藏鸡THRSP基因的克隆及生物信息学分析
2015-12-17聂晓庆徐亚欧左璐璐林亚秋
聂晓庆,徐亚欧,吕 明,左璐璐,李 想,林亚秋
(西南民族大学生命科学与技术学院,四川 成都 610041)
藏鸡THRSP基因的克隆及生物信息学分析
聂晓庆,徐亚欧,吕 明,左璐璐,李 想,林亚秋
(西南民族大学生命科学与技术学院,四川 成都 610041)
对藏鸡THRSP基因进行克隆、序列测定及生物信息学分析.结果表明:获得的藏鸡THRSP基因核苷酸序列为428 bp(GenBank登陆号:KT153607),在50-258 bp处存在1个CpG岛,ORF长度为390 bp,编码129个氨基酸,其中Leu所占比例为10.9%.THRSP蛋白分子量为14.18 ku,等电点(pI)为4.61,属于不稳定酸性核蛋白,存在8个磷酸化位点和5个O糖基化位点.预测其二级结构中α螺旋占51.2%,β折叠占3.1%,无规则卷曲占45.7%.同源性分析结果表明:藏鸡与原鸡THRSP基因核苷酸及预测的氨基酸序列同源性为100%,两者亲缘关系最近.
藏鸡;THRSP;基因克隆;生物信息学分析
甲状腺激素应答点14(Thyroid hormone responsive spot 14,THRSP)是参与控制脂肪合成代谢的转录调控因子,该基因主要通过启动子区的3个甲状腺素应答元件来对甲状腺激素产生快速应答反应[1]. THRSP基因编码的蛋白是一种小的酸性细胞核内蛋白质,又叫Spot 14[2],在脂类代谢中起重要的调控作用[3].Spot14是1981年Seelig等[4]从小鼠肝脏中提取的mRNA中发现的,随后Cogburn等[5]利用微阵列芯片技术在鸡肝脏中分离并得到确定,被定位于鸡1号染色体长臂41-44处(GGAlq41-44)[6],随后Wang等[7]进一步发现并电子克隆了2个THRSP同源cDNA序列.Wang等[8]研究发现THRSP mRNA主要在肝脏中存在高表达.闫文龙等[9]检测杏花鸡与隐性白洛克鸡F2代12个家系共439只个体基因型及其与脂肪性状的相关性发现,THRSPα基因与脂肪带宽和腹脂重呈显著相关(P<0.05),说明该基因与腹脂性状密切相关[7].因此该基因可作为一个研究脂肪性状的候选基因用于优质肉鸡的辅助育种中.藏鸡属于肉蛋兼用型优质独特地方鸡种,具有肉质鲜美、体型轻小、胸腿肌肉发达、活泼好动、耐粗放等特征[10-11].由于近年来外种鸡及杂鸡的引进,藏鸡作为国家级畜禽品种遗传资源保护品种有濒临灭绝的危险.因此,本研究对藏鸡THRSP基因序列进行克隆与生物信息学分析,为藏鸡地方良种保护和培育提供重要的基础资料.
1 材料与方法
1.1 样品采集
实验所用藏鸡(1只公鸡)来自四川成都益生康健农业有限公司,屠宰后立即取藏鸡的肝脏组织,置于液氮中,带回实验室放于-80℃冰箱中保存备用.
1.2 主要试剂
Trizol(Invitrogen);反转录试剂盒(Thermo Scientific);Taq酶及DH5α感受态细胞(天根生化科技有限公司);凝胶回收试剂盒(Axygen);pMD19-T载体(Takara);其他均为国产分析纯.
1.3 藏鸡THRSP基因克隆
1.3.1 引物的设计与合成
根据GenBank中登陆的原鸡THRSP基因序列(NM_213577),用Premier5.0设计克隆引物,上游引物:5′-GAGGAGAGGCAAGGAGTGG-3′,下游引物: 5′-TCTCCGTGTGCAGGCTCA-3′,送上海生物工程股份有限公司合成.
1.3.2 藏鸡肝脏总RNA提取
用Trizol法提取肝脏组织总RNA,溶解于30 μ LRNAase-free水中,用分光光度计检测总RNA的浓度且达到后续实验的要求.用反转录试剂盒对总RNA进行反转录,得到cDNA.
1.3.3 PCR扩增
PCR扩增体系共25 μL:ddH2O 9.5 μL,cDNA1 μ L,Taq聚合酶12.5 μL,上下游引物各1 μL.PCR反应程序:94℃预变性(5 min);94℃变性(30 s),58℃退火(30 s),72℃延伸(45 s),共35个循环,72℃延伸(5 min).取5 μLPCR产物,用1%琼脂糖凝胶进行检测,将目的条带用胶回收试剂盒进行纯化、回收.
1.3.4 克隆与测序
取纯化回收的PCR产物4.5 μL与pMD-19T载体0.5 μL在5 μL Solution连接液中16℃连接45 min,构建pMD19T-THRSP克隆载体,转化至DH5α感受态细胞中,菌落PCR鉴定,挑选5个阳性克隆,送上海生物工程股份有限公司双向测序.
1.4 藏鸡THRSP基因生物信息学分析
1.4.1 序列分析
通过NCBI网站中的ORF Finder寻找开放阅读框(open reading frame,ORF).将获得的cDNA序列和氨基酸序列在NCBI上进行Blast同源比对,采用MEGA5.0和clustalx1.83构建系统进化树.
1.4.2 Spot 14分析
通过ProtParam在线分析Spot14的理化性质;采用NetPhos 2.0、NetOGlyc 4.0、NetNGlyc 1.0、NetPhosK 1.0、SignalP 4.1Server分别分析预测Spot14磷酸化位点、O糖基化位点、N糖基化位点、保守特异性蛋白激酶作用位点及信号肽;采用PHYRE2 Serve、Predict-Protein、SMART程序分析预测Spot14三级结构、二级结构及结构域.最后进行CpG岛预测(http:// www.ebi.ac.uk/Tools/seqstats/emboss_cpgplot/).
2 结果与分析
2.1 生物信息学分析
克隆的藏鸡THRSP基因序列片段长度为428 bp,包含390 bp的完整开放阅读框,编码129个氨基酸(图1),氨基酸组成如图2.其中氨基酸残基Leu所占频率最高(10.9%),且负电荷的氨基残基数(Asp +Glu=19)多于带正电荷的氨基酸残数(Arg+Lys=10).Spot14蛋白无N端信号肽,说明该蛋白不属于分泌型蛋白.进一步分析其碱基序列组成:A=23.6%,C=32.8%,G=29.2%,T=14.4%;G+C=62%,高于(A+T)38%,说明THRSP基因CDS区DNA双链较稳定.编码蛋白的理化性质:蛋白质分子质量为14.18ku,等电点为4.61,说明该蛋白为酸性蛋白质;消光系数(M-1cm-1γ=280 nm)为7450,不稳定系数为53.04,说明该蛋白属于不稳定性蛋白;Spot14磷酸化位点分别在第4、8、41、60、65、68、89、114位(图3);O糖基化位点分别在第60、62、65、67、68位(图4);保守特异性蛋白激酶作用位点最强在110残基处(P=0.89).Spot14二级结构中α螺旋占51.2%,β折叠占3.1%,无规则卷曲占45.7%.由图5可知Spot14还有1个209bp长度的CpG岛.

图1 藏鸡THRSP基因核苷酸及预测的氨基酸序列Fig.1 Sequences of THRSP nucleotide and predicted amino acid of Tibetan chicken
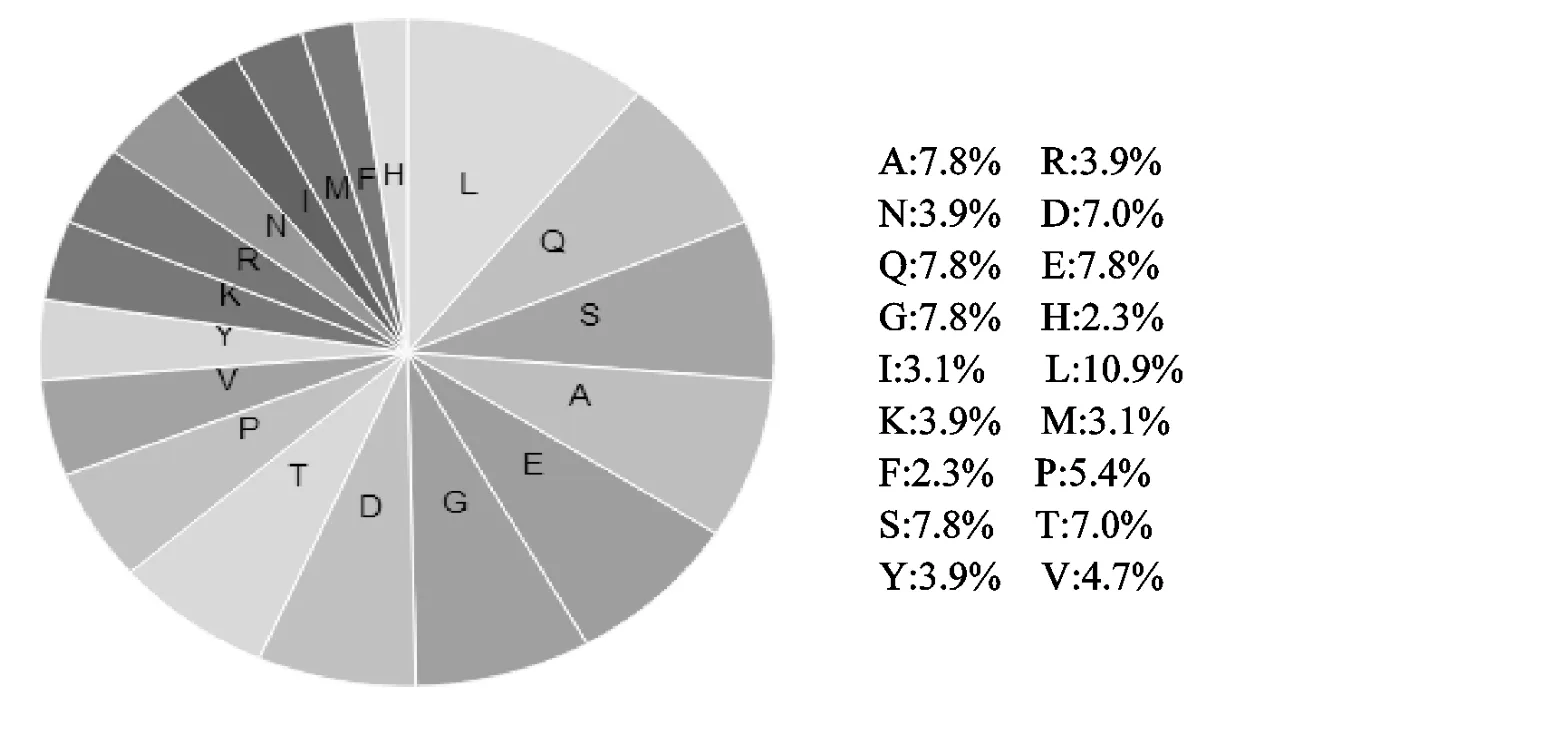
图2 藏鸡Spot14蛋白氨基酸组成Fig.2 The amino acid composition of Spot14 protein of Tibetan chicken

图3 藏鸡Spot14蛋白预测磷酸化位点Fig.3 Predicted phosphorylation sites of Spot14 protein of Tibetan chicken

图4 藏鸡Spot14蛋白预测O糖基化位点Fig.4 Predicted O-glycosylation sites of Spot14 protein of Tibetan chicken

图5 藏鸡Spot14蛋白CpG岛预测Fig.6 Predicted CpG Island of Spot14 protein of Tibetan chicken
由图5可知,50~258bp共有209 bp,(C+G)%>50%,CpG的观察值/预测值比例高于0.6,说明该核苷酸序列中存在1个CpG岛.
2.2 同源性分析
将克隆得到的藏鸡THRSP基因序列、氨基酸序列与原鸡等物种进行同源性分析(表1),并以核苷酸的同源性构建系统进化树(图6).由图7可知,在THRSP基因进化的过程中,藏鸡、原鸡、鸭、白头海雕、褐雨燕、乌鸦处于同一进化枝,其中藏鸡和原鸡亲缘关系最近,其核苷酸、氨基酸同源性100%.日本冠鹮与企鹅处于同一进化枝,原鸽单独处于一枝.

表1 藏鸡THRSP基因与其他已知物种的核苷酸、氨基酸序列比对结果Table 1 Comparison of nucleotide and amino acid sequences between Tibetan chicken THRSP gene and other known species

图6 藏鸡THRSP基因系统进化树Fig.6 The phylogenetic tree of THRSP of Tibetan chicken
3 讨论
本研究利用RT-PCR方法成功克隆藏鸡THRSP基因,获得完整的CDS区390 bp,共编码129个氨基酸,其中氨基酸残基Leu所占频率最高(10.9%).在DNAMAN上将藏鸡与原鸡的THRSP基因CDS区进行比对发现,从第240 bp到249 bp间藏鸡缺失GATGGCATA 9 bp连续的碱基,编码氨基酸为GID,其余位置没有发现碱基的突变.闫文龙[12]发现在鸡THRSPα基因编码区267 bp处存在9 bp的插入/缺失突变,其多态性在不同品种中的分布各不相同.根据核苷酸及氨基酸序列同源性比较结果可以看出,藏鸡与原鸡同源性为100%.从系统进化树也可发现,藏鸡和原鸡亲缘关系最近,其次是鸭,最后是原鸽.而王丛梅等[13]克隆猪的THRSP基因核苷酸(GenBank登陆号:JF_951726)与鸡的同源性比较发现,鸡共有54个碱基的缺失导致移码突变,使两者同源性最远.说明,藏鸡缺失的碱基没有产生移码突变,但可能会影响蛋白质的结构和功能,其内部9bp的缺失突变是否可以作为与脂肪性状连锁的标记用于优质鸡的选择育中值得深入研究.
藏鸡的Spot14蛋白预测磷酸化位点发现共存在8个磷酸化位点:分别在第60、65、68、89、114位氨基酸序列共5个丝氨酸(Ser)磷酸化位点,第8位1个苏氨酸(Thr)磷酸化位点,第4和41位共2个酪氨酸(Tyr)磷酸化位点.Spot14蛋白无N糖基化位点,但在第60、62、65、67、68位氨基酸序列存在5个O糖基化位点.翻译后修饰是蛋白质调节功能的重要方式,它与信号传导、细胞周期、生长发育等诸多生物学问题有密切关系[14].并且发现该基因CDS区含有一个CpG岛.高等真核细胞通常对DNA分子上5’-CpCr -3’序列的胞嘧啶进行甲基化修饰,同时也是最重要的表观遗传学修饰之一.研究发现,CpG岛甲基化水平与癌变密切相关[15].推测藏鸡连续9 bp的缺失与CpG岛有关,具体该位点CpG岛的甲基化状态及其与缺失碱基的关系、对蛋白功能的影响需要深入研究.
王丛梅等[13]检测了猪THRSP基因在14个组织的表达情况,发现在肝脏、脂肪等组织中表达水平最高,在脑、脾脏、胃等组织中表达量相对较低,在肾脏、肌肉中不表达.Wang等[8]也发现,鸡THRSP基因在肝脏中高表达.肝脏是脂肪代谢的重要的器官.说明,该基因及其编码的蛋白spot14确实在肝脏等脂肪组织代谢中发挥着重要的作用,并能有效的调控脂肪合成酶系基因的表达且已被广泛研究[16].Cao[17]等研究表明,鸡THRSPα基因9 bp插入/缺失和A213C突变与体重相关,说明该基因与鸡的生长具有重要的遗传效应.赵春平等[18]利用Agilent基因芯片对牛背最长肌组织样品进行差异表达基因分析发现,THRSP可能是影响牛肉质性状的重要基因.詹凯[19]经选择性Z检验表明,THRSP基因在鸡基因组上的选择作用的概率值P=0.02<0.05.说明THRSP基因是影响动物脂肪生成的重要候选基因且在鸡育种中具有较强的选择作用.下一步,可从基因表达与肌内脂肪含量相关性进行分析,为优质鸡的育种提供新思路.
4 结论
本研究成功克隆得到藏鸡THRSP基因序列,并预测了编码蛋白的结构,为下一步进行组织表达情况及该基因生物学功能研究奠定了基础,为最终阐明藏鸡优良肉质形成的分子机制提供依据.
[1]DECUYPERE E,VAN AS P,VAN DER GEYTEN S,et al.Thyroid hormone availability and activity in avian species:A review[J].Domest Anim Endocrinol,2005,29(1):63-77.
[2]曹志平,李辉.Spot14基因研究进展[J].东北农业大学学报:自然科学版,2008,39(2):285-288.
[3]CHOU WY,CHENG YS,HO CL,et al.Human spotl4 protein interacts physically and functionally with the thyroid receptor[J].Biochem Biophys Res Commun,2007,357(1):133-138.
[4]SEELIG S,LIAW C,TOWLE H C,et al.Thyroid hormone attenuates and augments hepatic gene expression at a pretranslational level[J]. Proc Natl Acad Sci USA,1981,78(8):4733-4737.
[5]COGBUM L A,WANG XF,CARRE W,et al.Systems-wide chicken DNA microarrays,gene expression profiling and discovery of functional genes[J].Poultry Sci,2003,82(06):6378-6383.
[6]CARRE W,DIOT C,FILLON V,et al.Development of 112 unique expressed sequence tags from chicken liver using an arbitrarily primed reverse transcriptase-polymerase chain reaction and single strand conformation gel purification method[J].Anim Genet,2001,32(6):289-297.
[7]WANG X,CARRE W,ZHOU H J,et al.Duplicated Spot 14 genes in the chicken:characterization and identification of polymorphisms associated with abdominal fat traits[J].Gene,2004,332(5):79-88.
[8]WANG X,CARRE W,REJTO L,et al.Global gene expression profiling in liver of thyroid manipulated and or growth hormone injected broiler chickens[J].Poultry Sci,2002,81(Suppl.1):63.
[9]闫文龙,何超,杨关福,等.THRSP基因多态性与鸡肉质、屠体及脂肪性状的关联分析[J].江苏农业科学,2007,3:142-144.
[10]陈国宏,王克华,王金玉,等.中国禽类遗传资源[M],上海:上海科学技术出版社,2004:72-73.
[11]刁运华.四川畜禽遗传资源志[M].成都:四川科学技术出版社,2009:114-116.
[12]闫文龙.THRSP基因对鸡脂肪沉积的影响[D].广州:华南农业大学,2003.
[13]王丛梅,郭豫杰,王月影,等.猪甲状腺激素应答spot14基因的克隆及组织表达研究[J].中国畜牧兽医,2012,39(3):55-59.
[14]DELOM F,CHEVET E.Phosphoprotein analysis:from proteins to proteomes[J].Proteome Sci,2006,4:15.
[15]TSUMURA A,HAYAKAWA T,KUMAKI Y,et al.Maintenance of self -renewal ability of mouse embryonic stem cells in the absence of DNA methyltransferases Dnmtl,Dnmt3a and Dnmt3b[J].Genes Cells,2006,11(7):805-814.
[16]CHOU W Y,CHENG Y S,HO C L,et al.Human spot14 protein interacts physically and functionally with the thyroid receptor[J].Biochem Biophys Res Commun,2007,357(1):133-138.
[17]CAO Z P,WANG S Z,WANG Q G,et al.Association of spot14α gene polymorphisms with body weight in the chicken[J].Poultry Sci,2007,86(9):1873-1880.
[18]赵春平.创伤性应激对安格斯牛肉品质影响机制研究[D].陕西杨凌:西北农林科技大学,2012.
[19]詹凯.鸭甲状腺激素协同作用基因的克隆、表达及多态性研究[D].北京:中国农业大学,2006.
(责任编辑:李建忠,付强,张阳,罗敏;英文编辑:周序林,郑玉才)
Cloning and bioinformatics analysis of THRSP in Tibetan chicken
NIE Xiao-qing,XU Ya-ou,LU Ming,ZUO Lu-lu,LI Xiang,LIN Ya-qiu
(School of Life Science and Technology,Southwest University for Nationalities,Chengdu 610041,P.R.C.)
Cloning,sequencing and bioinformatics analysis of THRSP gene of Tibetan chicken were carried out.The results showed that the cDNA length of THRSP gene was 428 bp(GenBank accession number:KT153607)with a CpG island located in 50-258 bp.The ORF of THRSP gene with a length of 390 bp encoded a protein of 129 amino acids in which Leu residue had a high proportion of 10.9%.The molecular weight was 14.18 ku,and its theoretical isoelectric point was 4.61.The protein belongs to unconventional and acidity nucleoproteins with 8 phosphorylation sites and 5 O-glycosylation sites.The predicted secondary structure of the protein was composed of α helix(51.2%),β sheet(3.1%)and random coil(45.7%).The sequence homology analysis showed that the nucleotide sequence and the deduced amino acids sequence of Tibetan chicken shared 100% homology with Gallus gallus.They have a close genetic relationship.
Tibetan chicken;THRSP;gene cloning;sequence analysis
S813;S831
A
2095-4271(2015)04-0397-06
10.11920/xnmdzk.2015.04.001
2015-04-23
林亚秋(1976-),女,汉族,内蒙古人,副教授,博士,研究方向:动物遗传育种,Email:linyq1999@163.com.
四川应用基础项目(2013JY0044);四川省畜禽育种攻关项目(2011NZ0099-6);西南民族大学项目(2012NFW001)
