丹毒丝菌SpaA基因免疫保护区的克隆及其在毕赤酵母中的表达
2016-01-25蒋志琼钟泽民谭博敏余希尧黄毓茂华南农业大学兽医学院广东广州510642
蒋志琼,钟泽民,谭博敏,余希尧,黄毓茂(华南农业大学兽医学院,广东广州510642)
丹毒丝菌SpaA基因免疫保护区的克隆及其在毕赤酵母中的表达
蒋志琼,钟泽民,谭博敏,余希尧,黄毓茂
(华南农业大学兽医学院,广东广州510642)
摘要:【目的】以毕赤酵母Pichia pastoris X-33为宿主表达猪丹毒丝菌Erysipelothrix rhusiopathiae SpaA基因氨基端的免疫保护区蛋白.【方法】以采集于广东某猪场的猪丹毒丝菌为模板,根据NCBI中已发表的Spa基因cDNA序列设计1对引物,通过PCR扩增得到SpaA-N,并将其连接到表达载体pPICZαC上,得到重组表达质粒pPICZαC-SpaA-N; 用Sac I酶将重组表达质粒pPICZαC-SpaA-N线性化后电转化入毕赤酵母X-33,经含博来霉素ZencinTM抗性的YPDS平板筛选和PCR鉴定的阳性转化子,用含不同浓度博来霉素抗性的YPDS平板筛选出高拷贝子并进行甲醇诱导培养,分别于诱导48、72、96 h后离心收集上清液,立即做SDS-PAGE,并进行SDS-PAGE及Western-blot试验.【结果和结论】成功克隆并表达了SpaA-N基因,构建了重组表达质粒pPICZαC-SpaA-N,并以毕赤酵母X-33为宿主成功表达了猪丹毒丝菌SpaA基因氨基端的免疫保护区蛋白.丹毒丝菌SpaA-N作为免疫保护区在酵母宿主中得以成功表达.
关键词:丹毒丝菌; SpaA基因;毕赤酵母;克隆和表达;亚单位疫苗
蒋志琼,钟泽民,谭博敏,等.丹毒丝菌SpaA基因免疫保护区的克隆及其在毕赤酵母中的表达[J].华南农业大学学报,2015,36(3) : 20-25.
优先出版时间:2015-04-14
优先出版网址: http: / /www.cnki.net/kcms/detail/44.1110.s.20150414.0925.002.html
丹毒丝菌Erysipelothrix rhusiopathiae属于丹毒丝菌属,是一种兼性厌氧、无芽孢、不产酸的纤小革兰阳性杆菌[1],是引起猪丹毒和人类丹毒症的病原体[2].该菌能够在各种环境中长期存在(包括海中)[3],并且能够感染各种野生动物和家畜、鸟类和鱼类[4-8],从而引发疾病(鱼除外),该病在全球广泛存在,呈散发或流行性发生,特别是能够引起猪的急性热性败血症、亚急性疹块、慢性关节炎和心内膜炎,给养猪业造成了巨大的损失[9].目前国内猪丹毒病仍时有发生[10-14],虽然感染阳性率不高,但是猪丹毒的阳性检出率却比较高[15],因此猪丹毒的隐性感染值得关注.目前,常用防治猪丹毒的方法还是传统疫苗,虽然此类疫苗在一定程度上起到了相应的作用,但对慢性猪丹毒的作用效果并不强,而且还在很大程度上增加了猪慢性关节炎的发病率[16-17],因此研究一种新型猪丹毒疫苗迫在眉睫.
近年来国内外学者对猪丹毒免疫保护性蛋白(rSpa)进行了大量的研究,为猪丹毒的诊断和防治提供了科学的理论依据.其中表面保护抗原A (SpaA)由N端的免疫保护性区域和C端的细胞结合区域组成,大量研究表明其具有良好的免疫保护功能[8,18-20].而SpaA蛋白中起主要免疫保护效应的核心区段为氨基端(SpaA-N),其免疫保护作用受到国内外学者的广泛关注,并进行了大量的试验研究[21-25].研究者们利用原核表达技术成功地克隆出Spa-N片段基因序列,并且研究了其免疫功能[23,25].
本试验成功地扩增了Spa-N基因序列,并利用酵母表达系统对其进行表达,为Spa-N作为一种亚单位疫苗投入生产时的高产量、生长快速、易操作和低成本提供了理论依据.
1 材料与方法
1.1材料
1.1.1菌株和载体猪丹毒菌分离自广东某猪场,pPICZαC为Invitrogen公司产品,大肠埃希菌Escherichia coli DH5α为TaKaRa公司产品,毕赤酵母Pichia pastoris X-33由华南农业大学兽医学院传染病教研室保存.
1.1.2试剂与酶类质粒提取试剂盒、胶回收试剂盒、纯化试剂盒等为Omega公司产品;各限制性内切酶、T4连接酶、DNA聚合酶、DNA Marker等为TaKa-Ra公司产品; PCR引物合成及测序由Invitrogen公司完成;博莱霉素、D-生物素、酵母氮碱等为Sigma公司产品;培养基成分如蛋白胨、酵母提取物、葡萄糖、培养基以及SDS-PAGE试验所需试剂、HRP标记的羊抗鼠IgG等均为北京鼎国生物技术公司产品;其他试剂均为国产分析纯.
1.1.3培养基LB液体培养基: 10 g胰蛋白胨,5 g酵母提取物,10 g NaCl,双蒸水定容至1 L.固体LB培养基:在液体LB培养基基础上加1.5%琼脂粉.血清琼脂培养基:在普通LB培养基的配方基础上,双蒸水定容至90 mL后混匀高压,待冷却至一定温度时加入10%的兔血清.YPD培养基:酵母提取物10 g,胰蛋白胨20 g,溶于900 mL双蒸水中,高压灭菌20 min,再加入100 mL已过滤0.2 g·mL-1的D-葡萄糖.YPDS培养基:酵母提取物10 g,胰蛋白胨20 g,山梨醇182.2 g,双蒸水定容至900 mL,高压20 min后加入100 mL已灭菌的0.2 g·mL-1的D-葡萄糖.BMGY培养基:酵母提取物10 g、胰蛋白胨20 g溶于700 mL双蒸水中,高压灭菌后冷却至室温,分别加入100 mL已除菌的1 mol·L-1磷酸缓冲液(pH6.0)、134 g·L-1酵母氮碱、体积分数为10%的丙三醇及20 mg·L-1的生物素2 mL.BMMY培养基:酵母提取物10 g,胰蛋白胨20 g,溶于700 mL双蒸水中,高压灭菌后冷却至室温,分别加入100 mL已除菌的1 mol·L-1磷酸缓冲液(pH6.0)、134 g·L-1酵母氮碱、体积分数为10%的甲醇10 mL及20 mg·L-1的生物素2 mL.
1.2重组毕赤酵母表达载体的建立
1.2.1引物的设计与合成根据NCBI上SpaA序列,登录号: AB259654.1,运用Primer premier 5.0软件设计1对特异性引物用于扩增SpaA-N.上游引物P1加入Xho I酶切位点;下游引物P2加入Xba I酶切位点和终止密码子.本试验预采用毕赤酵母表达载体的α因子信号肽,由于质粒pPICZαC双酶切中会丢失KEX2蛋白酶的酶切位点Lys-Arg,因此在设计上游引物的时候,还加入了编码Lys-Arg的密码子AAAAGA.引物由Invitrogen公司合成.引物序列:上游引物P1,5'- CCCTCGAGAAAAGAGGGTACCAAAGTTTCGAAGC-3';下游引物P2,5'-GC TCTAGATTAGATCTTTAGGTTTTTCTTCATCAA-3'.
1.2.2丹毒丝菌SpaA-N的PCR扩增利用煮-冻-煮法裂解猪丹毒丝菌的细胞壁,进行菌落PCR.PCR反应体系: Ex 10×PCR Buffer 2 μL,2.5 mmol dNTP Mixture 2 μL,引物P1、P2各0.5 μL,Ex Taq DNA聚合酶0.125 μL,ddH2O 17.375 μL,模板2 μL.反应条件: 94℃预变性5 min; 94℃变性30 s,54℃退火30 s,72℃延伸70 s,30个循环; 72℃终延伸10 min.经8 g·L-1琼脂糖凝胶电泳分析产物后,利用DNA回收试剂盒进行纯化回收.
1.2.3重组表达载体的构建对胶回收后的回收产物Spa-N和表达载体pPICZαC同时进行Xho I和Xba I双酶切,并利用DNA胶回收试剂盒进行纯化回收.将酶切回收后的目的基因与表达载体连接后转化大肠埃希菌DH5α感受态细胞,将最后一步的重悬液均匀涂布于含ZencinTM(25 μg·mL-1)的低盐LB固体培养基上,放入37℃培养箱中培养12~16 h.
1.2.4重组表达质粒的鉴定PCR鉴定:挑选低盐LB平板上的可疑菌落于含ZencinTM(100 μg·mL-1) LB液体培养基中37℃震荡培养8 h,做菌液PCR鉴定.PCR体系与反应条件同“1.2.2”.
单酶切和双酶切鉴定:按照相应体系,利用酶Xho I与Xba I进行双酶切鉴定,用酶Xba I进行单酶切鉴定.
1.3 SpaA-N基因在毕赤酵母中的表达
1.3.1毕赤酵母X-33感受态细胞的制备参照Invitrogen EasySelect Pichia Expression kit介绍的方法进行制备.1.3.2重组质粒pPICZαC-SpaA-N的电击转化根据Invitrogen公司实验手册进行.先利用Sac I单酶切重组质粒pPICZαC-SpaA-N,使之线性化,同时对空质粒pPICZαC也进行同酶同系线性化,反应体系(50 μL) : 10×Buffer 5 μL,pPICZαC-SpaA-N 30 μL,Sac I 3 μL,ddH2O 12 μL.将75 μL新制备的感受态酵母细胞与25 μL线性化的重组质粒轻轻混匀后转入预冷的电转杯中冰浴5 min.设定电转参数305 V,进行15 ms电转后立即加入1 mL预冷的1 mol·L-1山梨醇,混匀后转入5 mL离心管,30℃条件下静置1~2 h,再加入YPD液体培养基1 mL,30℃震荡培养1 h.4 000 r·min-1收集菌体,用50~200 μL YPD重悬菌体后铺于新鲜制备的YPDS平板(含ZencinTM100 μg·mL-1)上,将平板倒置于30℃培养箱中培养3~5 d.
1.3.3重组酵母菌菌液PCR鉴定及高拷贝转化子的筛选挑取单菌落于1 mL YPDS培养基中(含ZencinTM200 μg·mL-1)扩菌培养,采用煮-冻-煮法制备PCR模板分析毕赤酵母转化子,以P1、P2为引物扩增出1 029 bp的片段为阳性转化子.PCR体系与反应条件同“1.2.2”.再经不同浓度ZencinTM的YPDS平板筛选高拷贝克隆工程菌,用于高效诱导表达SpaA-N蛋白.
1.3.4抗血清的制备购买8只体质量都在15 g左右的SPF雌鼠.培养猪丹毒菌,用平板计数法进行计数,并用生理盐水将其稀释至10 LD50(1.1×102cfu),加入体积分数为0.1%的甲醛进行灭菌处理,划板验证为无菌后进行下一步试验.将铝胶佐剂与经过灭菌处理的猪丹毒菌液按照体积比1∶5的比例进行混合,第1次免疫时每只小鼠腹腔注射100 μL混合液,14 d后进行第2次免疫,第2次免疫10 d后进行第3次免疫,3次注射剂量均相同,在第3次免疫10 d后进行摘眼取血.采集的血液先置37℃条件下凝固约2 h,然后置4℃条件下过夜,使血块收缩,第2天5 000 r·min-1离心5 min,收集血清,于-80℃条件保存备用.
1.3.5高拷贝酵母工程菌的诱导表达和SDS-PAGE
挑取阳性单克隆转化子,接种至BMGY中,28℃摇至D600 nm为2~6,用BMMY重悬细胞至D600 nm为1.0进行诱导表达,每24 h加甲醇至甲醛终体积分数为1%以继续诱导.分别在48、72、96 h时离心收集上清液,立即做SDS-PAGE或-20℃条件保存备用,并做SDS-PAGE及Western-blot分析.
1.3.6免疫及动物保护试验将16只体质量为18~20 g的昆明小鼠随机分为2组,每组8只.将生理盐水、经过毕赤酵母表达的SpaA-N蛋白分别与铝胶佐剂以体积比5∶1进行混合,每只小鼠腹腔注射混合液100 μL进行第1次免疫,分别命名为组1、组2,其中组1为空白对照组.第1次免疫14 d后进行第2次免疫,各组的免疫剂量与方法同第1次免疫.分别对第2次免疫14 d后的2组免疫小鼠进行腹腔注射攻毒,攻毒剂量为100 LD50(1.1×103cfu),连续观察10 d,记录各组小鼠死亡数,计算各免疫组的保护率.
2 结果
2.1 Spa-N基因的合成
以菌液为模板,P1、P2为引物,进行PCR扩增,经10 g·L-1琼脂糖凝胶电泳可观察到有单一条带,与实际条带1 029 bp相符(图1).
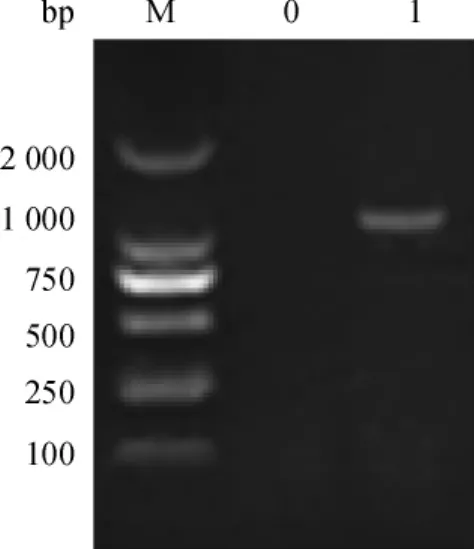
图1 Spa-N基因PCR扩增Fig.1 PCR amplification of Spa-N gene
2.2重组表达质粒的PCR鉴定
从转化后培养的低盐LB平板上挑取疑似阳性的单菌落,接种于含ZencinTM25 μg·mL-1的LB液体培养基中,37℃条件下震荡过夜,直接以菌液为模板进行PCR鉴定,以P1、P2为引物,扩增产物经10 g·L-1琼脂糖凝胶电泳,可观察到大小约为1 029 bp的单一条带(图2),而阴性对照无任何条带.结果表明猪丹毒SpaA-N已成功插入表达载体中.

图2 重组质粒pPICZαC-SpaA-N的PCR鉴定Fig.2 Identification of recombinant plasmid pPICZαC-SpaA-N by PCR
2.3重组表达质粒pPICZαC-SpaA-N的单酶切及双酶切鉴定
以抽提的质粒分别在20 μL体系中做Xho I单酶切鉴定及Xba I和Xho I的双酶切鉴定,并且以不加酶的重组表达质粒作阴性对照,于37℃恒温水浴过夜后,以1 g·L-1的琼脂糖凝胶电泳检测结果.单酶切出现1条4.6 kb左右条带,与实际相符;双酶切出现2条带,其中1条大小约为1 029 bp,与外源目的基因大小1 029 bp相符(图3).

图3 重组质粒pPICZαC-SpaA-N的酶切鉴定Fig.3 Identification of recombinant plasmid pPICZαC-SpaA-N by restriction endonucleases digestion
2.4重组表达质粒的序列测定结果
将经过PCR和酶切鉴定的阳性重组表达质粒,送Invitrogen公司,用载体引物(即通用引物)对重组质粒pPICZαC-SpaA-N进行测序.结果显示,目的片段与NCBI已发表的Spa-N序列的相似性高达99%.
2.5重组酵母菌液PCR鉴定
挑取电转的阳性菌落于1 mL的YPDS液体培养基中(含ZencinTM100 μg·mL-1),30℃条件下180 r·min-1振荡培养,取少量菌液进行PCR鉴定.同时取空载体pPICZαC/X-33转化的重组酵母菌作阴性对照,电泳检测.结果显示:重组质粒转化的重组酵母菌液可以扩增出大小约为1 029 bp的特异条带,而空质粒转化的酵母菌液在1 029 bp处无条带(图4).
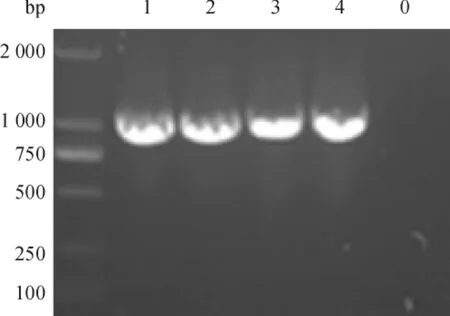
图4 重组酵母菌液PCR鉴定Fig.4 Identification of recombinant yeast by PCR
2.6 SpaA-N表达产物的SDS-PAGE分析
高拷贝重组酵母工程菌株经甲醇诱导表达,分别于诱导48、72、96 h后经10 000 r·min-1离心收集酵母上清液后进行SDS-PAGE(12%分离胶和5%浓缩胶)表达分析,考马斯亮蓝染色鉴定.结果显示,相对分子质量约为40 000的Spa-N蛋白得以表达(图5),表明猪丹毒Spa-N蛋白在酵母X-33中成功表达.

图5 Spa-N表达产物的SDS-PAGE分析Fig.5 SDS-PAGE analyses of expression products of Spa-N gene
2.7 SpaA-N表达产物的Western-blot分析
表达蛋白的Western-blot免疫印迹检测结果显示,抗天然SpaA蛋白抗体(抗血清)和通过酵母表达的SpaA-N蛋白发生了特异性反应,说明SpaA-N蛋白在毕赤酵母中得到成功的表达(图6).

图6 SpaA-N表达产物的Western-blot分析Fig.6 Immunoreactivies of expression products of SpaA-N gene by Western-blot
2.8免疫及动物保护试验结果
组1的生理盐水对照组小鼠在攻毒后表现出一些不适的症状:食欲下降、毛发起皱及精神沉郁等,第5天开始死亡,到第8天全部死亡.而组2的SpaAN蛋白试验组在攻毒的前2 d有一些不适症状,但到后期这些不适症状消失,没有出现死亡情况.试验结果表明试验所表达的SpaA-N蛋白具有免疫保护性.
3 讨论
现在世界各地猪丹毒仍频繁爆发[26],给世界各国的养猪业造成了巨大的经济损失.目前常用活菌疫苗、灭活疫苗及联苗等来预防和控制该病的发生和传播,但是接种活菌苗有可能使免疫动物变成带菌者和细菌的传播者,从而使得慢性病例逐渐增加;灭活疫苗的培养液本身是异体蛋白,易使接种动物发生过敏反应.而亚单位疫苗具有安全性高、产量高、稳定性好等优点,克服了活菌疫苗的毒力返强和灭活对猪只引起过敏反应等缺点.
近几年来,相继出现了一些对猪丹毒杆菌的分子生物学和免疫学的研究,Makino等[18]克隆和表达了猪丹毒丝菌表面保护性抗原SpaA,发现SpaA的N 端342个氨基酸残基组成的多肽承担了该蛋白的免疫保护功能.前人对猪丹毒杆菌SpaA的氨基酸进行原核表达和重组疫苗的研究结果表明,SpaA-N蛋白具有良好的免疫保护作用[21-25].但现如今的研究都是基于原核表达系统上,其表达的目的产物常以包涵体形式存在,无翻译后的修饰加工过程[27],包涵体复性困难且效率很低.而本试验采用的毕赤酵母表达系统与其他表达系统相比,有着不可比拟的优势,其既有原核生物繁殖快、易于培养、培养基廉价和试验过程简单可行等特点,又具有强有力的启动子,还可以对外源蛋白进行加工折叠和翻译后修饰[28]等真核生物表达体系的特点,而且巴斯德毕赤酵母还是一种能高效表达外源蛋白的表达系统,具有遗传稳定、表达水平高、蛋白可翻译后加工、产物可分泌、可高密度发酵等许多优点[29].
本研究成功克隆并表达了猪丹毒丝菌C43065 SpaA的N端免疫保护片段,测序结果分析发现其与NCBI上已登录的不同血清型菌株的SpaA基因的核苷酸序列的相似性高达99%,说明该基因具有很高的保守性.SDS-PAGE及Western-blot结果表明其具有免疫反应性,因此,本研究为猪丹毒新型亚单位疫苗的研制和大规模生产奠定了基础.
致谢:感谢兽医学院传染病教研室的老师和同学给予的支持和帮助!
参考文献:
[1]BROOKE C J,RILEY T V.Erysipelothrix rhusiopathiae: Bacteriology,epidemiology and clinical manifestations ofan occupational pathogen[J].J Med Microbiol,1999,48 (9) : 789-799.
[2]BOO T W,HONE R,HURLEY J.Erysipelothrix rhusiopathiae endocarditis: A preventable zoonosis[J].Ir J Med Sci,2003,172(2) : 81-82.
[3]SITT T,BOWEN L,MYRA T,et al.Cellular immune responses in cetaceans immunized with a porcine erysipelas vaccine[J].Vet Immunol Immunopathol,2010,137 (3/4) : 181-189.
[4]GARTRELL B D,ALLEY M R,MACK H,et al.Erysipelas in the critically endangered kakapo (Strigops habroptilus)[J].Avian Pathol,2005,34(5) : 383-387.
[5]HASSANEIN R,SAWADA T,KATAOKA Y,et al.Serovars of Erysipelothrix species isolated from the tonsils of healthy cattle in Japan[J].Vet Microbiol,2001,82 (1) : 97-100.
[6]BOERNER L,NEVIS K R,HINCKLEY L S,et al.Erysipelothrix septicemia in a little blue penguin (Eudyptula minor)[J].J Vet Diagn Invest,2004,16(2) : 145-149.
[7]HOLTIFIELD J L,COOPER G L,CHARLTON B R.An outbreak of erysipelas in 2-day-old poultry[J].Avian Pathol,2000,44(33) : 721-724.
[8]CHEUN H I,KAWAMOTO K,HIRAMATSU M,et al.Protective immunity of SpaA-antigen producing lactococcus lactis against Erysipelothrix rhusiopathiae intfection[J].J Appl Microbiol,2004,96(6) : 1347-1353.
[9]HASSANEIN R,SAWADA T,KATAOKA Y,et al.Pathogenicity for mice and swine of Erysipelothrix isolates from the tonsils of healthy cattle[J].Vet Microbiol,2003,91(2/3) : 231-238.
[10]朱凤琼,陈达燕,夏英杰,等.猪丹毒杆菌的分离及鉴定[J].现代农业科技,2012(4) : 316-317.
[11]邱美珍,杜丽飞,杨爱梅,等.猪丹毒杆菌临床分离鉴定与药敏试验[J].养猪,2011(6) : 111-112.
[12]郭良兴,陈克研,赵魁,等.猪丹毒杆菌的分离鉴定及耐药性试验[J].中国畜牧兽医,2011,38(4) : 199-202.
[13]车勇良,陈如敬,王隆柏,等.猪丹毒杆菌的分离鉴定及其SpaA基因的遗传变异分析[J].中国兽医学报,2011,31(11) : 1591-1593.
[14]万莉,韩国全,郭万柱,等.猪丹毒疫苗研究进展[J].猪业科学,2010(6) : 32-35.
[15]周绪斌,丹尼,李聪,等.猪丹毒———古老的传染病是否从中国规模化猪场消失了[J].农业新技术,2009 (5) : 22-24.
[16]FREDDY H,FRANK P,KOEN C,et al.Efficacy of vaccines against bacterial diseases in swine: What can we expect[J].Vet Microbiol,2004,100(3/4) : 255-268.
[17]SILVA A J,HORTA A C,IEMMA M,et al.Production of potential subunit vaccine against swine erysipelas in fedbatch cultures of E.coli BL21(DE3)[J].New Biotechnol,2009,255(6) : 187-188.
[18]MAKINO S,YAMAMOTO K,MURAKAMI S,et al.Properties of repeat domain found in a novel protective antigen,SpaA,of Erysipelothrix rhusiopathiae[J].Microb Pathog,1998,25(2) : 101-109.
[19]IMADA Y,GOJI N,ISHIKAWA H,et al.Truncated surface protective antigen (SpaA) of Erysipelothrix rhusiopathiae serotypes 1a and 2b in pigs[J].Infect Immun,1999,67(9) : 4376-4382.
[20]ALAINA L I,JAMES A R,PAUL J H.Erysipelothrix rhusiopathiae: Association of Spa-type with serotype and role in protective immunity[J].Vaccine,2010,28 (13) : 2490-2496.
[21]曹文尧.丹毒丝菌表面保护抗原A作为重组亚单位疫苗和核酸疫苗的初步研究[D].乌鲁木齐:新疆大学,2007.
[22]谭侃侃.丹毒丝菌的分离鉴定及其SpaA-N蛋白和lipo蛋白的免疫原性分析[D].长沙:湖南农业大学,2011.
[23]吾鲁木汗·那孜尔别克,张磊,何翠,等.猪丹毒丝菌天然SpaA和重组SpaA-N免疫保护效果的评价[J].微生物学报,2010,50 (3) : 367-372.
[24]李伟杰,赵耘,康凯,等.红斑丹毒丝菌SpaA抗原基因的克隆、序列分析及蛋白结构预测[J].中国兽医学报,2011,31(11) : 1591-1594.
[25]DA A J,SILVA M R,IEMMA A C,et al.Cloning,autoinduction expression,and purification of rSpaA swine erysipelas antigen[J].Curr Microbiol,2012,65(4) : 369-374.
[26]OPRIESSNIG T L,HOFFMAN J,HARRIS D L,et al.Erysipelothrix rhusiopathiae: Genetic charact erization of Midwest US isolates and live commercial vaccines using pulsed field gel electrophoresis[J].J Vet Diagn Invest,2004,16(2) : 101-107.
[27]JIN F L,XU X X,YU X Q,et al.High-level expression of active recombinant ubiquitin carboxyl terminal hydrolase of drosophila melanogaster in Pichia pastoris[J].Protein Expr Purif,2009,65(2) : 115-121.
[28]杨梅,温真,林丽玉.毕赤酵母蛋白表达系统研究进展[J].生物技术通报,2011(4) : 46-51.
[29]DALY R,HEARN M T W.Expression of heterologous proteins in Pichia pastoris: A useful experimental tool in protein engineering and production[J].J Mol Recognit,2005,18(2) : 119-138.
【责任编辑柴焰】
Cloning surface protective antigen A gene from Erysipelothrix rhusiopathiae and its expression in Pichia pastoris
JIANG Zhiqiong,ZHONG Zemin,TAN Bomin,YU Xiyao,HUANG Yumao
(College of Veterinary Medicine,South China Agricultural University,Guangzhou 510642,China)
Abstract:【Objective】Immunization protection area protein,which lies in the N-terminal protective domain of surface protective antigen A(SpaA) of Erysipelothrix rhusiopathiae,was expressed in Pichia pastoris X-33.【Method】Using the swine erysipelas,which was isolated from a pig farm in Guangdong as template,a pair of primers was designed according to the cDNA sequence of Spa gene from NCBI.The amino terminal sequence was achieved by PCR amplification,after being inserted into the expression vector pPICZαC to get the recombinant plasmid of pPICZαC-SpaA-N.Recombinant plasmid of pPICZαCSpaA-N by Sac I enzyme was linearized,and electro transformed it into P.pastoris X-33.The positive transformant,screened by YPDS tablet with ZencinTMand identified by PCR at different concentrations of ZencinTM,achieved the high number of copies which were induced and cultured by methanol.Supernatant was collected by centrifugation at 48,72,96 h respectively after induction,and SDS-PAGE and Western-blot tests were carried out.【Result and conclusion】The SpaA gene was successfully cloned and expressed,and the recombinant plasmid of pPICZαC-SpaA-N was constructed.The immunization protection area protein,which lies in SpaA gene amino terminal of swine E.rhusiopathiae in P.pastoris X-33,was successfully expressed.SpaA gene of swine E.rhusiopathiae has been successfully expressed asim-book=21,ebook=25munization protection area protein in yeast host,which will lay a foundation for the development and the mass production of subunit vaccine of swine E.rhusiopathiae.
Key words:Erysipelothrix rhusiopathiae; SpaA gene; Pichia pastoris; cloning and expression; subunit vaccine
作者简介:蒋志琼(1987—),女,硕士研究生,E-mail: 827209378@ qq.com;通信作者:黄毓茂(1964—),男,副教授,博士,E-mail: ymaohuang@ scau.edu.cn
收稿日期:2014-04-21
文章编号:1001-411X(2015) 03-0020-06
文献标志码:A
中图分类号:S855
