3群体中均检测到的控制千粒重的QTL为3个,分别位于第2、3、6染色体上,使千粒重增加的等位基因均来自亲本中嘉早17。
3讨论
水稻产量相关性状大多属于数量性状,其遗传特性除了受基因位点本身的遗传力控制外,还受外界环境、内部其他基因以及不同世代遗传背景的影响,因而数量性状的遗传比较复杂[18,19]。本研究以中嘉早17/D50 F2和F2∶3为研究群体,在不同年际间、不同世代群体分析中嘉早17的高产QTL,理论上可以减少外界环境、内部其他基因以及遗传背景对水稻产量相关性状的影响。
本研究中,中嘉早17/D50 F2和F2∶3群体中共检测到产量相关QTL有66个,其中在F2和F2∶3群体中均检测到的QTL有13个,在这13个QTL中效应值较大的有8个,包括株高:qPH-3-2、qPH-6、qPH-10;剑叶长QTLqFLL-2;剑叶宽QTLqFLW-4;千粒重QTLqTGW-2、qTGW-3、qTGW-6。在这8个不同年际间、不同世代群体稳定遗传且效应值较大的QTL中,某些QTL区间包含已经克隆的产量相关基因,其中株高的qPH-3-2与罗炬等[14]之前报道过的株高QTLqPH3同属一个区域;qPH-6区域内包含已经被克隆的抽穗期基因d35[20];qPH-10定位区间内包含已被克隆的影响多个产量相关性状基因brd2[21]。剑叶宽qFLW-4的区间已覆盖之前报道的、已被克隆的影响叶宽及产量的LSCHL4位点[22]。另外,千粒重qTGW-2、qTGW-3的检测区间分别覆盖已经克隆的影响水稻千粒重和粒形的基因GW2[12]和GS3[11];qTGW-6与此前报道过的被精细定位的GW6属同一区域[23]。表明超级稻品种中嘉早17携带许多与高产相关的基因位点。
前人研究表明,虽然数量性状遗传受外界环境影响较大,但某些遗传力较大的QTL在年际间还是能被重复检测到[24-25]。在本研究中也同样存在此现象。然而在本研究中某些在年际间、不同世代重复检测到的QTL倾向于聚集在同一区域,如在第3染色体上RM1373与RM7389之间同时检测到株高、剑叶长和穗长的QTL;在第7染色体上RM118与RM248之间同时也检测到控制株高、剑叶长和穗长的QTL;另外,在第10染色体上RM1375与RM25664之间同时检测到控制株高、剑叶长、剑叶宽、穗长和有效穗数的QTL。对于控制多个数量性状的QTL聚集于同一染色体区段的原因,学者一般认为有两种可能,即某个重要QTL的功能存在同时控制水稻多个农艺性状的作用;另外是控制相关性状的QTL呈连锁分布状态位于同一染色体区域[26-27];这就能阐释在水稻遗传育种过程中,往往欲改良某个性状,但同时会引入其他不想要的劣质性状且很难剔除。就本研究中检测到的相关性状QTL聚集区域,其各性状之间相关性极显著,因而很可能在这些QTL聚集区域内存在同时控制多个农艺性状的QTL,而非不同的QTL呈连锁分布形式。
对水稻产量相关性状QTL分析,其最终目的是为了克隆产量相关的QTL;此外,针对在年际间、不同世代稳定表达的效应值较大的重要农艺性状QTL开发相应的分子标记,利用分子标记辅助选择对一些表型难以察觉的性状可快速决选,进而加快育种进程[28]。本研究中检测到多个在年际间、不同世代稳定表达的产量相关QTL,而且某些检测到的QTL区间包含已被克隆的产量相关基因,因而可根据检测到的产量相关QTL进一步精细定位,进而开发相应的功能标记,并利用这些功能标记在今后的育种实践中快速高效选择携带中嘉早17产量相关的优良基因,进而快速培育产量更高、综合性状更优良的超级稻新品种。
参考文献:
[1] Normile D. Reinventing rice to feed the world.Science, 2008, 321: 330-333.
[2] 程式华. 中国超级稻育种. 北京: 科学出版社, 2010.
Cheng S H. The Super Rice Breeding of China. Beijing: Science press, 2010: 1.
[3] 李绍波, 杨国华, 章志宏, 等. 水稻产量要素相关性状QTL定位. 武汉大学学报:理学版, 2008, 54(6): 713-718.
Li S B, Yang G H, Zhang Z H, et al. Mapping of QTL controlling yield component related traits in rice.JWuhanUniv, 2008, 54(6): 713-718. (in Chinese with English abstract)
[4] 刁林林, 赵宏伟, 王敬国, 等. 水稻重要农艺性状的QTL定位. 东北农业大学学报, 2012, 43(1): 48-54.
Diao L L, Zhao H W, Wang J G, et al. QTL analysis of important agronomic traits in rice.JNortheastAgricUniv, 2012, 43(1): 48-54. (in Chinese with English abstract)
[5] 许凌, 张亚东, 朱镇, 等. 不同年份水稻产量性状的QTL分析. 中国水稻科学, 2008, 22(4): 370-376.
Xu L, Zhang Y D, Zhu Z, et al. Dissection of QTLs in two years for yield component traits in rice (Oryzasativa).ChinJRiceSci, 2008, 22(4): 370-376. (in Chinese with English abstract)
[6] 占小登, 于萍, 林泽川, 等. 利用大粒籼/小粒粳重组自交系定位水稻生育期及产量相关性状QTL. 中国水稻科学, 2014, 28(6): 570-580.
Zhan X D, Yu P, Lin Z C, et al. QTL Mapping of heading date and yield-related traits in rice using a recombination inbred lines (RILs) population derived from BG1/XLJ.ChinJRiceSci, 2014, 28(6): 570-580. (in Chinese with English abstract)
[7] 王兰, 李智, 郑杏梅, 等. 普通野生稻矮化突变体的株高与分蘖基因的QTL定位及主效基因的遗传分析. 华北农学报, 2014, 29(5): 5-9.
Wang L, Li Z, Zheng X M, et al. Mapping quantitative trait loci associated with height and tillers ofOryzarufipogonGriff. dwarf mutant and genetic analysis of major quantitative locus.ActaAgricBorealiSim, 2014, 29(5):5-9. (in Chinese with English abstract)
[8] 陈明亮, 熊焕金, 胡兰香, 等. 水稻产量相关数量性状基因研究进展. 江西农业学报, 2014, 26(12): 16-20.
Chen M L, Xiong H J, Hu L X, et al. Research advances in genes of yield-related quantitative traits in rice.ActaAgricJiangxi, 2014, 26(12):16-20. (in Chinese with English abstract)
[9] Xue W Y, Xing Y Z, Weng X Y, et al. Natural variation inGhd7 is an important regulator of heading date and yield potential in rice.NatGenet, 2008, 40(6): 761-767.
[10]Fan C C, Xing Y Z, Mao H L, et al.GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein.TheorApplGenet, 2006, 112(6): 1164-1171.
[11]Mao H L, Sun S Y, Yao J L, et al. Linking differential domain functions of theGS3 protein to natural variation of grain size in rice.ProcNatAcadSciUSA, 2010, 107(45): 19579-19584.
[12]Song X J, Huang W, Shi M, et al. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase.NatGenet,2007, 39: 623-630.
[13]Orjuela J, Garavito A, Bouniol M, et al. A universal core genetic map for rice.TheorApplGenet, 2010, 120: 563-572.
[14]罗炬, 邵高能, 魏祥进,等. 一个控制水稻株高QTLqPH3的遗传分析. 中国水稻科学, 2012, 26(4):417-422.
Luo J, Shao G N, Wei X J, et al. Genetic analysis of a QTLqPH3 for plant height in rice.ChinJRiceSci, 2012, 26(4): 417-422. (in Chinese with English abstract)
[15]van Ooijen J W. Join Map 4, software for the calculation of genetic linkage maps in experimental populations. Wageningen, Netherlands: 2006.
[16]Wang S, Basten C J, Zeng Z B. Windows QTL cartographer 2.5 department of statistics. Raleigh, USA: North Carolina State University, 2006.
[17]McCouch S R. Gene nomenclature system for rice.Rice, 2008, 1: 72-84.
[18]赵建国, 蒋开锋, 杨莉, 等. 水稻产量相关性状QTL定位. 中国水稻科学, 2013, 27(4): 344-352.
Zhao J G, Jiang K F, Yang L, et al. QTL mapping for yield related components in a RIL population of rice.ChinJRiceSci, 2013, 27(4): 344-352. (in Chinese with English abstract)
[19]Liu G F, Yang J, Zhu J. Mapping QTL for biomass yield and its components in rice (OryzasativaL.).ActGeneSini, 2006, 33(7): 607-616.
[20]Hironori I, Tomoko T, Tomoaki S, et al. A rice semi-dwarf gene, Tan-Ginbozu (D35), encodes the gibberellin biosynthesis enzyme, ent-kaurene oxidase.PlantMolBiol, 2004, 54: 533-547.
[21]Hong Z, Miyako Ueguchi-Tanaka, Shozo F, et al. The rice brassinosteroid-deficient dwarf 2 mutant, defective in the rice homolog of arabidopsis DIMINUTO/DWARF1, is rescued by the endogenously accumulated alternative bioactive brassinosteroid, dolichosterone.PlantCell, 2005, 17, 2243-2254.
[22]Zhang G H, Li S Y, Wang L, et al.LSCHL4 from japonica cultivar, which is allelic to NAL1, increases yield of Indica super rice 93-11.MolPlant, 2014, 7: 1350-1364.
[23]Li Y Y, Tao H J, Zhao X Q, et al. Molecular improvement of grain weight and yield in rice by usingGW6 gene.RiceSci, 2014, 21(3):127-132.
[24]李孝琼, 韦宇高, 国庆, 等. 水稻遗传图谱构建及粒形相关性状的QTL定位. 南方农业学报, 2014, 45(7): 1154-1159.
Li X Q, Wei Y, Gao G Q, et al. Construction of genetic map and mapping quantitative trait loci for grain shape-related traits in rice.JSouthernAgric, 2014, 45(7):1154-1159. (in Chinese with English abstract)
[25]张向阳, 张红宇, 徐培洲, 等. 重穗型水稻穗部性状及剑叶宽的QTL定位. 杂交水稻, 2014, 29(6): 56-61.
Zhang X Y, Zhang H Y, Xu P Z, et al. QTL mapping of flag leaf width and panicle traits of heavy-panicle rice.HybridRice, 2014, 29(6): 56-61. (in Chinese with English abstract)
[26]陈燕华, 黄大辉, 邱永福, 等. 水稻主要农艺性状的QTL 分析. 华南农业大学学报, 2014, 35(5): 42-51.
Chen Y H, Huang D H, Qui Y F, et al. A QTL analysis of main agronomic characters in rice,Oryzasativa.JSouthChinaAgricUniv, 2014, 35(5):42-51. (in Chinese with English abstract)
[27]Kazumitsu O, Horiuchi Y, Noriko I, et al. A QTL cluster for plant architecture and its ecological significance in Asian wild rice.BreedingSci, 2007, 57(1): 7-16.
[28]Yang J, Hu C, Hu H, et al. QTL Network: Mapping and visualizing genetic architecture of complex traits in experimental populations.Bioinformatics,2008, 24: 721-723.
QTL Mapping of Yield Related Traits in Super Rice Variety Zhongjiazao 17
SHENG Zhong-hua1, #, ZHU Zi-liang2, #, MA Ning1, LI Wei2, HE Ji-wai1, WEI Xiang-jin1, SHAO Gao-neng1, WANG Jian-long2, HU Pei-song1, TANG Shao-qing1,*
(1ChinaNationalRiceResearchInstitute,Hangzhou310006,China;2CollegeofAgronomy,HunanAgriculturalUniversity,Changsha410128,China;#Theseauthorscontributedequallytothiswork;*Correspondingauthor,E-mail:sqtang@126.com)
SHENG Zhonghua, ZHU Ziliang, MA Ning, et al. QTL mapping of yield related traits in super rice variety Zhongjiazao 17. Chin J Rice Sci, 2016, 30(1): 35-43.
Abstract:The rice yield related agronomic traits were examined, and the genetic linkage maps were constructed with the F2 populations derived from Zhongjiazao 17 × D50. A total of 66 QTLs for yield related traits were mapped, with the contribution rate ranging from 0.08%~20.45%, distributed on all the 12 chromosomes except for chromosome 9. Numbers of QTLs for yield related traits were mapped in the same intervals on Chr.3, Chr.7 and Chr.10,respectively, in F2 or F(2∶3) populations, and the positive alleles were contributed by Zhongjiazao 17. Among the mapped 66 QTLs,these detected in both F2 and F(2∶3) populations with high contribution rate include qPH-10, qFLW-4, qTGW-2,of which the positive alleles were also contributed by Zhongjiazao 17. In total, 66 QTLs for yield related traits were preliminarily examined in super rice variety Zhongjiazao 17, which will provide theoretical and technical supports to breed new super rice varieties with higher yield and better comprehensive characteristics by transfering excellent genes from Zhongjiazao 17 into other rice varieties.
Key words:QTL mapping; yield related trait; super rice
文章编号:1001-7216(2016)01-0035-09
中图分类号:Q343.1+5; S511.032
文献标识码:A
基金项目:浙江省自然科学基金资助项目(LY14C130012); 浙江省公益基金资助项目(2015C32045)。
收稿日期:2015-04-08; 修改稿收到日期: 2015-07-20。
