野生型和栽培型马齿苋的种源鉴定及生物学特性与生态适应性比较
2016-01-15杜建梅云晶晶杨颜颜张利娟胡卫成陆长梅
杜建梅, 云晶晶, 杨颜颜, 张利娟, 胡卫成, 陆长梅
(1.南京师范大学生命科学学院, 江苏 南京210023;2.江苏省环洪泽湖生态农业生物技术重点实验室(淮阴师范学院), 江苏 淮安223300)
根据《中国植物志》记载,马齿苋(Portulaca oleracea L.),别名半支莲、松叶牡丹等,属马齿苋科马齿苋属一年生肉质药食兼用草本植物。它不仅具有丰富的营养价值,还具有消炎止痛、降血脂、降血压的特殊功效。近年来发现的马齿苋体内高含量的α-亚麻酸[1]以及褪黑激素[2]等特殊保健药用成分更显示其巨大的药用保健价值。此外,作为C 4兼性CAM植物,马齿苋分布广泛,并对高温、干旱、重金属污染等逆境的耐受能力极强[3],显示其巨大的生态修复前景。目前国内外大量研究正围绕这2个方向展开。但马齿苋种源多样,对于在不同情形、不同目的下如何选择不同的马齿苋种源,目前尚未见文献涉及。
我国现有马齿苋资源一般分为2种类型,野生型和栽培型[4]。栽培型马齿苋,系欧洲的地产种,从荷兰引进(商品名:荷兰马齿苋),并于1995年在河南首次试种成功。两者除了在株型上有较大的差异外[4],在种源、生物学特性以及营养价值、抗逆性等方面的差异以及各自的应用方向目前尚少见文献涉及。
鉴于上述现状,本研究以南京地产野生型马齿苋和栽培型荷兰马齿苋的种子为材料,比较两者的种子特征、萌发特性和植株生长发育特性,并从种子特征、萌发特性以及植株生物学特性等角度初步分析两者在生态修复和农业生产上的应用前景,从而为发展马齿苋积累基础资料。
1 材料与方法
1.1 材 料
供试野生型马齿苋种子于2012年9月下旬采自南京师范大学仙林校区植物园(118°95′E,32°15′N);栽培型马齿苋种子购于北京京研益农种苗公司。
1.2 方 法
1.2.1 种子宏观形态特征
千粒重采用百粒法测定[5]。种子大小采用扫描仪法测定[6]。
1.2.2 种皮微观形态
选择饱满种子,超声清洗100s后,同向粘贴于扫描电镜样品台的双面胶上,真空喷镀金膜320s,并于扫描电镜下(JSM-5610LV,日本电子公司,日本),观察种皮微形态。
1.2.3 种子萌发
采用培养皿滤纸法,于30℃、12h/12h(L/D)光周期下进行种子萌发[7]。记录初始萌发时间、萌发结束时间、萌发率等。
1.2.4 植株培养与观测
野生型种子在35℃水中浸种12h后,与栽培型种子同时播种于已充分吸胀的泥炭块(jiffy公司,加拿大)表面,于 Adaptis A-1000型生物培养箱(CONVIRON,加拿大)中,14h/10h(L/D)、32℃/28℃(L/D)、RH 70%、光强500μmol/(m·s)条件下萌发培养。隔天浇水1次,第6天开始每周在浇水时施加1‰全营养型速溶肥(花多多1号,美国Scotts公司)1次。每天记录种子萌发及植株生长发育状况。用直尺测量主茎高度,游标卡尺测量茎粗、叶长、叶宽和花直径等指标。每个指标重复10次。每个蒴果内种子数以随机取30个蒴果统计。
1.3 统计方法
结果以平均值显示。数据处理与分析用Microsoft office Excel 2007和 Spss 18.0 软件完成。图表中“*”和“**”分别表示2种类型马齿苋存在显著(p<0.05)与极显著差异(p<0.01)。
2 结果与分析
2.1 种子形态特征比较
肉眼观察,2种类型的马齿苋种子均呈近圆形,在自然光下呈黑色或黑褐色,近于亚光。两者均属小种子,野生型的更小,肉眼可明显区分。表1中数据显示,野生型种子的千粒重仅为栽培型的1/6、长度和宽度约为栽培型的1/2。

表1 野生型与栽培型马齿苋种子大小比较
扫描电镜下正面观察(图1-A 1、A 2),两者均呈宽倒卵肾形,在合点处凹陷形成脐部,并在脐部有蝶翅形脐膜覆盖;脐膜疏松多孔,呈丝瓜络状(图1-C 1、C 2);栽培型种子的脐膜相对较大(图1-A 1、A 2,C 1、C 2)。从侧面观察(图1-B 1、B 2),2种子均呈厚圆饼状,脐部稍尖,脐部对侧较为钝圆。两者的种皮表面均有纹饰,野生型扁平花样状纹饰排列整齐(图1-A 1、C 1、D 1),栽培型的纹饰不仅不规则,且纹饰上还有大小不一、排列不规则的瘤状突起(图1-A 2、C 2、D 2)。在种子侧面两者均具瘤状突起,但野生型的突起大小更均一,排列更整齐(图1-B 1、B 2)。
2.2 种子萌发特征比较
表2显示,在未经任何预处理情形下,栽培型马齿苋种子在6h时开始萌发,1d内萌发结束,萌发率高达95%;而野生型马齿苋种子在24h后开始萌发,虽然最终萌发率可达85.7%,但萌发时间需要持续8~10d。与多数常见植物种子的萌发速度相比,马齿苋种子的萌发方式均属于爆发型,但栽培型种子萌发更早、萌发迅速整齐[8]。

表2 野生型和栽培型马齿苋种子的萌发特征比较
2.3 植株生物学特性比较
按照材料与方法中的植株培养与观察方法,播种、培养2种类型马齿苋,并观察植株生长发育全过程,具体结果见表3。栽培型种子虽未经预处理,但萌发速度和整齐度仍显著高于野生型,子叶和真叶抽出时间更早。两者茎均为肉质圆柱状,光滑无毛,节间短。当野生型马齿苋主茎长至约0.5cm时,腋芽先后从子叶和第1对真叶叶腋内长出,形成4个一级侧枝。虽然后续仍有侧枝长出,但基部4个一级侧枝长势最旺,并逐渐与主茎相当,形成以茎基部为中心向四周辐射状的野生型马齿苋株形。栽培型马齿苋茎直立生长,主茎基部最多出现2条侧枝,且主侧枝区分明显。两者叶序均为互生,少近对生,均为肥厚马齿状,无叶柄。栽培型的叶长和叶宽均在野生型的2倍以上。约20d后,两者生长速度明显加快,栽培型主茎伸长速度是野生型的2倍(表3)。分别约在30d和40d时,茎顶端出现花苞,主茎伸长减缓至停止,栽培型主茎的最终长度约为野生型的2倍(表3)。
野生型的主茎和4个基部一级侧枝伸长停止后,各自从基部产生次级侧枝并次第形成花苞,如此以往,最终形成3~4级侧枝。栽培型则在主茎出现花苞后,部分植株在主茎顶端叶腋内抽出次级侧枝,并逐渐出现花苞,栽培型马齿苋多见二级侧枝。在花苞出现10d后,约在09:00时花苞开始开放,10:00时完全展开,11:00时开始闭合,12:00时完全闭合。
两萼片基部为半圆形合生,相对抱合成一个花苞,花苞无梗。栽培型花苞大于野生型,但单株花苞数(10~30个)远少于野生型(约300个)。花黄色,两性,花程式均为 K(2)C 5A10-12G(4-6∶1∶∞)[9]。栽培型花直径约为野生型的2倍(表3)。两者花苞开放或不开放均可完成受精作用。受精7~8d后,种子成熟,蒴果盖裂,种子散出。野生型马齿苋的花苞总数以及每一蒴果内种子数高于栽培型,致使野生型马齿苋种子量远大于栽培型。
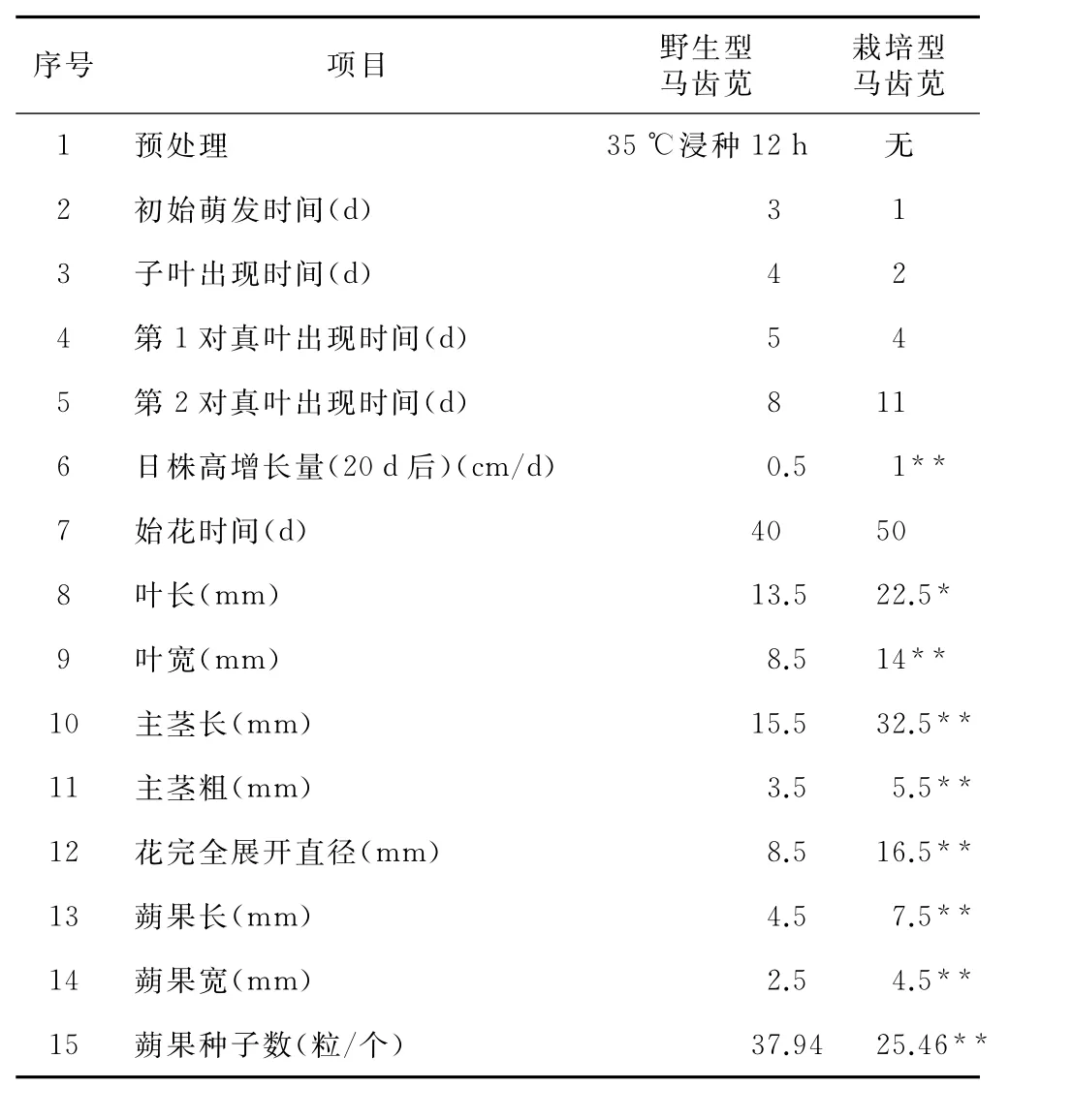
表3 野生型和栽培型马齿苋的部分生物学特性比较
3 讨 论
3.1 种源鉴定

图1 野生型和栽培型马齿苋种子形态比较
种子发育全过程都是在比较封闭的子房内进行,其形态及其表面细微结构受环境影响小而相对稳定,有着重要的分类学价值。Danin等[10-13]依据种子大小和种皮微结构将苏丹、地中海等多个地区的马齿苋分为1 9个亚种。对比Danin等的马齿苋分类检索表[13],并经Danin确认,将本实验涉及的野生型马齿苋归属到 P.granulatostellulata(Poelln.)C.Ricceri et P.V.Arrigoni亚种,栽培型马齿苋归属到 P.edulis Danin &Bagella亚种。P.edulis为天然6N,染色体总数为54;P.granulatostellulata为天然4N,染色体总数为36[13]。
表2和表3中的结果显示,相比野生型马齿苋,栽培型马齿苋种子萌发迅速,株型高大,生长旺盛,花部器官大,但有性生殖能力相对较弱(蒴果和蒴果内种子数少)。这与一般植物体随着染色体倍数和基因拷贝数的增加,株型增大、有性生殖能力减弱等特性相一致[14]。是否栽培型马齿苋比野生型马齿苋的抗性更强,次生代谢物质含量更丰富还有待进一步探讨[14]。栽培型马齿苋的这种萌发迅速整齐、株型直立高大的特点有利于田间迅速建群与采摘,提示其在农业栽培上的应用前景可能更大。
3.2 生态适应性
种子大小和形态特征直接影响种子的传播、散布和存活几率。Westoby等[15]和 Thompson等[16]的研究表明,0.1~100mg重量范围的种子可以通过风雨、附着、蚂蚁以及脊椎动物等传播散布方式,而小于0.1 mg的种子趋向于没有媒介。本研究涉及的2种类型的马齿苋种子均为小种子(表1),种子表面没有如芒、刺、绒毛等附属物(图1),整体附着能力较差、随风飘散能力也较弱;种子成熟后,种子多以蒴果盖裂方式散落于母体周围。这表明,两者种子的空间扩散范围均很有限,质量和体积较大的栽培型马齿苋(0.4±0.001)mg种子的空间扩散范围可能会由于有蚂蚁等小动物参与而稍大。
越细小的马齿苋种子越容易通过土壤缝隙陷入土壤深处形成种子库[16]。深埋于地下20~40年的马齿苋种子仍能正常萌发[17]的现象提示,在极端环境来临时,土壤种子库可为马齿苋提供良好的缓冲场所;在极端环境消失后,种子一旦到达土壤表层,就会成为占据土表的先锋植物。这不仅极大地降低了种群灭亡几率,同时也为其成为生态修复备选材料提供了可能。这可能也是马齿苋资源广布全球温带和热带,种群数量位居第八的重要原因之一。
栽培型马齿苋种子表面密集的瘤状突起(图1),增加了种子表面积,便于种子对水分的快速吸收和快速萌发(表2和表3)。这种萌发对策,在遭遇极端环境时,可能会导致植物区域性灭绝。而野生型种子较为光滑的表面(图1)、相对较慢和较不整齐的萌发策略(表2),致使种子即使在最适环境下也不会一次性萌发,这属于一种可避免种群灭绝的、谨慎的萌发策略[18]。这也提示,野生型马齿苋可能在生态修复方面具有更大的应用前景。
2种类型马齿苋在花苞完全、不完全、甚至不开放时,都可以完成授粉授精,产生正常种子[9]。马齿苋的这种闭花传粉受精方式,可以让马齿苋在环境不适于开花传粉时依然完成生殖过程,同时还可使花粉免遭雨水淋湿和昆虫吞食,从而提高马齿苋对逆境的适应能力。这可能也是马齿苋,尤其是种子产生量很大的野生型马齿苋,分布广泛的重要原因之一。
总之,2种类型马齿苋在倍性、种子大小、萌发行为以及生物学特性方面的差异,带来它们的生态适应性差异,并提示它们的应用方向也可能存在一定差异。小种子的特性决定了在播种时,需要减少土壤缝隙存在,以免大量种子陷入土壤深处,无法萌发;如果在常规土壤栽培供菜用或饲料用,可以选择栽培型马齿苋,以充分利用其种子萌发迅速整齐、植株直立便于采摘的特性;如果在干旱等极端土壤播种供生态修复用,则可以选择野生型马齿苋,主要利用其种子量大,不需年年播种、萌发谨慎、种群不易被灭绝等特性。
[1]Simopoulos AP.Salem Jr N:Purslane:a terrestrial source of w-3 fatty acids[J].New England Journal of Medicine,1986,315:833.
[2]Simopoulos AP,Tan DX,Manchester LC,et al.Purslane:a plant source of omega-3fatty acids and melatonin[J].Journal of Pineal Research,2005,39:331-332.
[3]Miller TE,Aing JS,Huete AR.The agricultural potential of selected C 4plants in arid environments[J].J Arid Environ.1984(7):275-285.
[4]戚跃科,陈策,陆燕宾,等.野生和栽培马齿苋黄酮含量的比较研究[J].丽水学院学报,2009,31(2):40-42.
[5]国际种子检验协会(ISTA)编;农业部全国农作物种子质量监督检测中心,浙江大学种子科学中心译.1996国际种子检验规程[M].北京:中国农业出版社,1999.
[6]刘长江,林祁,贺建秀.中国植物种子形态学研究方法和术语[J].西北植物学报,2004,24(1):178-188.
[7]杨子仪,徐亚莉,葛峰,等.兽药洛克沙胂处理对空心菜生长代谢和营养价值的影响[J].地球与环境,2013,40(4):441-450.
[8]王桔红,崔现亮,陈学林,等.中、旱生植物萌发特性及其与种子大小关系的比较[J].植物生态学报,2007,31(6):1 037-1 045.
[9]陈国元,朱旭东,陈素娟.野生马齿苋生物学特性调查[J].中国野生植物资源,2012,31(5):61-63.
[10]Danin A,Bagella S.A new cultivar microspecies of Portulaca oleracea aggregate from the East Mediterranean[J].Willdenpwia,2012,42(1):63-65.
[11]Danin A,Reyes-Betancort JA.The status of Portulaca oleracea L.inTenerife[J].The Canary Islands,Lagascalia.2006,26:71-81.
[12]Danin A,Domina G,Raimondo FM.Microspecies of the Portulaca oleracea aggregate found on major Mediterranean islands (Sicily,Cyprus,Crete,Rhodes)[M].Flora Mediterranea,2008,18:89-108.
[13]Danin A,Raus T.A key to 19microspecies of the Portulaca oleracea aggregate.In Plant stories,Chapter G:Portulaca oleracea-a Complex Full of Riddles[M].Flora of Israel Online,http://flora.org.il/en/books/plant-stories-2/chapter-k/useful_plants_k2/.
[14]肖勇,杨耀东,夏薇,等.多倍体在植物进化中的意义[J].广东农业科学,2013(16):127-130.
[15]Westoby M,Jurado E,Leishman M.Comparative evolutionary ecology of seed size[J].Tree,1992,7(11):368-372.
[16]Thompson K,Band SR,Hodgson JG.Seed size and shape predict persistence in soil[J].Functional Ecology,1993,7:236-241.
[17]李善林,刘德荣,韩烈刚.草坪杂草[M].北京:中国林业出版社,1999:6.
[18]Gutterman Y.Environmental factors and survival strategies of annual plant species in the Negev Desert,Israel[J].Plant Species Biology.2000,15:113-125.
