海州湾前三岛海域底播刺参群体特征初探
2015-12-15张宏晔杨红生
张宏晔 , 许 强 刘 辉 杨红生
(1.中国科学院 海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071; 2.中国科学院大学,北京 100049)
仿刺参Apostichopus japonicus(Selenka), 又称刺参, 营底栖生活, 喜分布于藻床、礁石等生境中[1],是具有重要经济价值的水产养殖种类。刺参的主要养殖方式可分为: 池塘养殖、围堰养殖、沉笼养殖、浅海底播养殖等[2]; 其中底播养殖方式是将刺参苗种直接撒播到海底自然生长, 因此产品的品质较好[2]。底播养殖按照养殖海域可以分为近岸海域和岛屿海域。与近岸海域相比, 岛屿海域的潮流畅通, 受陆源输入影响小, 水质优良, 且大部分为岩礁底质, 适宜开展刺参的底播养殖。
目前, 国内有关刺参群体生物学特征的相关研究多集中在池塘、围堰等人工养殖系统[2-5]。国外对不同种类海参群体生物学特征与生境关系的研究较多, 如 Kazanidis等[6]研究了爱琴海海参Holothuria tubulosa的种群动态, Toral-Granda等[7]研究了加拉帕戈斯群岛海域海参Isostichopus fuscus的种群结构以及增殖特征, Ramón等[8]研究了地中海海参Stichopus regalis的分布和资源开发, Navarro等[9]研究了北美大加那利岛海参Holothuria arguinensis的生境选择特征, Ginny等[10]研究了加州海峡群岛海参Pachythyone rubra的分布与环境因素的关系。
前三岛位于黄海海州湾内, 距内陆沿岸最近距离41 km, 近岛海域大型藻类资源丰富, 是刺参在中国自然分布的最南界[1]。2003年起, 该海域就已成功开展刺参底播养殖, 但迄今尚无对其群体生物学特征的系统调查研究。该海域水深变化幅度大, 底质类型复杂, 底栖生境多样, 养殖刺参群体分布与其所处生境类型之间的关系亟待揭示。
本研究采用潜水样带法, 研究了海州湾前三岛海域底播刺参群体的周年变动和体质量结构的变化, 比较了不同生境中刺参的分布密度, 探讨了刺参分布与生境类型的关系, 以期揭示该海域底播刺参群体变动规律, 同时也可为岛屿刺参资源保护提供基础数据。
1 材料与方法
1.1 海域位置
调查海域为海州湾前三岛(平岛、达山岛、车牛山岛)中的平岛海域(35°08′12″~35°08′36″N, 119°54′6″~119°55′0″E, 见图1)。2012 年秋季, 在平岛北部东北湾和西北湾分别投放规格为 10~25 g/头的刺参苗种 5 000 kg。
1.2 调查方法与内容
1.2.1 环境因子调查
在刺参非夏眠期间2013年3月26日、5月2日、11月16日和2014年4月5日在前三岛进行调查。在上述调查时间以及2013年8月15日对环境因子进行调查, 用多参数水质测定仪(YSI-650)测定了两个海湾的表层(1 m), 中层(5 m), 底层(10 m)海水前三岛海域的水质特征(水温、盐度、溶解氧)。

图1 研究区域及海湾示意图Fig.1 The sketch map of the research zone & bay
1.2.2 刺参群体特征的调查
在4个调查季节对刺参进行潜水调查。除2014年 4月养殖公司存在刺参采捕行为, 其余时间刺参群体无人为扰动。
采用改进的样带调查法: 使用长50 m、直径6 mm的钢丝绳, 每5 m作一个标记、共10个标记, 作为调查样带引导绳。在岛北东北湾和西北湾内随机布设样带, 并记录样带两端经纬度。样带上每个标记点周围形成以标记点为中心的2 m ×2 m的正方形样方,每条样带形成10个样方, 面积共40 m2。潜水员在水下沿样带采集每个样方内所有刺参。取样后立即计数, 并在 0.5 h内称取个体质量(湿质量)。根据实际情况, 每个海湾分别布设1~4条样带。
1.2.3 不同生境刺参密度的调查
2013年5 月对不同生境中刺参的分布密度进行了调查。根据潜水员观察和水下录像分析, 将前三岛海域底栖生境分为有藻类覆盖的礁石区、无藻类覆盖的礁石区和泥沙质区 3种类型。潜水员在采集刺参的同时, 记录每个样方内的主要生境类型。本次调查共记录了8条样带中80个样方的刺参密度和对应的生境类型。
1.3 统计分析
采用SPSS v21数据统计软件进行数据分析。采用单因素方差分析法(ONE-WAY ANOVA)检验两个海湾水文指标之间的差异。由于刺参密度和生物量并不呈正态分布, 所以对不同季节的刺参密度和生物量采用Kruskal-Wallis H非参数检验法, 此方法在样本不呈正态分布的时候可以对样本之间的差异进行检验。不同调查时间内刺参群体湿质量结构用Kolmogorov-Smirnov单样本检验法检验其分布的正态性, 然后采用 LSD检验法进行比较[6]。不同类型生境间的刺参密度差异采用单因素方差分析法(ONE-WAY ANOVA), 并且在两两不同类型间进行多重比较[11], 显著性水平均设置为0.05。
2 结果
2.1 前三岛海域水质特征
各个监测月份岛东北湾和西北湾之间水质指标无显著差异(P>0.05)。在5次监测中, 3月水温最低为4.39℃, 2013年 8月水温最高为 27.73℃。盐度为29.28~32.20, 溶解氧为 7.02~13.36 mg/L, 最低均为2013年8月, 最高均为2013年3月(表1)。
2.2 刺参密度和生物量的季节变化特征
对不同季节的刺参密度和生物量的 Kruskal-WallisH检验结果表明, 4个调查时间内刺参的密度并无显著性差异(χ2=6.557,P>0.05), 生物量差异也未达到显著性水平(χ2=4.858,P>0.05)。对两个海湾4个季节刺参的多重比较结果表明, 2013年11月东北湾刺参密度和生物量显著高于其他调查月份(P< 0.05), 而西北湾刺参密度则没有显著差异(P>0.05), 2013年3月西北湾刺参生物量显著高于其他月份(P<0.05)。从空间对比上看, 除2013年11月东北湾的刺参生物量显著高于西北湾(P<0.05), 其余调查时间两个海湾的生物量差异不显著(图2)。从空间分布上看, 2013年 11月东北湾刺参较多, 其余季节刺参群体没有明显变化(图3)。

表1 调查期间前三岛海域基本水质指标变动Tab.1 Basic water parameters of Qiansan Islands
2.3 不同规格刺参分布特征
不同调查时间刺参平均湿质量差异显著(F=12.34,P<0.05)。2013年5月刺参群体总体湿质量水平较3月份高, 2013年11月刺参湿质量水平虽较5月份低, 但是个体湿质量分布跨度较小, 2014年4月刺参总体湿质量水平较2013年11月高, 个体湿质量分布跨度最小, 主要集中在75~125g/头。两个海湾刺参湿质量组成, 除东北湾2013年3月刺参湿质量不呈正态分布以外(P=0.023), 其余时间两个海湾的刺参湿质量均呈正态分布(P>0.05), 东北湾的刺参在2013年5月和11月湿质量分布与正态分布曲线拟合效果较好, 2014年4月稍差。西北湾2013年5月份湿质量分布与正态分布曲线拟合最好, 其他月份拟合效果稍差(图4)。
2.4 不同生境类型的刺参群体分布特征
单因素方差分析结果表明, 有藻类分布的礁石区的刺参密度显著高于无藻类分布的礁石区(东北湾F=12.06,P<0.05, 西北湾F=71.70,P<0.05), 详见图5。无藻类分布的礁石区刺参密度高于泥沙质平底区,但差异并未达到显著性水平。
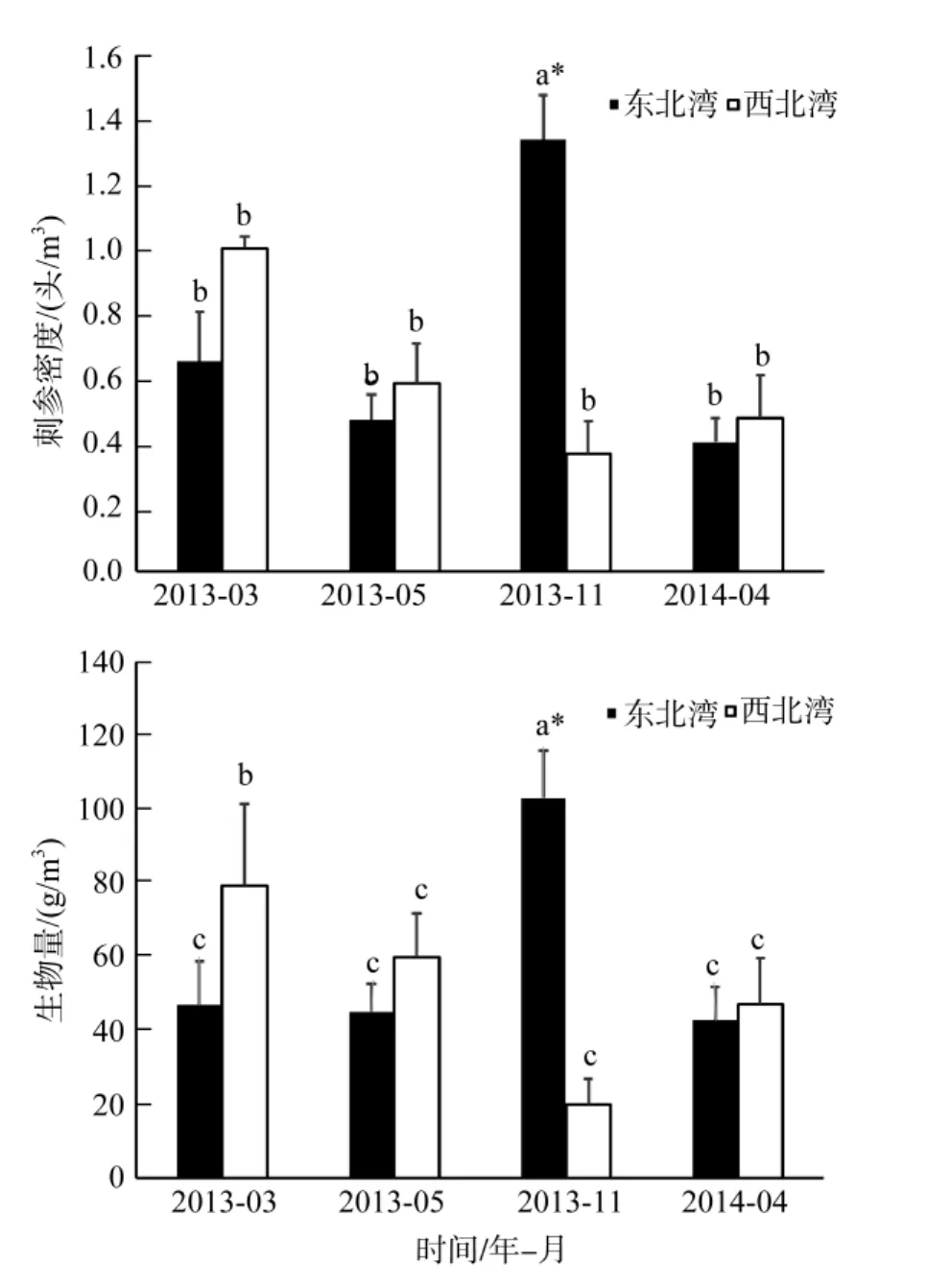
图2 不同季节不同海湾的刺参密度及生物量统计Fig.2 The population & biomass density of Apostichopus japonicus in different bays and different seasons
3 讨论
3.1 底播养殖刺参群体特征
底播养殖刺参在自然环境中生长, 此过程与野生海参种群类似。野生海参的种群在自然状态下种群密度较低, 种群结构相对稳定[6-7]。本研究中, 前三岛海域底播养殖刺参密度和生物量在周年调查内虽然表现一定季节变化, 但是差异并不显著, 可能是由于每次取样的地点数量不同而体现的差异。秋季东北湾的刺参较多, 原因可能是调查样带所在底质类型符合刺参的偏好而出现刺参的聚集行为[12]。本研究中底播刺参的生物量为 42.69 ~103 g/m2, 并未达到该海域的刺参养殖容量109.4 g/m2[13], 刺参密度为0.4~1.33 头/m2, 同为楯手目的海参Australostichopus mollis在野生条件下种群密度最高可以达到0.6 头/m2[11],与本研究中的刺参密度接近。前三岛海域的刺参底播养殖密度较低, 接近野生状态海参, 所以品质较好。

图3 4个调查时间刺参的空间分布Fig.3 Spatial distribution of Apostichopus japonicas in 4 survey season
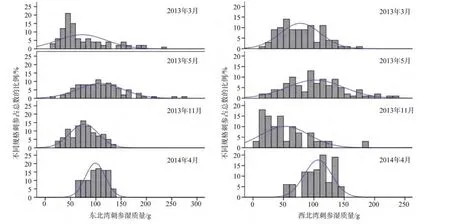
图4 调查时间内刺参不同湿质量组成分布Fig.4 The wet weight distribution of Apostichopus japonicas
由于目前对海参的年龄缺乏统一的界定方法,所以关于海参的种群结构研究主要是利用体长、湿质量等生物学指标判别[14]。在野生海参群体的研究中, 海参的湿质量组成近似正态分布[6-8], 海参的分布和湿质量也受到多种因素的影响, Navarro等[9]的研究表明较高的 POM 含量对海参Holothuria arguinensis有吸引作用, Slate等[15]的研究说明营养较丰富的底质类型有利于海参Australostichopus mollis

图5 不同类型生境刺参密度Fig.5 The population density of Apostichopus japonicus in different habitats
的生长, 而刺参在夏眠过程中会出现能量的消耗而使得湿质量减小[5,16], 恢复摄食以后, 能量收入增加,湿质量也随之增加[17]。在本研究中, 前三岛海域冬春季节(2013年3月到5月)水温上升, 刺参摄食增加[18],湿质量增长。秋季(2013年11月) 降至18℃以下时,刺参由于夏眠期间身体消耗[16]较大且此时刚恢复摄食不久, 因而湿质量较春季下降; 2013年 11月到2014年 4月期间, 刺参恢复正常摄食, 湿质量快速增长。2013年 3月, 东北湾刺参湿质量分布并不符合正态分布, 其原因是群体中有一部分个体是 2012年前投入的较大规格的底播刺参。其他调查时间两个湾由于没有人为干扰及投放等因素, 刺参湿质量均呈正态分布, 符合一般海参的湿质量组成特征。
3.2 岛屿海域底播刺参分布与生境的关系
野生海参群体在自然状态下常呈不均匀的斑块状分布, 其原因主要是不同生境类型在食物供给[10],底质类型[11], 隐蔽场所[10,19]等方面的差异导致了海参对不同生境的倾向性选择。已有研究发现在人工码头环境中, 刺参选择易于躲避并且食物充足的码头地基生境[20]。在本研究的离岸岛屿生境中, 底播养殖刺参对有藻类分布的礁石生境具有明显的倾向性,从实验结果看在西北湾倾向性比东北湾更明显, 这可能是由于西北湾藻类较少导致刺参的聚集。海藻(草)床是刺参喜栖息的环境, 多种来源的丰富的有机碎屑可为刺参提供充足的优质饵料[21-22], 而且大型藻类可以为刺参的活动、隐蔽、繁殖等提供丰富的栖息空间。本研究同时发现, 刺参也倾向于选择无藻床分布的礁石底质。自然或人工礁体环境不仅可为刺参提供丰富的栖息空间, 而且可有效地存积水体自然沉积物, 礁体基质也可附着底栖微藻和其他微型生物, 大大促进刺参的摄食和生长[23-24]。在离岸岛屿这一特殊生境中, 刺参对藻床、礁石区生境的选择性可让刺参能更有效获得优质饵料保证正常生长,更容易找到隐蔽场所躲避恶劣的海况, 这对于刺参适应岛屿海域浪大流急、供饵力相对较低的特点十分重要。
4 结语
本研究中刺参群体变化规律以及的生境选择性可让刺参能更有效获得优质饵料保证正常生长, 更容易找到隐蔽场所躲避恶劣的海况, 对于刺参适应岛屿海域浪大流急供饵力相对较低的生境特征十分重要, 同时也为合适的刺参底播养殖区开发以及优质人工刺参养殖设施的建设有一定意义。
[1]廖玉麟.中国楯手目海参的研究[C]//中国科学院海洋研究所.海洋科学集刊.北京: 科学出版社, 1984:231-247.
[2]杨红生, 周毅, 张涛, 等.刺参生物学——理论与实践[M].北京: 科学出版社, 2014: 276-277.
[3]乔聚海.刺参池塘养殖技术的研究[J].海洋科学,1988, 4: 1-5.
[4]常亚青, 隋锡林, 李俊.刺参增养殖业现状、存在问题与展望[J].水产科学, 2006, 25(4): 198-201.
[5]Qi Z, Wang J, Mao Y, et al.Feasibility of offshore co-culture of abalone,Haliotis discus hannaiIno, and sea cucumber,Apostichopus japonicus, in a temperate zone [J].Journal of the World Aquaculture Society,2013, 44(4): 565-573.
[6]Kazanidis G, Antoniadou C, Lolas A P, et al.Population dynamics and reproduction ofHolothuria tubulosa(Holothuroidea: Echinodermata) in the Aegean Sea [J].Journal of the Marine Biological Association of the United Kingdom, 2010, 90(5): 895-901.
[7]Toral-Granda M V, Martínez P C.Reproductive biology and population structure of the sea cucumberIsostichopus fuscus(Ludwig, 1875) (Holothuroidea) in Caamaño, Galápagos Islands, Ecuador [J].MarineBiology, 2007, 151: 2091-2098.
[8]Ramón M, Lleonart J, Massutí E.Royal sea cucumber(Stichopus regalis) in the northwestern Mediterranean:Distribution pattern and fishery [J].Fisheries Research,2010, 105: 21-27.
[9]Navarro P G, García-Sanz S, Tuya F.Contrasting displacement of the sea cucumberHolothuria arguinensisbetween adjacent nearshore habitats [J].Journal of Experimental Marine Biology and Ecology,2014, 453: 123-130.
[10]Eckert G L.Spatial patchiness in the sea cucumberPachythyone rubrain the California Channel Islands[J].Journal of Experimental Marine Biology and Ecology, 2007, 348: 121–132.
[11]Morgan A D.Patterns of distribution and abundance of the temperate sea cucumberAustralostichopus mollison a rocky subtidal reef [J].New Zealand Journal of Zoology, 2011, 38(3): 195-206.
[12]张俊波, 梁振林, 黄六一, 等.不同材料、形状和空隙的人工参礁对刺参诱集效果的试验研究[J].中国水产科学, 2011, 18(4): 899-907.
[13]邢坤.刺参生态增养殖原理及关键技术[D].青岛:中国科学院海洋研究所, 2009.
[14]Nelson E J, MacDonald B A, Robinson S M C.Robinson, et al.A review of the northern sea cucumberCucumaria frondosa(Gunnerus, 1767) as a potential aquaculture species [J].Reviews in Fisheries Science,2012, 20(4): 212-219.
[15]Slater M J, Jeffs A G.Do benthic sediment characteristics explain the distribution of juveniles of the deposit-feeding sea cucumberAustralostichopus mollis? [J].Journal of Sea Research, 2010, 64: 241–249.
[16]Yang H, Yuan X, Zhou Y, et al.Effects of body size and water temperature on food consumption and growth in the sea cucumberApostichopus japonicus(Selenka)with special reference to aestivation [J].Aquaculture Research, 2005, 36: 1085-1092.
[17]汪洋, 郑忠明.周朝生等.南移刺参摄食三种沉积物饵料下的生长和能量收支比较 [J].生态科学, 2012,31(6): 613-618.
[18]Dong Y, Dong S.Growth and oxygen consumption of the juvenile sea cucumberApostichopus japonicus(Selenka) at constant and fluctuating water temperatures[J].Aquaculture Research, 2006, 37: 1327-1333.
[19]Glenn R.Shiell, Brenton Knott.Diurnal observations of sheltering behavior in the coral reef sea cucumber Holothuria whitmaei [J].Fisheries Research, 2008, 91:112–117.
[20]Yamana Y, Hamano T, Goshima S.Seasonal distribution pattern of adult sea cucumberApostichopus japonicas(Stichopodidae) in Youshimi Bay, western Yamaguchi Prefecture, Japan [J].Fisheries Science,2009, 75: 585-591.
[21]Liu Y, Dong S, Tian X, et al.The effect of different macroalgae on the growth of sea cucumbers(Apostichopus japonicusSelenka) [J].Aquaculture Research, 2010, 41(11): 881-885.
[22]刘旭佳, 周毅, 杨红生, 等.大叶藻碎屑作为刺参食物来源的实验研究 [J].海洋科学, 2011, 37(10):30-36.
[23]秦传新, 董双林, 牛宇峰, 等.不同类型附着基对刺参生长和存活的影响 [J].中国海洋大学学报, 2009,39(3): 392-396.
[24]王吉桥, 张俊涛, 姜玉声, 等.礁石堆密度对仿刺参幼参生长和存活的影响[J].渔业现代化, 2008, 35(6):40-43.
