百山祖木荷的种群结构与分布格局
2015-12-30陈德良顾莎莎丁炳扬骆争荣
陈德良,顾莎莎,丁炳扬,骆争荣
(1. 浙江凤阳山——百山祖国家级自然保护区百山祖管理处,浙江 庆元 323800;2. 温州大学 生命与环境科学学院,浙江 温州 325035;3. 丽水学院 生态学院,浙江 丽水 323000)
百山祖木荷的种群结构与分布格局
陈德良1,顾莎莎2,丁炳扬2,骆争荣3
(1. 浙江凤阳山——百山祖国家级自然保护区百山祖管理处,浙江 庆元 323800;
2. 温州大学 生命与环境科学学院,浙江 温州 325035;3. 丽水学院 生态学院,浙江 丽水 323000)
通过调查百山祖北坡250 m×200 m样地内木荷分布和胸径/基径情况,分析种群的径级结构,并采用方差均值比(Cx)、负二项参数(K)、丛生指数(I)平均拥挤度(m*)与聚块性指标(C)5个分布格局指标评估比较该常绿阔叶林中木荷不同生长阶段的分布格局。结果表明:木荷种群径级分布呈现双峰型,幼苗和中等大小的树木较丰富,种群更新基本良好;从整个种群和不同发育阶段来看,木荷的分布格局都呈聚集分布,但随年龄增长,聚集度逐渐降低;种子扩散限制、生境异质性是形成木荷聚集性分布格局的主要原因;由于种内竞争和密度制约效应使木荷的聚集度随着植物的生长不断下降。
聚集;双峰型;更新良好;密度制约;竞争
森林群落中的植物种群是构成群落的基本单位,在个体、群落和生态系统等层次中起到了桥梁的的作用,是生态学研究最基本的层次。种群的年龄结构和分布格局是种群生态学的核心研究内容[1]。分布格局是研究种群个体在空间上的配置情况,是种群自身特性、环境条件等因素综合作用的结果[2]。同时,物种的分布格局对物种的生长、繁殖、死亡、再生、资源利用以及林窗的形成等具有显著的影响[3]。而年龄结构则是指植物群落中不同年龄层次个体的分配情况,由不同年龄段个体死亡率和增补率决定的,在一定程度上可以反映种群的动态发展趋势[4]。不同的植物种群在群落空间中均有相对稳定的配置状况,分析种群的地位和作用,可以揭示种群分布格局形成的原因,阐述种群及群落的动态变化[5],所以研究种群结构和分布格局对进一步认识群落中物种的生态过程、了解植物群落和环境因子之间相互作用的关系有着重要的意义。
木荷(Schima superba)为山茶科木荷属乔木,终年常绿,广泛分布于亚洲热带和亚热带,是我国亚热带东部常绿阔叶林的主要优势种之一[6]。优势种对群落的结构塑造和群落环境的形成有明显的控制作用, 既是群落骨架, 又决定了群落外貌和群落类型[7]。木荷作为亚热带常绿阔叶林的优势种,在群落演替过程中的动态规律, 学术界已讨论多年[4,6, 8~9]。但较少有研究结合空间分布和径级结构共同探讨木荷在常绿阔叶林中的作用和动态。本研究分析木荷的时空格局,不仅可以使我们更深入的了解木荷在常绿阔叶林中的动态,也有助于我们认识群落结构的形成与维持机制。
1 研究地概况
本研究在“浙江凤阳山——百山祖国家级自然保护区”位于庆元县境内的百山祖部分(27° 37′ 45″ ~ 27° 50′30″ N,119° 7′ 45″ ~ 119° 19′ 20″ E)进行。植物种类丰富,多样性高,植被组成有明显的垂直带谱系列。区内以侵蚀地貌为主,山地坡度大多在30°以上,海拔800 m以上以黄壤为主。中亚热带季风气候是本地区地带性气候,降水充沛,相对湿度大,水热同步,光温互补,由于海拔较高,平均温度较低。据百山祖保护站气象观测点(海拔1 100 m)的观测资料显示保护区年均气温12.8℃,年降水量2 341.8 mm,相对湿度84%,10℃以上年积温3 653.3℃,极端高温32℃,极端低温-13℃,无霜期187 d[10]。由于受到人类长期开发的影响,原生植被很少保存,常绿阔叶林几近消亡,仅在北坡中山地带(海拔1 300 ~ 1 700 m)还保存着较大面积的常绿阔叶林,具有重要的保护和研究价值。
2 研究方法
2.1 样地设置
2002-2003年,中国科学院植物研究所、浙江大学和凤阳山——百山祖国家级自然保护区百山祖管理处合作在百山祖北坡,27° 45′ 35″ N,119° 19′ 20″ E,海拔1 400 ~ 1 600 m,建立1个250 m×200 m的中亚热带常绿阔叶林生物多样性永久监测样地,对百山祖地区生物多样性进行长期监测,该样地又是“中国森林多样性长期监测网络”的一部分[11]。
2.2 调查内容
2003年样地完成第一次普查,调查以5 m×5 m的小样方为基本单位,对样方内存活的以及死亡的乔木和高度为2 m以上的灌木进行挂牌、测量和定位。2008年7-8月展开第一次复查,对先前挂牌的样本生存现状进行调查,测量乔木、灌木的胸径(实测)、树高(估测)、冠幅(估测)。而对于样方内新增的个体同样也进行挂牌处理,测量其胸径(实测)、树高(估测)、冠幅(估测),并测量其在小样方中的坐标(直角坐标系)进行定位。
样地中有常绿阔叶树种90种,木荷在本群落乔木层中的重要值排名第二。本研究的所有分析都基于2008年样地复查中记录的木荷数据上进行。
2.3 数据处理
2.3.1 种群结构 出于树木保护的角度出发,采取伐木的方法测定个体年龄并不是最佳选择,而木荷材质较硬,采用钻木取芯的方法测定年龄不仅难度较大,而且要耗费大量人力、物力,与此同时胸径、高度、冠幅等外部特征又不能作为判断年龄的依据,在实际野外工作中精确地组测定种群年龄较为困难,而在相同的环境条件下,木荷的年龄结构和大小级结构能反映出相似的规律,因此采用大小级结构来替代年龄结构分析种群的数量动态[12]。依据木荷种群的生长特点,将木荷种群划分成4等10级:
第1等幼树,划分为两级:第1级(胸径<1.0 cm,基径≤0.8 cm)、第2级(胸径<1.0 cm,基径 > 0.8 cm)。
第2等小树,划分为两级:第3级(1.0 cm≤胸径≤5.0 cm)、第4级(5.0 cm < 胸径≤10.0 cm)。
第3等中树,划分为三级:第5级(10.0 cm < 胸径≤15.0 cm)、第6级(15.0 cm < 胸径≤20.0 cm)、第7级(20.0 cm < 胸径≤25.0 cm)。
第4等大树,划分为三级:第8级(25.0 cm < 胸径≤30.0 cm)、第9级(30.0cm < 胸径≤35.0 cm)、第10级(胸径≥35.0 cm)。
2.3.2 空间分布格局 种群分布格局的测定方法有多种,在实际测量中,不同的方法具有不同的优缺点和适用性,其测定结果也常不尽相同[13]。因此,综合分析各指数测定结果一般可获得可靠结论。本研究中,我们在5 m ×5 m尺度下,采用多个指数对木荷种群空间分布格局进行测定。同时,利用种群不同大小等级结构分布格局的变化来揭示种群发育过程中分布格局动态变化。本研究还分析比较幼树、小树、中树、大树4等群体的分布格局的变化。由于t检验的可靠性与个体平均密度有关[14],个体平均密度过高或过低都会影响t检验的可靠性,因此对于同一数据源来说,运用指标不同所得的结果会有偏差,但平均密度的高低却不影响K值t检验的准确度,因此本研究主要依据K值来进行木荷不同生长阶段的分布格局的比较。
(1)方差均值比法(Cx)
计算方法:

根据Cx值的大小可以判断出种群的分布格局,判断标准如下:当Cx= 1时,随机分布;当Cx< 1时,均匀分布;当Cx> 1时,集群分布[12]。但实际上理论值与实际测量的值由于种种原因会存在偏差,因此可用t值检验确定两者的偏离程度,,在t = t0.05时,差异不显著;t < t0.05,则符合泊松分布,否则为集群分布。
(2)负二项参数(K)
负二项参数起源于二项展开式,K的近似值计算:

K值可用于衡量种群的聚集程度,与种群密度无关。K > 0种群为集群分布,当K值趋向与无穷大时则可认定为随机分布,并且K值越小,种群的聚集程度越大。
(3)丛生指数(I)
David和Moore于1954年提出[16],丛生指数计算公式:
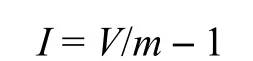
当I < 0时为均匀分布,I = 0时为随机分布,I > 0时为集群分布。
(4)平均拥挤度(m*)与聚块性指标(C)[13]
平均拥挤度指的是每个个体在相同的取样单位中遇到其他个体的平均数,计算公式:

C是平均拥挤度与样本在样方中个体数均值之比,计算公式:

根据C值的大小可用来判断种群分布情况:C = 1时为随机分布,C < 1时为均匀分布;C > 1时则为集群分布。
3 结果
3.1 木荷群落的大小级结构
大样地中共有木荷1 539株,按如上方法划分为10级,其中幼树所占比例为40.29%,包括1级树488棵,2级树132棵和3级树78棵;小树所占比例为
21.25 %,包括4级树125棵和5级树202棵;中树所占比例为28%,包括6级树268棵和7级树163棵;而大树仅占5.39%,包括8级树60棵、9级树19棵和10级树4棵。从整体来看,木荷的径级结构呈现双峰型。第1级为第一个高峰,从第1级至第2级,数量骤降,第3至第6级数量增长趋势明显,在第6级达到一个峰值后从第7级至第10级,数量下降,其中以第7级至第8级最明显。

图1 木荷径级分布结构Figure 1 Diameter distribution of S. superba
3.2 木荷群落的分布格局
如表2所示,幼树方差/均值(Cx)为2.985,大于1,且t检验表明在该条件下假设成立,幼树为集群分布,对于幼树来说:K = 0.290、I = 1.985均大于0,并且C = 4.448 > 1,通过以上3个指标判定也能够得出相同的结论:幼树为集群分布。同样木荷小树、中树、大树也为都体现出集群分布。
根据K值可以判断出不同发育阶段木荷种群分布格局存在一定的差异,聚集程度大小依次为:小树>幼树>中树>大树,除小树外,整体来说种群的分布格局随着个体的不断发育有逐渐趋于松散的趋势。

表1 木荷种群不同生长阶段的分布格局动态Table 1 Distribution pattern of S. superba at different growth stages
4 讨论与结论
4.1 木荷种群的大小结构
种群的重要特征之一就是年龄结构,研究年龄结构不仅可以揭示种群的未来发展趋势,还可以为研究群落演替史、群落更新提供参考依据。为了研究的方便,以及出于可持续发展的长远眼光考虑,许多学者在进行群落研究过程中都会采用大小结构分析法[15],本研究也采用了“空间序列代替时间变化”的方法,以径级结构代替年龄结构的方法来分析种群的结构和动态。
木荷的幼树中第1径级所占比例最大,达到31.71%,虽然从第1径级到第2径级木荷幼树数量大幅度下降但是总体来说幼树所占比例较大。这主要是由木荷的生物学特性和环境条件所决定。木荷是百山祖植物群落的优势种之一,整个样地含有成年树(5级以上)716棵。据野外种子观察,木荷每年能产生大量的种子,结实量大。同时,在充足的水分、适宜的温度下木荷种子萌发率较高[16]。这就使得第1径级幼苗数量最多。但是幼苗萌发后环境因素对其影响很大,幼苗生长过程中存在一道很强的环境筛——光照。已有的研究表明光合有效辐射是影响木荷光合效率的最主要因素[17]。除木荷外,在百山祖植物群落中还存在其他优势种:短尾柯(Lithocarpus brevicaudatus)、光叶水青冈(Fagus lucida)、多脉青冈(Cyclobalanopsis multinervis)等,这些乔木生长高大,乔木上层树冠不连续,下层郁闭度较高[10,13]。木荷幼苗在发育为小树、中树、大树的过程中,由于所处的环境郁闭度较大,随着幼苗的继续生长,对光照的需求进一步增加,势必会影响木荷幼苗的生长率。再者,样地位于百山祖北坡,较南坡而言北坡光照相对较少,大多数幼苗很难忍受长期的低光照环境而死亡,所以第2径级数量急剧减少。木荷从小树到中树数量又快速增加,这可能与生长因子光照以及木荷自身的生物学特性有关,原因分析如下:①通过局部林窗恢复效应而产生的[13]。由于树木的衰老死亡,自然灾害(夏季台风、冬季雪压和雾淞等)造成了森林群落中树木的倒伏现象。有研究显示飓风引起树木的损坏(不同程度的连根拔起)给森林带来了大小不同的各种林窗,飓风过后的五年中松林死亡率呈现下降趋势[18],林窗的形成可以给林下树木带来充足的阳光;②与人类干扰有关。近几十年来百山祖自然保护区对外开放旅游、参观,虽然百山祖北坡受人为因素干扰较少,但是并不能完全排除人为干扰,即百山祖北坡的样地植被并不是一成不变的原始森林,可能由于砍伐、火灾等人为破坏,致使群落内高大乔木死亡、倒伏,森林群落有轻度退化现象,在一定程度上群落的郁闭度降低,即处于第5、6径级的小树生长环境良好,能够获得充足的阳光;③木荷自身生物学特性。对于木荷来说,不同的发育阶段生长速率可能不同。在同样的级差条件下,幼树和小树胸径增加5 cm与中树和大树胸径增加5 cm,所需年间可能不同。因此,中树期(第5、第6、第7径级)的个体数较多,整个种群处于成熟阶段。从第6径级开始个体数又有减小的趋势,其中第7级至第8级减小幅度最大,随后递减率逐渐减小,原因是第5、第6两级数量较多,直径也较大,个体需要更多的营养、阳光、水分、空间以维持继续生存,所以在资源利用上种内和竞争加剧,而且在样地中还有竞争优势更强的其他乔木种,种间竞争也随之加剧,通过自疏、他疏作用个体数量逐渐减少[19]。大树所占比例为5.39%,为衰退级别。
整体来说木荷大小级结构呈基部、中间宽,顶部窄的形状,其年龄结构属于稳定型,木荷幼树、幼苗储备丰富,中树数量较多,大树数量较少,总体上能够满足种群的自然更替和自我更新[20]。
4.2 木荷种群空间分布格局
种群分布格局是研究种群的动态指标之一,是物种与物种间、物种与生存环境间长时间形成的一种相互作用、适应的结果,大量研究表明绝大多数自然种群都呈现聚集分布[3,21],吴宁在研究贡嘎山优势种群的分布格局时也发现在麦吊杉群落中除建群种和优势种呈随机分布外,其余优势种群的分布格局几乎呈聚集分布[22]。
本研究结果表明木荷幼树、小树、中树、大树均为聚集分布,但是不同发育阶段木荷种群的空间分布格局存在一定的差异,聚集程度依次为:小树 > 幼树 > 中树 > 大树,这是外界生态因素综合影响的结果,也由种群本身的生物学特性所决定[22]。
种子传播距离有限性对木荷分布格局有很大影响。木荷属于山茶科,果实类型为蒴果,较大而坚硬,其内的种子小而轻、有翅,极易从果壳内掉落[23]。又因木荷是百山祖自然保护区的优势种之一每年繁殖季节来临时大量木荷果实成熟,此阶段蒴果略微开裂,受到风力、重力影响质量较小的种子便会从果实中掉落,在风力作用下轻小有翅的木荷种子会产生一段水平扩散距离,但是样地内木荷种群密度较高,风力扩散的木荷种子扩散距离有限,大部分木荷种子分布在母株周围。

图2 木荷的空间分布散点图Figure 2 Spatial distribution of S. superba
环境的空间异质性是造成木荷聚集分布的重要原因。从木荷种群的空间散点图上可以结看到,木荷的分布于地形呈现关联性,沿着山脊成斑块状分布(图 2)。王伟等研究发现,百山祖自然保护区大部分常见种群的生长密度与生境密切相关[24]。如在山脊生境中,阳光、水分充足,木荷生长状况良好,密度最大,而在沟谷和陡坡生境木荷种群生长状况最差。李先琨等通过研究发现生境异质性不仅在大尺度(生态系统、群落)层面上存在,而且在小尺度(个体,构件)上也被探测到[25]。小尺度的生境异质性表现为斑块性,样地整体上郁闭度较高,但是也存在林窗或者郁闭度不均匀的现象,即不同的生境条件下一些非生物因子(如光照、水分、营养元素等)的分配情况也不同,生长在小生境中的木荷种群也随之受到影响,因此种群的空间分布格局也会产生不同的差异。聚集分布的幼树通过环境筛的筛选,但是环境筛的过滤并不均等,但是总的来说大约 2/3幼苗因光照等非生物因子长期得不到满足,虽然数量大幅度减少,但是由于百山祖自然保护区木荷种群分布较集中,且在一个样方内经常发现有几株拥在一起生长的现象,又因小生境内资源分布的差异性,因此样地中小树呈现出更加强烈的聚集分布。这种分布格局在某种程度上有利于形成木荷小树发育、成长的微环境,也可以增强外界不利环境的抵抗性,能够使木荷种群在森林群落中保持一定的竞争力和优势,有利于增加个体的成活率。
种内、种间竞争造成木荷空间分布的聚集程度随着年龄的增加不断下降。在种群生长发育的整个过程中,聚集分布格局并不是完全有利的。幼树向中树的发育过程中聚集程度明显减小,随着小树个体长大,其对空间、营养资源的需求也在增加,聚集分布的小树种群内种间竞争加剧。根据密度制约效应,生长在母株周围的子株由于分布密集,较分布于母株较远的子株来说对资源的竞争更加激烈,而来自母株的病菌侵害的风险较高,因此前者死亡率更高[26]。再加上百山祖自然保护区易受自然因素的干扰。如台风、冻雨形成的雾淞及大雪带来的树木倒伏、损坏能够为森林群落带来自然林窗,林窗的干扰使个体生长的微环境中光照量增加,空气流通速度加快,种间竞争加剧,综合以上因素小树到大树的发育过程中种群呈扩散趋势。随着种群的继续发育,种内、种间竞争更加激烈,为了争夺更多的生态位,个体死亡率提高,扩散程度加大,聚集分布格局不明显,这也是植物种群适应环境的一种对抗策略。
综上所述可以得出如下结论:
(1)木荷的种群结构呈稳定型,其中的幼苗更新良好,由于林窗出现的随机性,其种群也存在一定的波动。
(2)木荷种群各个生长阶段分布格局均为集群分布,且集群程度随着年龄的增长逐步降低。说明种内竞争和密度制约效应对木荷的分布格局有一定影响,但其确定性作用的是木荷种子的扩散方式和环境异质性是其形成聚集格局的主要原因。
[1] 张育新,马克明,祁健,等. 北京东灵山海拔梯度上辽东栎种群结构和空间分布[J]. 生态学报,2009,29(6):2 789-2 796.
[2] 苏志尧,吴大荣,陈北光. 粤北天然林优势种群结构与空间格局动态[J]. 应用生态学报,2000,11(3):337-341.
[3] Luo Z R, Ding B Y, Mi X C, et al. Distribution patterns of tree species in an evergreen broadleaved forest in eastern China[J]. Front Biol China, 2009, 4(4):531-538.
[4] 宋坤,孙文,达良俊. 米槠—木荷林优势种群的年龄结构及其更新策略[J]. 生态学报,2011,31(19):5 839-5 850.
[5] 吴翠,董元火,王青锋. 水蕨(Ceratopterist halicroides)种群的分布格局[J]. 武汉大学学报,2004,50(4):515-519.
[6] 蔡飞,宋永昌. 武夷山木荷种群结构和动态的研究[J]. 植物生态学报,1997,21(2):138-148.
[7] 刘郁,李琪安,刘蔚秋,等. 深圳围岭公园植被类型及主要植物群落分析[J]. 中山大学学报(自然科学版),2003,42(S2):14-22.
[8] 彭少麟. 森林群落植物种群分布格局变因探讨[J]. 生态科学,1984,7(3):214-221.
[9] 胡喜生,洪伟,吴承祯,等. 木荷天然种群生命表分析[J]. 广西植物,2007,27(3):469-474.
[10] 徐敏,骆争荣,于明坚,等. 百山祖北坡中山常绿阔叶林的物种组成和群落结构[J]. 浙江大学学报(农业与生命科学版),2007,33 (4):450-457.
[11] 陈小荣,李乐,夏家天,等. 百山祖亮叶水青冈种群结构和分布格局[J]. 浙江农林大学学报,2012,29(5):647-654.
[12] 党海山,江明喜,田玉强,等. 后河自然保护区珍稀植物群落主要种群结构及分布格局研究[J]. 应用生态学报,2004,15(12):2 206 -2 210.
[13] 潘霞,周荣飞,顾莎莎,等. 百山祖北坡常绿阔叶林多脉青冈种群结构和分布格局[J]. 亚热带植物科学,2013,42(3):227-232.
[14] 韩路,王海珍,彭杰,等. 塔里木河中游灰杨种群的径级结构与格局动态[J]. 干旱区资源与环境,2009,23(11):183-185.
[15] 哀建国,丁炳扬,丁明坚. 凤阳山自然保护区福建柏种群结构和分布格局研究[J]. 西部林业科学,2005,34(3):45-49.
[16] 边才苗. 木荷种子萌发及对干旱胁迫的响应[J]. 福建林业科技,2005,32(3):112-115.
[17] 张文标,金则新,柯世省,等. 木荷光合特性日变化及其与环境因子相关性分析[J]. 广西植物,2006,26(5):492-498.
[18] Xi W M,Robert K P,Dean L U. Changes in forest structure,species diversity and spatial pattern following hurricane disturbance in a PiedmontNorth Carolina forest USA[J]. J Plant Ecol,2008,1(1):43-57.
[19] 陈小勇,张庆费,吴化前. 黄山西坡青冈种群结构与分布格局研究[J]. 生态学报,1996,16(3):325-327.
[20] 刘荟,韩庆杰,孙学刚. 甘肃省屈吴山自然保护区不同群落类型中白桦的大小级结构与种群动态[J]. 甘肃农业大学学报,2004,39(2):208-212.
[21] 张峰,上官铁梁. 山西赤果油树群落优势种群分布格局研究[J]. 植物生态学报,2000,24(5):590-594.
[22] 吴宁. 贡嘎山麦吊杉群落优势种群的分布格局及相互关系[J]. 植物生态学报,1995,19(3):270-279.
[23] 姜新. 我国木荷研究现状与展望[J]. 河北农业科学,2013,17(1):42-45.
[24] 王伟,骆争荣,周荣飞,等. 百山祖常绿阔叶林木本植物的生境相关性分析[J]. 生物多样性,2011,19(2):134-142.
[25] 李先琨,黄玉清,苏宗明. 元宝山南方红豆杉种群分布格局及动态[J]. 生物多样性,2000,11(2):169-172.
[26] Luo Z R,Mi X C,Chen X R,et al. Density dependence is not very prevalent in a heterogeneous subtropical forest[J]. Oikos,2012,121(8):1 239-1 250.
Population Structure and Distribution Pattern of Schima superba in Baishanzu
CHEN De-liang1,GU Sha-sha2,DING Bing-yang2,LUO Zheng-rong3
(1. Baishanzu Office of Fengyangshang-Baishanzu National Nature Reserve of Zhejiang, Qingyuan 323800, China; 2. Life and Environment College of Wenzhou University, Wenzhou 325035, China; 3. Ecological School of Lishui College, Lishui 323000, China)
To study the population structure and spatial distribution pattern of Schima superba, a permanent plot of 5 hectares was established in the north slope of Baishanzu Nature Reserve of Zhejiang in 2003. Location and size of each S. superba individuals in the plot was recorded in 2003 and checked in 2008. The size structure of the population of S. superba was analyzed. Distribution pattern at different growth stages was assessed by variance/mean value, negative binomial parameter, index of mean crowing and of patchiness. The results showed that diameter distribution of the population was bimodal, with abundant seedlings and medium-sized trees, which indicating good regeneration in the plot. The results also showed that the population of S. superba at different growth stages had clumped distribution pattern, but the clumping degree was decreased with tree age. And the distribution pattern was formed by seed dispersal limitation and habitat heterogeneity, as well as by intraspecific competition and density.
∶clumped; bimodal; regeneration; density; competition
S718.54
A
1001-3776(2015)01-0001-07
2014-02-21;
2014-08-25
陈德良(1962-),男,浙江庆元人,高级工程师,从事生物多样性保护研究。
