内蒙古自治区乌兰察布市燕麦炭疽病病原鉴定
2015-11-25何苏琴荆卓琼赵桂琴丁文姣郭满库柴继宽
何苏琴, 荆卓琼, 赵桂琴, 丁文姣, 郭满库, 柴继宽
(1.甘肃省农业科学院植物保护研究所, 兰州 730070; 2. 甘肃省农业大学草业学院, 兰州 730070;3. 甘肃省农业科学院农业质量标准与检测技术研究所, 兰州 730070)
内蒙古自治区乌兰察布市燕麦炭疽病病原鉴定
何苏琴1, 荆卓琼1, 赵桂琴2, 丁文姣3, 郭满库1, 柴继宽2
(1.甘肃省农业科学院植物保护研究所, 兰州 730070; 2. 甘肃省农业大学草业学院, 兰州 730070;3. 甘肃省农业科学院农业质量标准与检测技术研究所, 兰州 730070)
2010年8月,从内蒙古自治区乌兰察布市燕麦炭疽病病叶上分离得到产生镰刀形分生孢子的刺盘孢菌,依据柯赫氏法则、病菌形态特征和生物学特性,将其鉴定为ColletotrichumcerealeManns。病菌分生孢子盘生大量黑色针状刚毛;分生孢子单细胞,镰刀形,无色,(17.43~24.90)μm×(1.99~3.74)μm,平均(22.02±1.80)μm×(2.85±0.36)μm;分生孢子萌生的附着胞黑色,卵圆形或近圆形,(6.23~7.97)μm×(4.98~6.23)μm,平均(7.30±0.52)μm×(5.82±0.48)μm;菌丝生附着胞黑色,近圆形、长椭圆形或裂瓣状,(7.47~14.94)μm×(4.98~9.96)μm,平均(10.87±1.46)μm×(6.74±1.01)μm。病菌菌丝在5~35 ℃可生长,适宜生长温度为25 ℃;在人工接种条件下(离体叶片,23~25 ℃),菌株蒙-3-1对供试的2个裸燕麦品种和8个皮燕麦品种均具有强的致病性,但不侵染小麦、玉米、谷子和高粱。
燕麦; 炭疽病;Colletotrichumcereale; 寄主专化性
禾谷类作物,如小麦、燕麦、大麦的炭疽病,在20世纪的前50年间呈现出间歇性暴发流行的特点。燕麦炭疽病曾于1933-1955年、1963年在北美洲暴发流行[1-2],2004年墨西哥报道了燕麦炭疽病的发生[3]。
20世纪30-60年代,燕麦炭疽病在我国的河北、山西、内蒙古、黑龙江、广西、贵州、云南、吉林有发生记录[4-5],但未见深入的研究报道。
2010年8月,笔者在内蒙古燕麦病害调查中发现乌兰察布市卓资县的一块约1.5 hm2的燕麦田(裸燕麦)炭疽病发生严重,依据柯赫氏法则、病菌形态特征和生物学特性,对卓资县燕麦炭疽病的病原进行了鉴定,报告如下。
1 材料与方法
1.1 病害标样采集
2010年8月6日,从内蒙古乌兰察布市卓资县燕麦田(裸燕麦)采集典型症状的炭疽病病叶,阴干后保存于标本袋带回实验室,用于病原菌分离。
1.2 培养基及配方
PDA:马铃薯200 g,葡萄糖15 g,琼脂粉12 g,自来水1 000 mL。
PSA:马铃薯200 g,蔗糖15 g,琼脂粉12 g,自来水1 000 mL。
1.3 病原菌分离及形态学鉴定
1.3.1 病原菌分离
采用常规组织分离法及孢子稀释法分离病原。将病健交界处病组织剪成大小约0.3 cm×0.5 cm小块,75%乙醇表面消毒3 s,灭菌水冲洗3次,用灭菌滤纸吸干病组织表面水分,置PDA平板上,20 ℃下培养,长出的真菌及时转至无菌PDA平板上,并挑取菌丝尖端进行纯化;从病斑上挑取分生孢子盘及分生孢子,用灭菌水稀释后,取1 mL涂于PDA平板上,20 ℃培养,挑取单孢子菌落。
式中,Ng是指单位面积上所发生的年雷击大地次数,单位为次/(km2·年),反映的是某地区雷击频繁程度[12]。其中,N为湖南省ADTD型雷电监测网提供的某区域地闪总次数,本研究取2009—2017年的年平均雷击大地次数。S为区域面积,单位为km2。
1.3.2 柯赫氏法则证病
选取无破损、无变色的燕麦(裸燕麦:‘白燕2号’,皮燕麦:‘科燕1号’)健康种子;55 ℃温汤浸种;20 ℃催芽;将萌芽种子播种于盛有灭菌蛭石土的营养钵中,待植株生长至3~4片叶时,剪取供试作物健康叶片,置于直径18 cm培养皿内的湿滤纸上准备接种用。
将试验菌株蒙-3-1接种于PDA平板上,(27±2)℃下培养12 d,将培养物研碎,加水调至2×106/mL用于接种。采用悬滴法,将菌悬液接种于叶片上,每个叶片接种1~3滴菌悬液,每滴菌悬液约10 μL,每品种接种10个叶片。以接种清水为对照。接种后,将培养皿置于23~25 ℃、自然散射光下保湿培养,逐日观察发病情况,对发病叶片进行病原菌的重分离。
1.3.3 燕麦炭疽病菌寄主范围测定
种子处理、栽培管理和接种方法同1.3.2。
供试作物及品种:裸燕麦:‘白燕2号’和‘燕2007’;皮燕麦:‘坝燕1号’、‘丹麦444’、‘黄燕麦’、‘白燕7号’、‘科燕1号’、‘陇燕1号’、‘陇燕2号’、‘陇燕3号’;玉米:‘黄早四’;高粱:‘熊岳253’以及小麦:‘蓝天13’。每品种接种10个叶片。
1.3.4 病原菌种类鉴定
依据在自然寄主及接种后发病燕麦上产生的病菌分生孢子盘及分生孢子的形态、附着胞形态等特征进行形态比对,并结合病菌寄主范围进行种类鉴定。
1.4 温度对燕麦炭疽病菌菌丝生长的影响
取PSA平板上,20 ℃下培养5 d的蒙-3-1菌丝块,直径5 mm,置于新鲜PSA平板上,分别在5、10、15、20、25、30、35和40 ℃下黑暗培养7 d,测量菌落直径,每处理重复3次。
2 结果与分析
2.1 燕麦炭疽病症状
燕麦炭疽病菌可以侵染燕麦的叶片及叶鞘。病斑呈黄褐色,梭形,长约2 cm,病斑上可见黑色小粒点,即病菌的分生孢子盘,叶片干燥后病斑中央溃烂撕裂状,见图1。
2.2 燕麦炭疽病病原菌分离及形态学鉴定
用常规组织分离法及孢子稀释法均分离得到菌落形态一致的真菌。原始分离物菌丝灰黑色,在PSA平板上20 ℃下培养15~20 d或25 ℃下培养7~10 d即可产生具黑色针状刚毛的分生孢子盘及镰刀形分生孢子(长期保存或继代培养后,菌落形态和产孢特性可发生改变),将其中具代表性的单孢菌株编号为蒙-3-1。
用蒙-3-1接种燕麦,23~25 ℃下,接种后3~5 d显症,接菌点及周围失绿黄化,或形成浸润斑;或形成黄褐色枯斑。接种后7~10 d,部分发病燕麦叶片上出现分生孢子盘,见图2。对照未发病。对发病组织进行病原菌的再分离,原接种菌的分出率达100%。

图1 燕麦炭疽病田间自然发病症状Fig.1 Symptoms of oat anthracnose in the field
在本文试验条件下,所有裸燕麦和皮燕麦品种均于接种后3~5 d显症,接菌后7 d,病叶率达100%,对照未发病。小麦、玉米和高粱接菌后7 d均未见发病,表明燕麦炭疽病菌在该试验条件下不侵染小麦、玉米和高粱。

图2 燕麦炭疽病菌菌株蒙-3-1接种燕麦离体叶片发病症状Fig.2 Symptoms of oat anthracnose by inoculating strain Meng-3-1 on detached oat leaves
病菌分生孢子盘具有大量黑色针状刚毛;分生孢子为单细胞,镰刀形,无色,大小为(17.43~24.90)μm×(1.99~3.74)μm,平均(22.02±1.80)μm×(2.85±0.36)μm;分生孢子萌生的附着胞黑色,卵圆形或近圆形,大小为(6.23~7.97)μm×(4.98~6.23)μm,平均(7.30±0.52)μm×(5.82±0.48)μm。菌丝生附着胞黑色,近圆形、长椭圆形或裂瓣状,大小为(7.47~14.94)μm×(4.98~9.96)μm,平均(10.87±1.46)μm×(6.74±1.01)μm。见图3。
依据柯赫氏法则及观察到的病菌形态特征,并结合病菌的寄主范围,参考文献[1,6-7]将内蒙古自治区乌兰察布市燕麦炭疽病的病原鉴定为ColletotrichumcerealeManns。
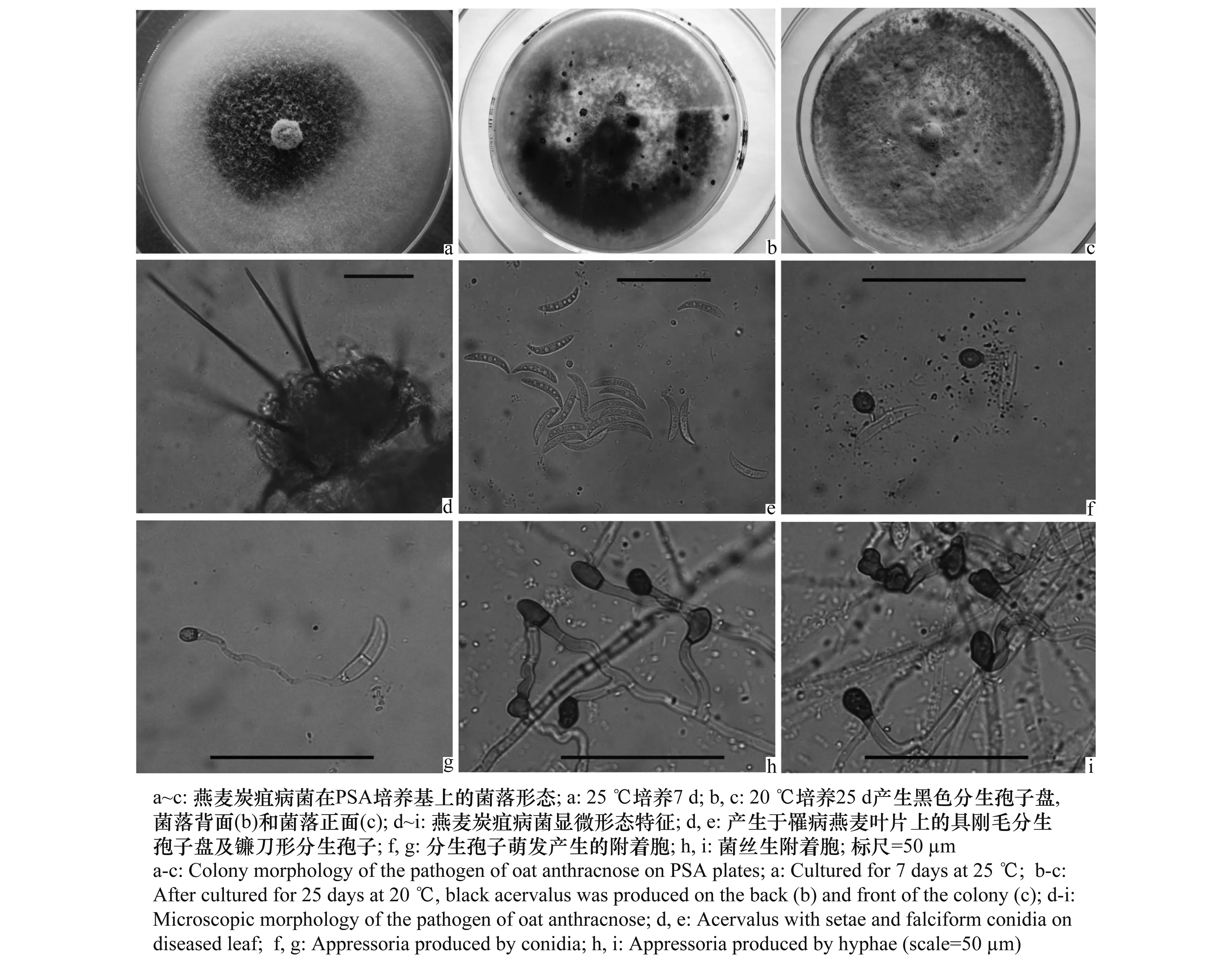
图3 燕麦炭疽病菌形态特征Fig.3 Morphological characteristics of the pathogen of oat anthracnose

图4 燕麦炭疽病菌菌株蒙-3-1在PSA平板上不同温度下培养7 d的菌落直径Fig.4 Colony diameters of strain Meng-3- cultured for 7 d on PSA plates at different temperatures
2.3 温度对燕麦炭疽病菌菌丝生长的影响
从图4可以看出,蒙-3-1的菌丝在5~35 ℃均可生长,最适生长温度为25 ℃。5 ℃下,菌丝表现为直立生长,但菌落不扩展;随着培养温度的升高,菌落直径增加,在25 ℃下培养7 d,菌落直径达83.33 mm;将40 ℃下培养7 d的菌丝块转移至25 ℃下继续培养,4 d后,有1/3的接种菌丝块可恢复生长。
3 讨论
Colletotrichumcereale于1909年由Selby和Manns建立,最初报道发生在小麦、燕麦和无芒雀麦上,由于相似的形态和共同的禾草寄主,1914年被Wilson归作C.graminicola的同物异名;1957年,von Arx将所有的35个禾栖炭疽种都归为C.graminicola的异名,力图使假定的无性时期与新的有性型Glomerellatucumensis相一致。20世纪60年代,Sutton的研究工作又逐渐打破了C.graminicolasensu Arx的种群概念,恢复了C.caudatum,C.falcatum和C.sublineolum,C.graminicolasensuSutton的种群概念随后也得了到广泛的支持,受可用的形态学特征的局限,在之后的40年,C.graminicola仍然被用于描述除甘蔗、高粱或假高粱之外的禾草寄主上的炭疽病菌;2006年,Crouch及其团队采用多位点系统系统发育方法,使得C.cereale作为与早熟禾亚科Pooideae相关联的分类单元得以复活,其中包含了草坪草致病菌和大量的冷季型禾草,如小麦、燕麦和鸭茅的内寄生菌[1]。
C.cereale是一个宿主范围广泛的物种,其寄主包括剪股颖属(Agrostis)、燕麦属(Avena)、雀麦属(Bromus)、拂子茅属(Calamagrostis)、鸦茅属(Dactylis)、披碱草属(Elymus)、羊茅属(Festuca)、茅香属(Hierochloe)、绒毛草属(Holcus)、大麦属(Hordeum)、黑麦草属(Lolium)、早熟禾属(Poa)、棒头草属(Polypogon)、小麦属(Triticum)。尽管自20世纪初以来,对谷物和禾草的炭疽病已经展开了充分的研究,但最近的研究结果仍然显示出,该族群比之前的预期更大更复杂[1]。
从谷类作物、草原、草坪草得到的C.cereale存在明显的种群分化,种群结构相当复杂,可被划分为10个与生态系统和/或寄主植物相一致的高度专化的种群[8]。依据寄主范围和附着胞形态特征对刺盘孢属种的综合评判结果,与源于分子系统的刺盘孢属种群关系高度吻合[6]。
Harder等1963年对加拿大西部亚伯达省的小麦、燕麦、大麦和黑麦炭疽病发生情况进行了调查,结果显示炭疽病的发生与土壤类型无关,但是种植在低有机质土壤中的作物发病最严重,种植在有机质含量很高的土壤中的作物几乎不发病。种植在退化土壤中和山麓丘陵地带的作物发病率较高。土壤因子,如无机盐浓度、硝酸盐、游离碳酸盐和电导率与发病率无关。小麦、燕麦、大麦和黑麦都发现有炭疽病危害,以燕麦受害最严重[2]。
在植株成熟期,受害部会出现黑色具刚毛的分生孢子盘这一标志性特征,但在此特征出现之前,在作物的主要生长期都难以诊断出燕麦炭疽病,明显的病害症状包括植株活力降低、茎秆细弱、分蘖减少、基部叶片枯黄、植株矮化和提早成熟等,这些症状很容易与肥力不足、肥力不均或干旱造成的症状相混淆[2]。
本研究中,菌株蒙-3-1的菌丝在35 ℃下生长极缓慢,生长速率远低于10 ℃下,与Bruehl等[9]报道的燕麦炭疽病分离菌株在36 ℃的生长速率高于12 ℃,适宜生长温度为28 ℃稍有不同。
有资料显示,炭疽病菌可以在燕麦和大麦的根状茎上以菌核形态休眠[1]。炭疽病菌在燕麦种子上的带菌情况及作用尚不清楚。荆卓琼等对甘肃省燕麦种带真菌情况的检测中未检测到炭疽病菌[10]。但种子预处理对防止病菌传播、防治苗期病害、促进壮苗和提高产量具有积极作用[11-14]。与非寄主植物轮作、均衡施肥、清洁田园等是防治炭疽病的有效措施[15]。
[1] Crouch J A, Beirn L A.Anthracnose of cereals and grasses [J].Fungal Diversity, 2009, 39: 19-44.
[2] Harder D E, Skoropad W P.The occurrence of cereal anthracnose in Alberta [J].Canadian Plant Disease Survey,1968, 48(2):39-42.
[3] Leyva-Mir S G, Soto-Herrera A, Espitia-Rangel E, et al. Etiology and incidence of anthracnosis[Colletotrichumgraminicola(Ces.) G.W. Wils.] of oat (AvenasativaL.) in Michoacan, Mexico [J].Revista Mexicana de Fitopatología, 2004,22(3):351-355.
[4] 戚佩坤, 白金铠, 朱桂香. 吉林省栽培植物真菌病害志[M].北京: 科学出版社, 1966:254.
[5] 戴芳澜. 中国真菌总汇[M].北京: 科学出版社, 1979: 920.
[6] Crouch J A, Clarke B B, White J F, et al. Systematic analysis of the falcate-spored graminicolousColletotrichumand a description of six new species of the fungus from warm season grasses [J].Mycologia, 2009, 101(5):717-732.
[7] Hyde K D, Cai L, Cannon P F, et al.Colletotrichum-names in current use [J].Fungal Diversity, 2009, 39: 147-182.
[8] Crouch J A, Tredway L P, Clarke B B, et al. Phylogenetic and population genetic divergence correspond with habitat for the pathogenColletotrichumcerealeand allied taxa across diverse grass communities [J].Molecular Ecology,2009,18(1):123-135.
[9] Bruehl G W, Dickson J G.Anthracnose of cereals and grasses[J].US Department of Agriculture Technical Bulletin, 1950,1005:1-37.
[10]荆卓琼, 丁文娇, 何苏琴, 等. 甘肃省皮燕麦和裸燕麦种子带真菌情况检测[J].种子, 2012, 31(4):31-34.
[11]The next generation seed treatment for wheat and oats, Hombre, file7883[EB/OL].Bayer CropScience Pty Ltd. (2007)[2009-12-20].http:∥www.bayerresources.com.au/resources/uploads/Bulletin/file7883.pdf.
[12]Cereal Seed Treatment Comparison, 6399_SYN_SeedcarePoster.indd [EB/OL].SyngentaFarm.ca (2014-01-29)[2014-04-14].http:∥beyondagronomy.com/cmsFiles/documents/document5318f4b9b41cb.pdf.
[13]何苏琴,郭满库,赵桂琴,等.吡虫啉·戊唑醇种子处理悬浮剂对燕麦红叶病的田间防效[J].甘肃农业科技,2010(12):8-9.
[14]荆卓琼,郭满库,何苏琴,等.吡虫啉·戊唑醇种衣剂对裸燕麦的适宜拌种剂量筛选[J].甘肃农业科技,2013(1):32-34.
[15]Anthracnose of forage grasses and cereals, RPD No. 313[EB/OL].Department of Crop Sciences University of Illinois at Urbana-Champaign (1984. 11)[2014-04-14]. http:∥ipm.illinois.edu/diseases/rpds/313.pdf.
(责任编辑:田 喆)
Identification of the pathogen of oat anthracnose in Wulanchabu City of Inner Mongolia Autonomous Region
He Suqin1, Jing Zhuoqiong1, Zhao Guiqin2, Ding Wenjiao3, Guo Manku1, Chai Jikuan2
(1. Institute of Plant Protection, Gansu Academy of Agricultural Sciences, Lanzhou 730070, China; 2. College of Prataculture, Gansu Agricultural University, Lanzhou 730070, China; 3. Institute of Agricultural Quality Standards and Testing Technology, Gansu Academy of Agricultural Sciences, Lanzhou 730070, China)
Colletotrichumisolates were obtained from diseased leaves of oat anthracnose in Wulanchabu City of Inner Mongolia Autonomous Region in August 2010. Based on Koch’s rule, morphological and biological characteristics, the pathogen of oat anthracnose was identified asC.cerealeManns. Needle-like black setae were observed on acervulus in abundance; conidia were single-celled, falciform, and colorless, (17.43-24.90)μm×(1.99-3.74)μm, av. (22.02±1.80)μm×(2.85±0.36)μm in size; conidial appressoria were black, oval or elliptic, (6.23-7.97)μm×(4.98-6.23)μm, av. (7.30±0.52)μm×(5.82±0.48)μm in size; hyphopodia appressoria were black, rounded, long elliptic or lobate, (7.47-14.94)μm×(4.98-9.96)μm, av. (10.87±1.46)μm×(6.74±1.01)μm in size, Mycelia of the pathogen could grow at 5-35 ℃, and the optimum growth temperature was 25 ℃. Host specificity was confirmed. Under identical conditions (detached leaf inoculation, at 23-25 ℃, relative humidity 100%), strain Meng-3-1 showed aggressive pathogenicity to two hull-less oat varieties and eight hulled oat varieties, but did not infect wheat, maize, millet and sorghum.
oat; anthracnose;Colletotrichumcereale; host specificity
2014-04-16
2014-07-11
现代农业燕麦荞麦产业技术体系建设专项(CARS-08-C-1)
S 435.126
A
10.3969/j.issn.0529-1542.2015.03.014
* 通信作者 E-mail:gshesuqin@sina.com
