Genetic and Morphological Variations within Laudakia microlepis (Blanford, 1874) (Sauria: Agamidae) Populations in Southeastern Iran with Description of a New Subspecies
2015-10-31NasserSANCHOOLIHassanRAHIMIANNasrullahRASTEGARPOUYANIandEskandarRASTEGARPOUYANI
Nasser SANCHOOLI, Hassan RAHIMIAN, Nasrullah RASTEGAR-POUYANIand Eskandar RASTEGAR-POUYANI
1Faculty of Biological Sciences, College of Science, University of Tehran, Tehran, Iran
2Department of Biology, Faculty of Science, Zabol University, Zabol, Iran
3Department of Biology, Faculty of Science, Razi University, Kermanshah, Iran
4Iranian Plateau Herpetology Research Group (IPHRG), Razi University, Kermanshah 6714967346, Iran
5Department of Biology, Faculty of Science, Hakim Sabzevari University, Sabzevar, Iran
Genetic and Morphological Variations within Laudakia microlepis (Blanford, 1874) (Sauria: Agamidae) Populations in Southeastern Iran with Description of a New Subspecies
Nasser SANCHOOLI1,2, Hassan RAHIMIAN1*, Nasrullah RASTEGAR-POUYANI3,4and Eskandar RASTEGAR-POUYANI5
1Faculty of Biological Sciences, College of Science, University of Tehran, Tehran, Iran
2Department of Biology, Faculty of Science, Zabol University, Zabol, Iran
3Department of Biology, Faculty of Science, Razi University, Kermanshah, Iran
4Iranian Plateau Herpetology Research Group (IPHRG), Razi University, Kermanshah 6714967346, Iran
5Department of Biology, Faculty of Science, Hakim Sabzevari University, Sabzevar, Iran
A new subspecies of Laudakia microlepis (Blanford, 1874) is described, from Taftan Mountain in southeastern Iran, based on morphological and genetic characteristics. This new subspecies is distinguished from the nominal subspecies by having a yellowish head along with the following combination of characters: a relatively larger body size; different number of scales around middle of body; larger, weakly keeled, dorsal median scales arranged in 11–13 oblique longitudinal rows; 123–136 scales in a single row from posterior edge of gular fold to vent; 27–31 scales in the first complete whorl around the tail. The type locality of Laudakia microlepis taftanica ssp. nov., is hereby, reported as the Taftan Mountain, about 300 kilometer southeast of the nominal subspecies known range.
Laudakia microlepis taftanica ssp. nov., taxonomic status, ND4 gene, Sistan and Baluchistan province
1. Introduction
The genus Laudakia comprises about 20 species, mainly occurring in highland and mountainous regions of the central and southern Asia. Of those 20 species, at least five have been reported from Iran (Šmid et al., 2014). Using morphology-based, non-phylogenetic analysis of Laudakia species, Baig et al. (2012) divided the genus into three genera; Stellagama, Paralaudakia and Laudakia. But, shortly after that, a robust molecular phylogenetic analysis strongly supported monophyletic status of the genus Laudakia, suggesting that the changes were not supported (Pyron et al., 2013).
Laudakia microlepis was first described by Blanford (1874) from two localities in Iran: Khane-Sorkh Pass,with elevation of about 2 750 m, between Sirjan and Kerman, southeastern Iran, and Kushk-e-Zard in southcentral Iran. Subsequent studies, however, determined that further to southern Iran, L. microlepis is distributed in southern and western Pakistan, Afghanistan, and parts of Turkmenistan (Baig et al., 2012). The taxonomic status of L. microlepis has been controversial ever since its description. For instance, while Blanford (1876) pointed to the very close relationships between L. caucasia and L. microlepis. Wettstein (1953) considered L. microlepis as a subspecies of L. caucasia, Rastegar-Pouyani and Nilson (2002) and Baig et al. (2012) considered the two taxa as distinct species with sometimes sympatric distributions.
In the present communication, we describe a new subspecies of Laudakia microlepis based on comparison between distinguishing morphological characteristics and by comparing mitochondrial ND4 gene sequences, among specimens collected from Taftan Mountain with those from the type locality of L. microlepis.
2. Material and Methods
Fourteen specimens were collected in August 2011, from Taftan Mountain (28o36' 36.6" N 61o04' 51.1" E, elevation 2 530 m), in Sistan and Baluchistan province, southeastern Iran (Figure 1). In addition, five specimens were collected from Khane Sorkh, between Sirjan and Kerman. Specimens are deposited in the Zoology Museum of the University of Tehran (ZUTC).
Morphology Specimens collected from the Taftan Mountain, comprising eight males and six females, were preserved in 70% ethanol and deposited in the Zoology Museum of University of Tehran (ZUTC). All measurements are carried, to the nearest 0.01 millimeter, using vernier calipers, and scales on different parts of the body were counted with aid of a stereomicroscope. The following characteristics were considered: Scales around mid-body (SQ), number of scales in a single row around the widest part of body; Large Vertebral Scales (LVS), number of longitudinal rows of enlarged scales on the vertebral region; Anus-Gular Fold Scales (AGFS), number of ventral scales in a single row from posterior edge of gular fold to the vent; Fifth Caudal Whorl Scales(FCWS) scales around the fifth caudal whorl just behind the vent; Snout-Vent Length (SVL) from tip of the snout to the anterior edge of cloaca; Tail Length (TL) from posterior edge of cloaca to the tip of the tail; Head Length (HL) from tip of the snout to the posterior edge of tympanum; Head Width (HW) measuring widest part of the head; Head Height (HH) from top of the head to the lower jaw. Data were analyzed using statistical software SPSS (version 22). The significance level for all tests were set at P<0.05. Descriptive statistics were carried out to explore the means, standard errors and ranges of the characters.
Averages of those traits for L. caucasia, and L. erythrogastra are adopted from Rastegar-Pouyani and Nilson (2002).
Molecular analyses Fourteen specimens belonging to the genus Luadakia (for details refer to Table 1) were used in the molecular analyses. DNA was extracted using non-organic DNA Extraction Procedure (Proteinase K and Salting out). MT-ND4 (NADH dehydrogenase subunit 4; from 1 to 642, ND4; from 643 to 801, tRNA His [complete], tRNA Ser [complete] and tRNA Leu [Partial]) was amplified using standard PCR procedures with the following primers: ND4F, 5'-CACCTATGA CTACCAAAAGCTCATGTAGAAGC-3' (Thaung et al., 2009) and LeuR, 5'-CATTACTTTTACTTGGAT TTGCACCA-3' (Arevalo et al., 1994). PCR reactions performed in 20 μl with the following conditions: Initial denaturation stage of 95°C (05:00) followed by 36 cycles with denaturation at 95°C (00:40), annealing at 50°C (00:40) and extension at 72°C (01:40) then single extension cycle at 72°C (05:00). The PCR products were sequenced with an automatic DNA sequencer using manufacture’s protocols by BIONEER (Daejeon, Republic of Korea). In order to conduct a more accurate molecular analysis, four sequences belonging to Laudakia lehmanni (KF691670.1), L. microlepis (KF691699.1), L. stoliczkana (AF128519.1) and L. caucasia (AY053998.1) were retrieved from GenBank (http://www.ncbi.nlm.nih. gov) and included to our dataset. Nucleotide sequences were aligned using ClustalW, as implemented in Bioedit version 7.0.5.3 (Hall, 1999) with default parameters. Using the software package Mega 6 (Tamura et al., 2013) genetic distances between taxa were calculated. Bayesian analysis was carried out using Mr. Bayes 3.1.2 (Huelsenbeck and Ronquist, 2001). Bayesian analysis was performed with two runs and four chains for each run for 4 million generations and the current tree was saved every 100 generations. A 50% majority rule consensus tree was produced from the posterior distribution of the trees, and the posterior probabilities were calculated as the percentage of samples recovering any particular clade, with posterior probabilities P ≥ 95% indicating significant support.
3. Results
A data set of 677bp of ND4 following by tRNAs, from 677 to 844, including tRNA His (complete) + tRNA Ser (complete) + tRNA Leu (Partial) was generated. Genetic divergence (Kimura 2 parameters) among different specimens varied from 5.5 to 17.8% (Table 2). Phylogenetic relationships (BI inference) between Laudakia microlepis and Taftan specimens based on 844 nucleotides clearly indicate that Taftan specimens fall apart from L. microlepis, as illustrated in Figure 2.
Based on the results, presented here, the Laudakia microlepis populations are distributed on the southeastern parts of Iran and at least some of them, namely that of the Taftan Mountain, are genetically different from other populations of L. microlepis, sufficient to be considered as a subspecies of the nominal species. As such we call this new subspecies, Laudakia microlepis taftanica ssp. nov., against the nominal species L. m. microlepis.
3.1 Description Laudakia microlepis taftanica ssp. nov. (Figure 3 A, B and C)
Holotype and type locality An adult male (Figure 3A, B), ZUTC, REP 1438, collected on 25thAugust 2011, 25km north of Khash, Sistan and Baluchistan province, Southeastern Iran (28° 36.0' 25.1" N 61° 04' 47.3" E).
Paratypes Thirteen specimens (seven males and six females) collected, from the same locality as holotype (see Appendix).
Diagnosis Body size large, head and body moderately depressed; grayish-black dorsally; a cutaneous fold on each side of the neck; a dorsolateral cutaneous fold on each side of the body with smooth scales; enlarged vertebral scales, weakly keeled; body scales small; no patch of enlarged mucronate scales on fanks; tail divided into two distinct segments, each composed of two whorls of scales.
Description of Holotype Upper head scales heterogeneous, spiny scales around the tympanum relatively smaller than scales around orbit, spiny scales on each side of the neck; enlarged mid-dorsal scales in 13 rows, median dorsal scales equal in size with ventral scales; ventral scales smooth, scales on upper limbs larger than median dorsal scales and distinctly keeled; dorsal scales heterogeneous, scales on fanks smaller thanmid-dorsal scales; gular scales smooth with patches of yellowish scales; 198 scales around middle of the body, 110 scales in single row from gular fold to anterior edge of vent; 27 scales around fifth caudal whorl just behind vent; with mid-ventral and pre-anal callous scales.
Coloration and color pattern Dorsal and ventral surfaces of the body proper grayish-black; posterior part of the head, around tympanum and dorsal part of the neck yellowish (Figure 3A, B); a patch of yellowish scales on the anterior part of the head; proximal part of the tail with black-cream rings, distal part of tail black.
Measurements (in millimeters) Total length 276.6; SVL 121.5; TL 155.1; HL 31.8; HW 23.2; HH 15.8; SN 6.4; NN 8.4; FLL 69.0; HLL 102.3; GFV 81.4; MTrW 41.4; MTaW 18.7.
Variations of type series All paratypes closely resemble the holotype in both morphometric and meristic traits. However, there are some minute differences between male and female paratypes. The folds on sides of neck are more developed in males than those in females. In some females mid-ventral and pre-anal callous scales are absent. In males, scales of upper side of limbs are more strongly keeled than those in females. In females middorsal scales are larger than ventral scales. Furthermore, in some females, dorsal surface contains scattered palegreen or brownish scales. In some males, except for black rostrum, other parts of the head are yellow in coloration. Male paratypes All male paratype specimens (n = 7) were similar to the holotype in both morphometric and meristic characteristics, with slight variations in some specimens. Range and mean (in parenthesis) of different characteristics are as follows: The SVL 101.6–129.6 (119.7), TL 127.8–172.1 (162.8), HL 26.8–33.0 (29.9), HW 20.1–25.4 (23.7), HH 10.8–16.1 (14.5), SN 4.7–6.4 (5.3), NN 6.5–7.8 (7.3), FLL 53.7–79.2 (67.4), HLL 76.6–106.5 (100.5), GFV 10.9–108.7 (83.2), MTrW 30.6–44.0 (39.1), MTaW 13.9–19.9 (18.2), SQ 184–197 (191.8), LVS 11–13 (11.6), AGFS 120–136 (127.3), FCWS 26–31 (28.5). Mid-dorsal scales enlarged, almost homogeneous, weakly keeled; all dorsolateral, gular and ventral scales similar to the holotype; coloration almost identical to that of the holotype.
Female Paratypes Females (n = 6) rather different from the holotype. Range and mean (in parenthesis) of differentcharacteristics are as follows: The SVL 99.7–119.8 (114.2), TL 121.9–162.8 (146.6), HL 22.7–29.3 (26.4), HW 19.8–23.7 (21.6), HH 11.4–14.1 (13.1), SN 3.6–5.7 (4.5), NN 6.0–7.2 (6.5), FLL 52.4–66.1 (59.4), HLL 75.6–99.9 (84.1), GFV 80.7–108.2 (88.7), MTrW 30.0–47.8 (40.3), MTaW 12.1–16.7 (14.8), SQ 182–200 (192), LVS 10–13 (11.3), AGFS 118–130 (123.5), FCWS 23–33 (27.2). Mid-dorsal scales almost homogeneous, weakly keeled; dorsolateral scales smaller than median dorsal scales; mid-ventral and pre-anal callous scales present in only one specimen; females coloration different from that of the male holotype in the presence of scattered palegreen scales on posterior dorsal and hind limbs as well as patches of yellow scales on shoulders.
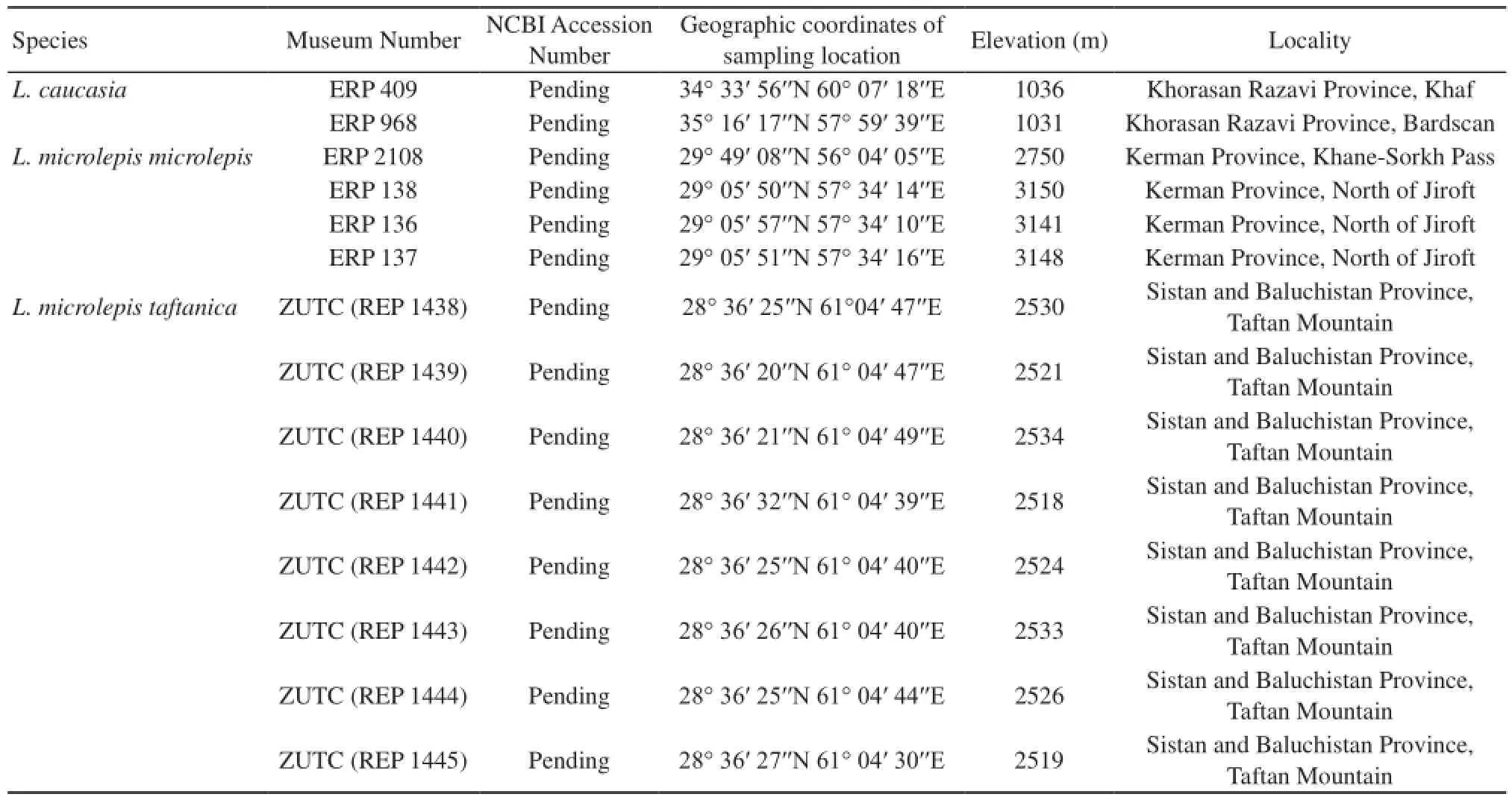
Table 1 List of the specimens studied in the present communication and their localities. ERP: personal collection of Dr. Eskandar Rastegar Pouyani, NCBI: National Center for Biotechnology Information ZUTC: Zoology Museum of University of Tehran.

Table 2 Genetic distances (Kimura 2 parameters) between Laudakia microlepis, L. microlepis taftanica ssp. nov., and related taxa.
4. Discussion
In comparison between different traits in specimens of Laudakia microlepis collected from Taftan Mountain and those collected from the type locality, we found some variations that, in combination with genetic variations, could justify the erection of a subspecies. Some of those traits have been used to distinguish different species within the genus. For example, In L. caucasia, males have 115–188 and females 119–174 scales around middle of body, respectively. These scales numbers are 80–114 in L. erythrogastra, 177–235 in males and 190–259 in female of L. microlepis microlepis (Anderson, 1999). In L. microlepis taftanica ssp. nov., males have 184–198 and females have 182–200 scales around middle of body (Table 3).

In L. caucasia mid-dorsal scales are fattened, keeled and in 7–11 longitudinal rows. In L. erythrogastra those scales often are large, keeled and mucronate, in 9–12 longitudinal rows (Rastegar-Pauyani and Nilson, 2002). In L. microlepis microlepis mid-dorsal scales are either fat or weakly carinated (Baig et al., 2012). In L. microlepis taftanica ssp. nov., mid-dorsal scales are large, weakly keeled, and arranged in 10–13 oblique longitudinal rows. Also, while L. caucasia has enlarged scales on fanks and smooth gular scales (Anderson, 1999), L. erythrogastra has flanks with several enlarged mucronate scales and strongly keeled gular scales (Baig et al., 2012), and L. microlepis microlepis has fanks with enlarged mucronate scales and small gular scales. L. microlepis taftanica ssp. nov., on the other hand, has fanks without enlarged scales and smooth gular scales. Sizes of different Laudakia species occurring in Iran are different. The snout-vent length of the largest L. caucasia male and female are 153 mm and 152 mm, respectively, and measures 133 mm and 149 mm for the largest male and female of L. microlepis microlepis (Anderson, 1999) and 148.5 mm and 150.5 mm for L. erythrogastra male and female, respectively (Aghili et al., 2010). Specimens of L. microlepis taftanica ssp. nov., were generally smaller than specimens of other species and unlike most other species, in L. microlepis taftanica ssp. nov., males were relatively larger than females (129.6 mm in male, 119.8 mm in female) (Table 3).
According to Baig et al. (2012) L. caucasia has 22–32 scales in the first complete whorl around the tail, whereas L. erythrogastra has 24–29 scales. In L. microlepis taftanica ssp. nov., those numbers were different for both males and females (27–31 in males and 23–33 in females).
Considering the color patterns, L. caucasia is light olive to dark gray, L. erythrogastra is olive-brown with many irregular black-edged light marks on vertebral region, and in L. microlepis microlepis shoulders and thorax are black, while the head, abdomen and anterior part of tail are light cream to yellow (Anderson, 1999). Even though L. microlepis microlepis color pattern is highly variable in different population (Baig et al., 2012), but the color pattern of L. microlepis taftanica ssp. nov., is distinct from both the nominal species and other close relative species.
In Iran, L. caucasia habitats range from sea level to elevation of about 4 000 m, on the northern parts of the Iranian plateau, while L. erythrogastra occupies northeastern parts of Iran to the vicinity of Mashhad, and L. m. microlepis habitats are the mountainous areas of the southern, central and eastern parts of the country (Anderson, 1999). L. microlepis taftanica ssp. nov. seems to be restricted to the Taftan Mountain at elevation of about 2 500 m in Sistan and Baluchistan province, southeastern part of Iran. It seems that the populations of the new subspecies were once connected to the populations of the nominal species on the mostsoutheastern edge of their distribution range, when the weather was more suitable and there were connecting populations in between during the last ice age. During the past 10 000 years, however, due to extreme changes in the weather and precipitation patterns (Lomolino et al., 2010) of the area, population of L. microlepis taftanica ssp. nov. is separated from populations of L. m. microlepis and driven to isolation on higher elevations of Taftan Mountain, where they found refuga, and became distinct in both molecular and morphological traits.
Habitat Typical habitat of this subspecies (Figure 3D) is mountainous terrain, with rocky out crops having crevices. The vegetation coverage consists of sparse Achillia, Astragalus, Alhagi, Cynedon, Phragmites, Lilium, Tamarix, Ziziphora, Rumex, Artemisia, Rheum, Zygophyllum, Anemone, Convolvulus, Ephedra and trees such as Tamarix, Armenica, Amygdalus, Acer, and Morus. Distribution Laudakia microlepis taftanica ssp. nov., is restricted to the Taftan Mountain at elevations of about 2 500 m, in Sistan and Baluchistan province, southeastern Iran (28° 36' 25.1" N; 61° 04' 47.3" E).
Etymology The new subspecies name is derived from its type locality, Taftan Mountain.
Comparison between different characteristics of lizard specimens sampled from Taftan Mountain, in Iran, with those of close relatives showed that these specimens belong to a new subspecies of Laudakia microlepis (Squamata: Agamidae). Reminding the place where this subspecies is found it is called Laudakia microlepis taftanica ssp. nov.
Acknowledgements We thank Mr A. Roohi Aminjan for his help with the statistical analyses. We would like to extend our gratitude to Mr H. Parsa and Mr H. Salehi for their assistance in specimen collections.
Aghili H., Rastegar-Pouyani N., Rajabizadeh M., Kami H. G., Kiabi B. H. 2010. Sexual dimorphism in Laudakia erythrogastra (Sauria: Agamidae) from Khorasan Razavi Province, Northeast Iran. Russ J Herpetol, 17: 51–58
Anderson S. C. 1999. The lizards of Iran. Saint Louis, Missouri: Society for the Study of Amphibians and Reptiles, I–VII, 442pp
Arevalo E., Davis S., Sites J. 1994. Mitochondrial DNA sequence divergence and phylogenetic relationships among eight chromosome races of Sceloporus grammicus complex (Phrynosomatidae) in central Mexico. Syst Biol, 43: 387–418
Baig K. J., Bohme W., Ananjeva N. B., Wagner P. 2012. A morphology-based taxonomic revision of Laudakia Gray, 1845 (Squamata: Agamidae). Verte Zoo, 62(2): 37–60
Blanford W. T. 1874. Description of New Lizards from Persia and Baluchistan. Ann Mag Nat Hist, 13: 453–455
Blanford W. T. 1876. Eastern Persia, an Account of the Journeys of the Persian Boundary Commission, 1870-71-72, Vol. II, The Zoology and Geology, 516 pp
Frost D. R., Etheridge R. 1989. A Phylogenetic Analysis and Taxonomy of Iguanian Lizards (Reptilia: Squamata). Miscellaneous Publications, University of Kansas Museum of Natural History, 81: 1–65
Hall T. A. 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser, 41: 95–98
Lomolino M. V., Riddle B. R, Whittaker R. J., Brown J. H. 2010. Biogeography. Sunderland, Massachusetts: Sinauer Associates Inc. Publishers, 878 pp
Macey J. R., Larson A., Ananjeva N. B., Papenfuss T. J. 1997. Evolutionary shifts in three major structural features of the mitochondrial genome among iguanian Lizards. J Mol Evol, 44: 660–674
Pyron R., Burbrink F., Wiens J. 2013. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol Biol, 13: 93
Rastegar-Pouyani N., Nilson G. 2002. Taxonomy and biogeography of the Iranian species of Laudakia. Zool Mid East, 26: 93–122
Schwenk K. 1994. Systematics and subjectivity: The phylogeny and classification of iguanian lizards revisited. Herpet Rev, 25:53–57
Šmid J., Moravec J., Kodym P., Kratochvil L., Hosseinian Yousefkhani S., Rastegar-Pouyani E., Frynta D. 2014. Annotated checklist and distribution of the lizards of Iran. Zootaxa, 3855: 1–97
Tamura K., Stecher G., Peterson D., Filipski A., Kumar S. 2013. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol Biol Evol, 30: 2725–2729
Thaung H., Willoughby T., Pradya S., Duong S., Setha T., Sein M., Chang M., Catherine W. 2009. Mitochondrial pseudogenes in the nuclear genome of Aedes aegypti mosquitoes: implications for past and future population genetic studies. BMC Genetics, 10: 11
Wettstein O. V. 1953. Herpetologia aegaea. Sber. Osterr. Akad. Wiss. 162: 651–683
Dr. Hassan RAHIMIAN, from College of Science, University of Tehran, Tehran, Iran, with his research focusing on lizards of Iran.
E-mail: h.rahimian@ut.ac.ir
26 November 2014 Accepted: 29 May 2015
杂志排行

Asian Herpetological Research的其它文章
- A New Species of Japalura (Squamata: Sauria: Agamidae) from Upper Lancang (Mekong) Valley of Eastern Tibet, China
- Seasonal Dynamics of Male and Female Reproductive Systems in the Siberian Salamander, Salamandrella keyserlingii (Caudata, Hynobiidae)
- Oviposition Site Selection in the Malayan Giant Frog (Limnonectes blythii) in Singapore: Conservation Implications
- Comparative Studies on Sperm Ultrastructure of Three Gecko Species, Gekko japonicus, Gekko chinensis and Hemidactylus bowrigii (Reptilia, Squamata, Gekkonidae)
- Potential Distribution Modeling and Morphology of Pelias barani (Böhme and Joger, 1983) in Turkey
- Characterization of the Genetic Diversity of Trachemys dorbigni and Phrynops hilarii