茶树中奎尼酸/莽草酸-羟肉桂酰基转移酶基因的克隆及功能研究
2016-02-06魏丽
魏 丽
(山东英才学院,山东 济南 250104)
茶树中奎尼酸/莽草酸-羟肉桂酰基转移酶基因的克隆及功能研究
魏 丽
(山东英才学院,山东 济南 250104)
从茶树中分离到1个绿原酸生物合成相关基因,命名为CsHCT。该基因编码1个430个氨基酸的多肽,推测分子质量约为48.6 ku。蛋白质序列比对表明,CsHCT含有酰基转移酶的保守域HXXXD和DFGWG,与其他物种HCT的同源性在58%~82%。系统发育分析表明,CsHCT与咖啡中的HCT同源性最高,连同其他植物HCT形成一个独立的组群。通过大肠杆菌异源表达,纯化获得了CsHCT蛋白,并进行了催化活性研究。结果表明,重组的CsHCT能有效催化奎尼酸和莽草酸与香豆酰辅酶A的转酰基反应,分别生成香豆酰奎尼酸和香豆酰莽草酸。CsHCT不能催化奎尼酸和咖啡酰辅酶A生成绿原酸。
茶树; 绿原酸; 奎尼酸/莽草酸-羟肉桂酰基转移酶
绿原酸是重要的抗氧化剂,广泛存在于金银花、苹果、番茄、马铃薯、茄子中[1-4]。除了对人类健康有益外,绿原酸也是非常重要的植物代谢产物[5]。绿原酸是G型和S型木质素的合成前体,并参与植物细胞对环境的应激反应[6]。由于绿原酸的重要作用,近些年对其生物合成的研究越来越多。该合成途径的前3步已经明确[7-9]。但接下来的几个步骤有待证实。其中有2个酰基转移酶可能与绿原酸的合成有关,一个为奎尼酸/莽草酸-羟肉桂酰基转移酶(HCT),另一个为奎尼酸羟基肉桂酰基转移酶(HQT)。HCT已从烟草中获得,对其活性分析表明,该蛋白质能够催化莽草酸或奎尼酸与香豆酸和咖啡酸成酯,对莽草酸的亲和力更高[10]。此外,还发现烟草HCT能催化绿原酸水解,形成咖啡酰辅酶A和奎宁酸。最近的功能分析表明,拟南芥中HCT基因沉默,导致木质素合成受阻,植株矮化,表明HCT是木质素合成的关键酶[11]。HQT基因已先后在烟草和番茄中被克隆[11]。研究发现,重组的烟草HQT能够催化奎尼酸到绿原酸的可逆反应。番茄中过表达HQT基因,叶片中绿原酸含量增加了85%,而HQT沉默引起叶片中的绿原酸含量减少了98%[11]。
茶树是重要的经济作物,其抗逆性是育种的重要目标之一。研究表明,绿原酸对茶蚜及鳞翅目害虫有一定的毒性,幼虫取食后,食欲大减,发育迟缓,甚至停止生长,很少正常产卵。在受到螨类危害时,一些茶树品种体内的绿原酸含量迅速增加。所以,茶树体内绿原酸含量的多少可作为判断茶树抗虫性强弱的重要依据之一。目前,关于茶树中绿原酸生物合成的研究未见报道。为揭示茶树中绿原酸的生物合成,本研究首次克隆了茶树HCT基因(CsHCT),并通过体外酶促反应,分析其催化特性,以期为茶树抗虫机制的研究奠定基础。
1 材料和方法
1.1 材料
供试茶树品种为竹山1号1年生盆栽茶苗,置于日光温室,生长条件为温度(25±2)℃、湿度70%±5%。盆栽的基质成分和比例为蛭石∶珍珠岩∶泥碳=1∶1∶4。
1.2CsHCT基因的分离
使用Autolabtech 植物总RNA提取试剂盒(天根生化科技有限公司,北京)从茶树叶片中提取总RNA。之后使用ImProm-IITM反转录试剂盒 (Promega WI,美国) 以及反转录特异引物RRP:5′-CCAGTGAGCAGAGTGACGAGGACTCGAGCTCAAG-CTTTTTTTTTTTTTTTTT-3′进行反转录。简并引物的设计参照“保守-简并杂合寡核苷酸引物”(CODEHOP)策略,设计引物CODF 5′-GTYGGHAACCTCTACGAC-3′和 CODR 5′-RTCWCKVGCNACHGCCCA-3′,其中R代表A或G,Y代表C或T,H代表A或C或T。以反转录产物为模板,使用上述引物进行PCR扩增获得CsHCT基因cDNA的核心片段,PCR扩增程序为:95 ℃变性3 min;94 ℃ 30 s、55 ℃ 40 s、72 ℃ 90 s ,30个循环;72 ℃延伸10 min。全长cDNA序列通过cDNA末端快速克隆技术(RACE)获得。进行3′-RACE的2个特异引物根据已经获得的核心片段序列设计如下,HCT3-1:5′-AAATAGATCTTGAAAGCACGAG-3′ ;HCT3-2:5′-GATAAAGCAGGAAGGACACACC-3′。第1轮PCR扩增使用HCT3-1和RRP引物组合,第2轮PCR扩增使用HCT3-2和RRP引物组合。具体的PCR扩增程序为:95 ℃变性3 min;94 ℃ 30 s、55 ℃ 40 s、72 ℃ 60 s,30个循环;72 ℃延伸7 min。cDNA 5′末端的扩增使用5′-RACE试剂盒 (Invitrogen, CA, 美国),2个用于5′-RACE扩增的特异引物依照已经获得的核心片断进行设计:5′-GTTCGCGATATTCCTCTG-3′ (HCT5-1)和5′-TTGTTGATGCTACGACCG-3′ (HCT5-2)。PCR扩增条件与3′-RACE相同。
1.3 异源表达和蛋白质纯化
CsHCTcDNA的ORF使用含有特定限制性内切酶位点的N-末端和C-末端引物进行PCR扩增。正义引物为:5′-TATAGAATTCATGGCATTCGCTC-TCCTC-3′,下划线为EcoRⅠ酶切位点;反义引物为:5′-TATACTCGAGCTAGTAATCCACTGGGACACG-3′,下划线为XhoⅠ酶切位点。扩增片段经过EcoRⅠ/XhoⅠ双酶切后,连接到经过相同酶切的原核表达载体pET-30a(+) (Novagen, Darmstadt, 德国)上,组氨酸六聚体标签构筑在重组蛋白的C-末端上。重组质粒转化BL21(DE3) (天根生化科技有限公司,北京), 挑取单克隆于37 ℃、200 mL LB培养基(含卡那霉素50 mg/L、氯霉素34 mg/L)中振荡培养。待OD600为0.6~0.8时加入1 mmol/L IPTG,28 ℃振荡培养4 h。离心收集菌体,用3 mL 0.1 mol/L磷酸缓冲液 (pH值7.5)重悬,冰上超声破碎10 min。冰上破碎的菌体匀浆于4 ℃ 10 000×g下离心10 min。上清通过Ni2+-琼脂糖柱(Novagen),用含有0.5 mol/L NaCl和40 mmol/L咪唑的0.1 mol/L磷酸钾缓冲液洗涤Ni2+-琼脂糖柱3次后,用含有400 mmol/L 咪唑的0.1 mol/L磷酸钾缓冲液洗脱重组目标蛋白。为了长期保持重组蛋白活性,洗脱缓冲液通过PD-10柱(Amersham Pharmacia Biotech,Uppsala,瑞典),用含10%(V/V)甘油的0.1 mol/L Tris-HCl (pH值7.5)置换缓冲液重新洗脱,洗脱的重组蛋白保存于-80 ℃。重组蛋白的纯化效率用SDS-PAGE检测。采用Bradford法测定蛋白质浓度。
1.4 体外酶促反应及产物分析
20 μL体外酶促反应体系中含有100 mmol/L 磷酸buffer (pH值7.5)、1 mmol/L二硫苏糖醇、1 μg 重组HCT蛋白、0.1~5 mmol/L的不同底物(香豆酰辅酶A、咖啡酰辅酶A、奎尼酸和莽草酸) 。反应自加入酶蛋白开始计时,30 ℃ 条件下反应30 min,加入20 μL 乙腈/HCl (99∶1)终止反应。
HPLC检测条件:C18 柱 (LiChroCART 125-4, Merck), 流动相A:90% H2O、9.9% CH3CN、0.1% HCOOH;B:80% CH3CN、19.9% H2O、0.1% CH3COOH。流速为0.6 mL/min,使用梯度条件为30% B 3 min、30%~70% B 27 min、70%~80% B 2 min、80%~95% B 3 min、95% B 5 min,检测波长320 nm。用相应的标准品对各种化合物进行定量。
1.5 动力学分析
本研究重组蛋白动力学常数的测定是在一个底物浓度饱和的情况下,用另一个底物介于其Km的0.2~6.0的5个浓度值进行计算获得的。试验使用上述标准的250 μL 0.1 mol/L 磷酸钾缓冲液反应体系,每个试验重复3次。各参数的测定在其主产物的最适反应温度和最适pH值条件下完成。Km和Kcat值由Lineweaver-Burke曲线结果得出。
2 结果与分析
2.1CsHCTcDNA的分离
利用简并引物,以茶树叶片cDNA为模板,通过PCR扩增,获得了1个 750 bp的中间序列(图1)。通过RACE方法,克隆了该基因的全长cDNA序列,命名为CsHCT。该cDNA全长1 417 bp,其中编码区为1 293 bp,推测其编码1个430个氨基酸的多肽,分子质量约为48.6 ku。蛋白质序列比对表明,CsHCT与其他物种HCT的同源性在58%—82%,含有酰基转移酶的保守域HXXXD(氨基酸153—157)和DFGWG(氨基酸377—381)(图2)。系统发育分析结果(图3)表明,CsHCT与咖啡(Cof.canephoraL.)中的HCT (ABO77957)聚合为1个分支,连同其他植物HCT形成1个独立的组群;另1个组群则由典型的HQT构成;来自于朝鲜蓟(CynaracardunculusL.)的HQT(ACF37072)形成1个独立的分支。

M.DNA分子质量标准; 1.PCR产物

图2 茶树CsHCT与其他植物HCT的氨基酸序列比对分析

图3 茶树CsHCT的系统发育分析
2.2 CsHCT的功能鉴定
为研究CsHCT的酶学特性,构建了CsHCT的pET-30a原核表达载体(图4A),经PCR(图4B)及酶切验证(图4C)后,导入大肠杆菌BL21(DE3)。经IPTG诱导,C-末端添加组氨酸标签的重组CsHCT在大肠杆菌中实现了体外表达。SDS-PAGE结果显示,纯化的重组蛋白在50 ku的位置上形成1个单一条带 (图4D)。
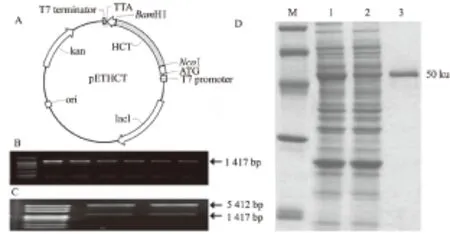
A.原核表达载体构建;B.表达载体PCR检测;C.表达载体酶切验证;D.CsHCT蛋白诱导表达及纯化,
取纯化的重组茶树CsHCT蛋白,加入到体外催化反应体系中。酶促产物的定性及定量分析结果显示,重组的CsHCT能有效催化奎尼酸和莽草酸与香豆酰辅酶A的转酰基反应,分别生成香豆酰奎尼酸和香豆酰莽草酸(图5),而对照(反应体系中不含CsHCT)没有反应产物出现。CsHCT不能催化奎尼酸和咖啡酰辅酶A生成绿原酸。
2.3 CsHCT酶学特性分析
分别以咖啡酰辅酶A和香豆酰辅酶A为酰基供体,以奎尼酸和莽草酸为酰基受体,进行了CsHCT的酶动力学研究。结果表明,CsHCT对莽草酸的亲和力更高(表1)。在香豆酰辅酶A饱和时,CsHCT对莽草酸的亲和力[Km=(83±37)μmol/L]约是奎尼酸[Km=(239±41)μmol/L]的3倍。
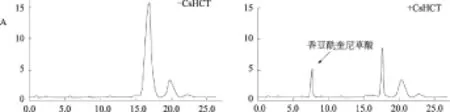
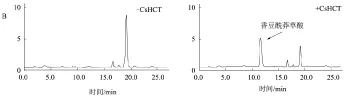
A.以奎尼酸和香豆酰辅酶A为底物;B.以莽草酸和香豆酰辅酶A为底物

表1 CsHCT的酶动力学分析
注:NA表示反应不能发生。
3 结论与讨论
本研究从茶树叶片中分离到1个酰基转移酶基因CsHCT,其编码1个430个氨基酸的多肽,推测的分子质量约为48.6 ku。该蛋白含有酰基转移酶家族的2个保守域HXXXD(氨基酸153—157)和DFGWG(氨基酸377—381),与咖啡中的1个HCT蛋白同源性最高,相似度82%。体外酶促反应表明,CsHCT能有效催化奎尼酸和莽草酸与香豆酰辅酶A的转酰基反应,分别生成香豆酰奎尼酸和香豆酰莽草酸,但不能催化奎尼酸和咖啡酰辅酶A生成绿原酸。
关于绿原酸的生物合成途径,目前认为可能有3种:(1)以咖啡酰-葡萄糖苷作为活性中间体合成绿原酸;(2)首先由酰基转移酶(HQT或HCT)催化奎尼酸与香豆酰辅酶A合成香豆酰奎尼酸,进而在香豆酸-3′-羟化酶作用下生成绿原酸;(3)酰基转移酶HCT催化莽草酸与香豆酰辅酶A合成香豆酰莽草酸,进而在香豆酸-3′-羟化酶作用下生成咖啡酰莽草酸,然后经酰基转移酶HCT催化获得咖啡酰辅酶A,最后与奎尼酸合成绿原酸。第1条途径仅在马铃薯中获得证实。Villegas等[12]从马铃薯中纯化到1个羟基肉桂酰D-葡萄糖奎尼酸酰基转移酶,该酶能够催化咖啡酰D-葡萄糖与奎尼酸生成绿原酸。途径2和途径3广泛存在于高等植物中,
是研究最多的合成途径,其主要区别在于香豆酰辅酶A发生羟化的方式不同。
植物中酰基转移酶HQT和HCT的不同催化特性,决定了该植物中绿原酸的合成方式。番茄中HCT对莽草酸的亲和力远高于奎尼酸,通过在番茄中抑制HQT基因表达,使绿原酸含量降低了98%,说明番茄中绿原酸的生物合成依赖于HQT。拟南芥中已报道的HCT不能催化奎尼酸为受体的转酰基反应,因此在拟南芥中并没有检测到绿原酸。拟南芥的hct突变体表现出木质素合成障碍,说明HCT主要参与木质素合成途径。而Hoffmann等[10]在烟草中分离到的1个HCT蛋白,能分别催化奎尼酸和莽草酸与香豆酰辅酶A或咖啡酰辅酶A的反应,推测其可能参与了2条不同的绿原酸合成途径。Comino等[13]在大叶菜蓟中也发现了类似的HCT蛋白。以上研究表明,不同物种的HCT对底物的专一性差异较大。
本研究中,CsHCT不能以咖啡酰辅酶A为底物,只能催化香豆酰酯的生成。推测CsHCT可能参与了绿原酸的生物合成,其较高的催化活性促进了茶树中绿原酸的积累。为进一步研究CsHCT在茶树绿原酸合成中的作用,今后将分别构建其过表达和RNAi载体,转化茶树,探讨基因表达与绿原酸积累的关系。
[1] Tamagnone L,Merida A,Parr A,etal.The AmMYB308 and AmMYB330 transcription factors fromAntirrhinumregulate phenylpropanoid and lignin biosynthesis in transgenic tobacco[J].Plant Cell,1998,10(2):135-154.
[2] Clifford M N.Chlorogenic acids and other cinnamates:Nature,occurrence,and dietary burden[J].J Sci Food Agric,1999,79(3):362-372.
[3] Wang M,Simon J E,Aviles I F,etal.Analysis of antioxidative phenolic compounds in artichoke(CynarascolymusL.)[J].J Agric Food Chem,2003,51(3):601-608.
[4] Schütz K,Kammerer D,Carle R,etal.Identification and quantification of caffeoylquinic acids and flavonoids from artichoke(CynarascolymusL.) heads,juice,and pomace by HPLC-DAD-ESI/MS(n)[J].J Agric Food Chem,2004,52(13):4090-4096.
[5] Abdulrazzak N,Pollet B,Ehlting J,etal.A coumaroyl-ester-3-hydroxylase insertion mutant reveals the existence of nonredundant meta-hydroxylation pathways and essential roles for phenolic precursors in cell expansion and plant growth[J].Plant Physiol,2006,140(1):30-48.
[6] Niggeweg R,Michael A J,Martin C.Engineering plants with increased levels of the antioxidant chlorogenic acid [J].Nat Biotechnol,2004,22(6):746-754.
[7] Lepelley M,Cheminade G,Tremillon N,etal.Chlorogenic acid synthesis in coffee:An analysis of CGA content and real-time RT-PCR expression of HCT,HQT,C3H1,and CCoAOMT1 genes during grain development inC.canephora[J].Plant Sci,2007,172(5):978-996.
[8] Dixon R A,Paiva N L.Stress-induced phenylpropanoid metabolism[J].Plant Cell,1995,7(7):1085-1097.
[9] Douglas C J.Phenylpropanoid metabolism and lignin biosynthesis:From weeds to trees[J].Trends Plant Sci,1996,1:171-178.
[10] Hoffmann L,Maury S,Martz F,etal.Purification,cloning,and properties of an acyltransferase controlling shikimate and quinate ester intermediates in phenylpropanoid metabolism[J].J Biol Chem,2003,278(1):95-103.
[11] Hoffmann L,Besseau S,Geoffroy P,etal.Silencing of hydroxycinnamoy-coenzymeashikimate/quinate hydroxycinnamoyltransferase affects phenylpropanoid biosynthesis[J].Plant Cell,2004,16(6):1446-1465.
[12] Villegas R A,Kojima M.Purification and characterization of hydroxycinnamoyl D-glucose:Quinate hydroxycinnamoyl transferase in the root of sweet potato,IpomoeabatatasLam.[J].The Journal of Biological Chemistry,1986,261(6):8729-8733.
[13] Comino C,Lanteri S,Portis E,etal.Isolation and functional characterization of a cDNA coding a hydroxycinnamoyltransferase involved in phenylpropanoid biosynthesis inCynaracardunculusL.[J].MBC Plant Boil,2007,7:14.
Cloning and Function Analysis of Hydroxycinnamoyl-CoA Quinate/Shikimate Hydroxycinnamoyl Transferase Gene inCamelliasinensis
WEI Li
(Shandong Yingcai University,Ji’nan 250104,China)
A hydroxycinnamoyl-CoA quinate/shikimate hydroxycinnamoyl transferase cDNA(CsHCT) was cloned fromCamelliasinensis.CsHCTencoded a protein of 430 amino acids,which molecular weight was speculated 48.6 ku.Sequencing results showed CsHCT contained two conserved acyltransferase domain HXXXD and DFGWG,and the homology between CsHCT and other plants HCTs ranged from 58%—82%.Phylogenetic analysis showed that the homology between CsHCT and HCT from coffee was the highest,composing a group with other plants HCTs.CsHCT protein was purified after heterologous expression inE.coli.Catalytic activity analysis showed that the recombinant CsHCT was capable of catalyzing quinic acid and shikimic acid to generate coumaroyl quinic and coumaroy shikimate;but CsHCT could not catalyze quinic acid and caffeoyl-CoA to generate CGA.
Camelliasinensis; chlorogenic acid; hydroxycinnamoyl-CoA quinate/shikimate hydroxycinnamoyl transferase
2016-04-06
国家自然科学基金项目(31200226)
魏 丽(1980-),女,山东菏泽人,副教授,硕士,主要从事抗性生理研究。E-mail:05300532@163.com
S722.3+6
A
1004-3268(2016)10-0035-06
