Zebularine 对肝癌HepG2 细胞凋亡的影响及其机制
2015-12-31丁佑铭於展飞廖晓峰
丁佑铭,於展飞,廖晓峰
1.武汉大学人民医院肝胆腹腔镜外科,湖北 武汉430060;2.襄阳市中心医院普外科
原发性肝癌(hepatocellular carcinoma,HCC)是高度恶性肿瘤,尽管其发病率在恶性肿瘤中仅排第5 位,但其死亡率高居第2 位。全球每年新增约75 万病例,并有超过70 万患者死亡[1]。研究发现,表观遗传学在肿瘤发生、发展及转归过程中扮演重要角色,其中DNA 甲基化是基因表达的遗传学改变的主要方式之一[2]。研究发现,抑癌基因失活机制之一是启动子CpG 高甲基化,因此,去甲基化药物作为新的抑制细胞增殖和诱导细胞分化的肿瘤治疗方法具有广阔的研究前景。Zebularine[1-(B-D-呋喃核糖苷)-1,2-二氢嘧啶-2-酮]作为新型DNA 甲基转移酶(DNA methyltransferase,DNMT)抑制剂,可抑制后者甲基化作用[3]。人类相关转录因子3(human runt-related transcription factor 3,RUNX3)基因在肝癌、乳腺癌、胃癌、大肠癌及肺癌等多种肿瘤组织中表达下调甚至缺失,在肿瘤的发生、发展中起着重要作用[4-5],研究发现,RUNX3 基因表达下调或缺失的主要机制是其基因启动子甲基化[6-7]。本研究以人肝癌细胞系HepG2 细胞作为研究对象,观察Zebularine 对肝癌细胞凋亡的影响,检测其对细胞凋亡相关蛋白Bcl-2、Bax、Caspase3、Caspase9 表达的影响,并检测其能否上调RUNX3 基因表达。
1 材料与方法
1.1 材料 人肝癌细胞系HepG2 细胞购自中国科学院上海细胞库,由武汉大学中心实验室冻存;Zebularine、MTT、DMSO、罗 丹 明 123 (Rhodamine 123,Rho123)购自Sigma 公司;胎牛血清购自Invitrogen 公司;RPMI 1640 培养液购自杭州四季青生物材料研究所;AnnexinV/PI 试剂盒购自BENDER 公司;Bcl-2、Bax、Pro-Caspase-3、Pro-Caspase-9、Cleaved-Caspase-3、Cleaved-Caspase-9 及RUNX3 单克隆抗体购自Cell Signaling Technology 公司;辣根酶标记兔抗山羊IgG 或兔抗山羊IgG 购自武汉博士德生物科技有限公司。
1.2 方法
1.2.1 细胞培养:HepG2 细胞培养于RPMI 1640 培养液中(含10%小牛血清,100 U/ml 青霉素,100 μg/ml链霉素),置于37 ℃,饱和湿度,5% CO2培养箱内培养,每次均取生长状态良好的并处于对数期的细胞进行实验。
1.2.2 MTT 法检测细胞增殖抑制率:取对数生长期的细胞配制成1.5 ×104个/ml 的细胞混悬液,将混悬液按每孔100 μl 加入96 孔培养板,并在周围孔内加入高压消毒的PBS 溶液,在孵箱中孵育培养约24 h 后取出,弃上清,换Zebularine 终浓度为0、100、200、400 μmol/L 的培养液,每组设置5 个复孔,同时设立调零孔,继续培养24、48、72 h,于每次实验结束前4 h 弃孔内液体,每孔加入20 μl MTT 溶液(浓度为0.5 mg/ml),培养4 h,弃孔内液,加150 μl DMSO,置于摇床上振荡10 min 后在酶标仪上检测490 nm 处的吸光值(A值),抑制率(%)=[1 -(实验组平均A 值-调零孔A值)/(对照组平均A 值-调零孔A 值)]×100%。
1.2.3 流式细胞仪检测细胞凋亡:将对数生长期的细胞配制成1 ×106个/ml 的细胞混悬液,将悬液接种于6 孔板中,在孵箱中孵育培养约24 h 后取出,弃上清。换含Zebularine 终浓度为0、100、200、400 μmol/L 的培养液,继续培养48 h 后,收集细胞,预冷PBS 洗涤细胞2 次,并将细胞重悬于Binding Buffer 中。加入5 μl Annexin V-FITC 室温、避光反应10 min,加入5 μl PI混匀,室温、避光反应15 min。筛网过滤后,上机检测细胞凋亡率。
1.2.4 Rho123 染色流式细胞仪检测细胞线粒体膜电位水平:细胞接种及分组同上,培养48 h 后,收集细胞,PBS 漂洗3 次,将细胞重悬于1 ml 的Rho123(1 μg/ml)中,放置培养箱中继续孵育30 min。PBS 洗涤3 次,用EPICS-XL 型流式细胞仪以507 nm 为激发波长,529 nm 为发射波长测定细胞内荧光强度。
1.2. 5 Western blotting 检测Bcl-2、Bax、Caspase-3、Caspase-9 及RUNX3 蛋白表达变化:细胞接种及分组同上,培养48 h 后,收集细胞,加入100 μl 裂解液+1.742 μl PMSF 蛋白酶抑制剂重悬细胞,用移液器吹打细胞置混匀,使裂解液和细胞充分接触。置于冰上裂解约45 min。后12 000 r/min(离心半行5 cm),4℃离心30 min,后吸取上清,将样品分装至预冷的EP管中,-80 ℃保存。BCA 法测定蛋白质浓度,加入1/5体积的5 ×上样缓冲液至提取的细胞蛋白样品中,沸水煮沸5 min,-20 ℃保存。将细胞蛋白样品行SDS-聚丙烯酰胺凝胶电泳(上样量20 μg/孔),电泳结束后转膜至PVDF 膜,再加入5%脱脂奶粉,室温封闭1 h,分别加入1 ∶1 000 稀释的兔抗或鼠抗人Bcl-2、Bax、Caspase-3、Caspase-9 及RUNX3 抗体,4 ℃过夜,β-actin作为内参,第2 天TBST 洗膜后,根据一抗来源不同分别加入1∶1 000 稀释的辣根酶标记的山羊抗或山羊抗鼠二抗,室温放置2 h,再次TBST 洗膜后加入ECL 发光液显色,暗室内显影并观察各条带深浅变化。
1.3 统计学处理 采用SPSS 17.0 统计软件进行分析,计量资料以s 表示,两样本间参数比较采用独立样本t 检验,P <0.05 为差异有统计学意义。
2 结果
2.1 Zebularine 对HepG2 细胞增殖的影响 MTT 法检测显示,Zebularine 能显著抑制HepG2 细胞增殖,Zebularine 对HepG2 细胞的增殖抑制作用呈时间和浓度依赖性,药物浓度越高、作用时间越长,其发挥抗肿瘤作用效果也越好(见图1)。
2.2 Zebularine 对HepG2 细胞凋亡的影响 HepG2细胞经Zebularine 处理48 h 后,流式细胞仪凋亡检测结果显示,Zebularine 组的细胞凋亡率从12.4%增加35.4%,而凋亡率仅为3.3%(见图2)。
2.3 Zebularine 对HepG2 细胞凋亡相关蛋白Bcl-2、Bax、Caspase-3、Caspase-9 表达的影响 Zebularine 处理HepG2 细胞48 h 后结果显示,随着Zebularine 浓度的升高,Bax 蛋白表达量也逐渐升高,而Bcl-2 蛋白表达量逐渐降低,Pro-Caspase-3、Pro-Caspase-9 表达减弱,而Cleaved-Caspase-3、Cleaved-Caspase-9 表达增强,呈浓度依赖性(见图3)。
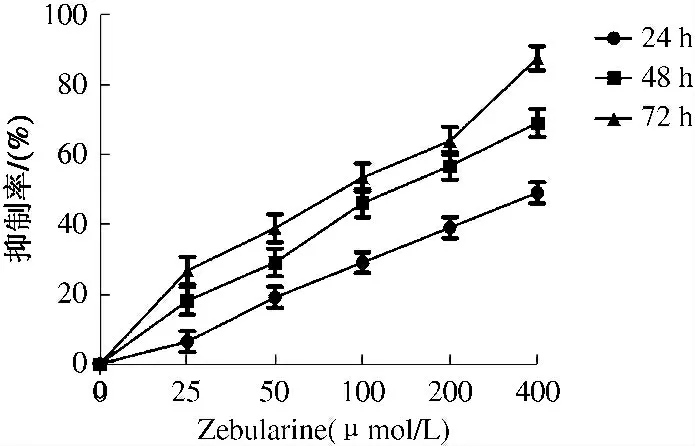
图1 Zebularine 对HepG2 细胞抑制率的影响Fig 1 Effect of Zebularin on cell inhibition ratio in HepG2 cells
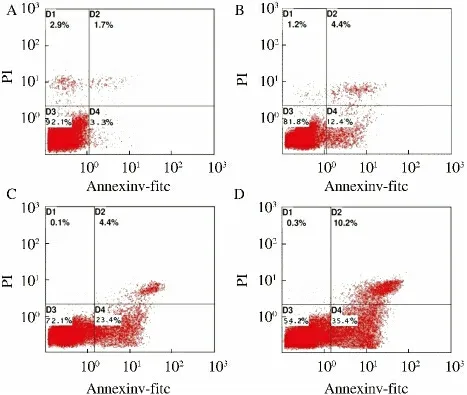
图2 Zebularine 对HepG2 细胞凋亡率的影响 A:0 μmol/L;B:100 μmol/L;C:200 μmol/L;D:400 μmol/LFig 2 Effect of Zebularine on cell apoptosis in HepG2 cells A:0 μmol/L;B:100 μmol/L;C:200 μmol/L;D:400 μmol/L

图3 Zebularine 对HepG2 细胞凋亡相关蛋白Bcl-2、Bax、Caspase-3、Caspase-9 表达的影响Fig 3 Effect of Zebularine on the expression of Bcl-2,Bax,Caspase-3 and Caspase-9 in HepG2 cells
2.4 Zebularine 对HepG2 细胞线粒体膜电位的影响Zebularine(0、100、200、400 μmol/L)处理48 h 后,HepG2 细胞线粒体荧光强度随着药物浓度的升高逐渐降低(P <0.05)。
2.5 Zebularine 对HepG2 细胞RUNX-3 蛋白表达的影响 Zebularine 处理HepG2 细胞48 h 后,Western blotting 检测结果显示,随着药物浓度的逐渐升高,抑癌基因RUNX3 表达逐渐增加(见图4)。

图4 Zebularine 对HepG2 细胞RUNX3 表达的影响Fig 4 Effect of Zebularine on the expression of RUNX3 in HepG2 cells
3 讨论
国内外研究发现,转录生长因子β(transforming growth factor,TGF-β)信号通路在肿瘤的发生、发展过程中具有重要意义[8-9]。RUNX3 参与了TGF-β 信号通路诱导生长抑制的过程[10]。研究证实,RUNX3 基因在多种肿瘤如乳腺癌、胃癌、大肠癌、肝癌及肺癌等肿瘤中表达下调、缺失,导致TGF-β 信号途径紊乱,引起TGF-β 信号失活,进而引发细胞凋亡障碍,导致肿瘤的发生,并在肿瘤发展过程中起重要作用[4-5]。CpG区异常高甲基化是除突变和缺失外肿瘤抑癌基因表达下调或沉默的机制,继而导致转录停止,RUNX3 失活,与细胞增殖、凋亡、分化、恶性转化密切相关[11]。有证据表明,恢复RUNX3 基因的表达能抑制细胞增殖、诱导细胞凋亡、调节细胞周期并抑制细胞侵袭转移[12]。
由于DNA 甲基化在肿瘤发生、发展和转归过程中扮演重要角色,且其甲基化改变是可逆的,因此,DNA甲基化可能成为药物治疗的理想靶点。Zebularine 是新型DNA 甲基转移酶(DNA methyltransferase,DNMT)抑制剂,能抑制DNMT 甲基化作用。研究发现,Zebularine 能使卵巢癌细胞中因启动子CpG 高甲基化而表达低下或沉默的RASSF1A 及Hmlh1 基因表达上调[13-14]。本实验发现,Zebularine 处理HepG2 细胞48 h 后,RUNX3 蛋白表达上调,且呈浓度依赖性。MTT法检测结果显示,Zebularine 对HepG2 细胞的增殖抑制作用呈时间和浓度依赖性,药物浓度越高、作用时间越长,其发挥抗肿瘤作用效果越好。流式细胞仪检测Zebularine 处理HepG2 细胞后可检测到明显凋亡,提示Zebularine 抑制HepG2 细胞增殖并诱导其凋亡的作用与其上调RUNX3 表达可能有关。
本研究进一步探讨了Zebularine 对细胞凋亡线粒体途径的影响,Zebularine 处理48 h 后,Western blotting 检测结果显示HepG2 细胞的Bcl-2 表达下调,Bax表达上调,Pro-Caspase-3、Pro-Caspase-9 表达减弱,而Cleaved-Caspase-3、Cleaved-Caspase-9 表达增强,且呈剂量依赖性。此外,Zebularine 能降低HepG2 细胞线粒体膜电位,表明Zebularine 可通过线粒体途径促进细胞凋亡。Bcl-2 是一种内膜蛋白,是公认的强效凋亡抑制分子,其主要是通过增强线粒体膜电位、保持线粒体内外膜的完整性、抑制钙离子释放、阻止核酸内切酶活化等机制发挥凋亡抑制作用[15-16];Bax 也属于Bcl-2家族成员,但其与Bcl-2 作用相反,Bax 可直接激活死亡效应因子Caspase 或改变细胞膜通透性引起细胞色素C 释放某些离子和小分子通过细胞膜,进而促进细胞凋亡。Caspase 是引起凋亡的直接效应物。Caspase-9 是线粒体凋亡途径的关键蛋白酶,处于Caspase“瀑布式”激活的顶端,其活化对线粒体凋亡通路具有重要意义,并进一步激活细胞凋亡的执行器Caspase-3,后者活化后可通过裂解DNA 修复相关分子、凋亡抑制蛋白、细胞外基质蛋白和骨架蛋白等多种机制,导致细胞凋亡。
综上所述,本研究认为,在肝癌细胞中,抑癌基因RUNX3 由于启动子甲基化而失活,Zebularine 能逆转RUNX3 甲基化状态,恢复RUNX3 表达,从而抑制细胞增殖,并通过细胞凋亡线粒体途径促进细胞凋亡。
[1] Jemal A,Bray F,Center MM,et al. Global cancer statistics[J]. CA Cancer J Clin,2011,61(2):69-90.
[2] Shenker N,Flanagan JM. Intragenic DNA methylation:implications of this epigenetic mechanism for cancer research [J]. Br J Cancer,2012,106(2):248-253.
[3] Yoo CB,Cheng JC,Jones PA. Zebularine:a new drug for epigenetic therapy[J]. Biochem Soc Trans,2004,32(Pt 6):910-912.
[4] Jeong P,Min BD,Ha YS,et al. RUNX3 methylation in normal surrounding urothelium of patients with non-muscle-invasive bladder cancer:potential role in the prediction of tumor progression[J]. Eur J Surg Oncol,2012,38(11):1095-1100.
[5] Shiraha H,Nishina S,Yamamoto K. Loss of runt-related transcription factor 3 causes development and progression of hepatocellular carcinoma[J].J Cell Biochem,2011,112(3):745-749.
[6] Lee CW,Ito K,Ito Y. Role of RUNX3 in bone morphogenetic protein signaling in colorectal cancer[J]. Cancer Res,2010,70(10):4243-4252.
[7] Kodach LL,Jacobs RJ,Heijmans J,et al. The role of EZH2 and DNA methylation in the silencing of the tumor suppressor RUNX3 in colorectal cancer[J]. Carcinogenesis,2010,31(9):1567-1575.
[8] Calone I,Souchelnytskyi S. Inhibition of TGFβ signaling and its implications in anticancer treatments [J]. Exp Oncol,2012,34(1):9-16.
[9] Liu LC,Tsao TC,Hsu SR,et al. EGCG inhibits transforming growth factor-β-mediated epithelial-to-mesenchymal transition via the inhibition of Smad2 and Erk1/2 signaling pathways in nonsmall cell lung cancer cells[J]. J Agric Food Chem,2012,60(39):9863-9873.
[10] Watanabe K,Sugai M,Nambu Y,et al. Requirement for Runx proteins in IgA class switching acting downstream of TGF-beta 1 and retinoic acid signaling[J]. J Immunol,2010,184(6):2785-2792.
[11] Ku JL,Kang SB,Shin YK,et al. Promoter hypermethylation downregulates RUNX3 gene expression in colorectal cancer cell lines[J].Oncogene,2004,23(40):6736-6742.
[12] Chi XZ,Yang JO,Lee KY,et al. RUNX3 suppresses gastric epithelial cell growth by inducing p21 (WAF1/Cip1)expression in cooperation with transforming growth factor {beta}-activated SMAD [J].Mol Cell Biol,2005,25(18):8097-8107.
[13] Shen WJ,Liu SL,Qiu GR. Zebularine regulates reexpression of RASSF1A gene in human ovarian cancer cell A2780[J]. J Southeast Univ (Med Sci Edi),2011,30(4):550-554.沈文静,刘石磊,邱广蓉. Zebularine 调控卵巢癌A2780 细胞RASSF1A 基因再表达的研究[J]. 东南大学学报(医学版),2011,30(4):550-554.
[14] Liu SL,Shen WJ,Qiu GR. Zebularine induces demethylation and reexpression of hMLH1 Gene in human ovarian cancer cell line[J].Journal of China Medical University,2011,40(7):597-599,604.刘石磊,沈文静,邱广蓉. Zebularine 调控卵巢癌细胞hMLH1 基因再表达[J]. 中国医科大学学报,2011,40(7):597-599,604.
[15] Gogada R,Prabhu V,Amadori M,et al. Resveratrol induces p53-independent,X-linked inhibitor of apoptosis protein (XIAP)-mediated Bax protein oligomerization on mitochondria to initiate cytochrome c release and caspase activation[J]. J Biol Chem,2011,286(33):28749-28760.
[16] Pucci B,Bertani F,Karpinich NO,et al. Detailing the role of Bax translocation,cytochrome c release,and perinuclear clustering of the mitochondria in the killing of HeLa cells by TNF[J]. J Cell Physiol,2008,217(2):442-449.
