缺素对土沉香幼苗根系生长和叶绿素荧光参数的影响
2015-10-21贾晓红周再知马华明梁坤南黄桂华余雪标
贾晓红 周再知 马华明 梁坤南 黄桂华 余雪标


摘 要 采用营养液培养法,以土沉香幼苗为试验材料,研究在缺乏9种营养元素(即-N、-P、-K、-Ca、-Mg、-Fe、-Mn、-B、-Zn)环境条件下,苗木的根系生长、根系活力和叶绿素荧光参数的变化和响应机制。结果表明:(1)与全素对照相比,不同缺素处理下的土沉香幼苗根系生物量值和根系活力显著下降,其中,-Ca和-K处理影响最显著,其次为-Fe处理;(2)不同缺素处理稳态荧光产量(Ft),光适应时最大荧光(Fm′),叶绿体光系统II(PSⅡ)在部分反应中心关闭下实际光化学量子产量(Yield),非循环电子传输速率(ETR),光下最小荧光(Fo′)以及潜在最大光化学量子产量(Fv/Fm)值与CK处理差异显著,其中,-P处理Ft值最小,Fe和P元素对Fm′抑制最明显,-Fe处理对Yield值影响最大,-Mg处理的ETR值最小,-Fe处理抑制Fo′值最明显,-P处理抑制Fv/Fm值最明显。
关键词 土沉香;缺素;根系生物量;根系活力;叶绿素荧光参数
中图分类号 S567.19 文献标识码 A
土沉香[Aquilaria sinensis(Lour.)Gilg](别名白木香、女儿香、牙香树、莞香、六麻树),为瑞香科(Thymelaeaceae)沉香属(Aquilaria Spp.),热带、亚热带常绿乔木,是传统名贵药材和天然香料,主要分布于广东、海南、云南、广西、福建、台湾等地区,被列为国家二级重点保护植物名录[1-4]。近年来,在培育土沉香人工林过程中,时常出现苗期或者种植时期,植株叶片发黄、矮小、萎蔫等症状,为了解决这些问题,学者开始致力于土沉香立地、营养施肥等方面的研究。王冉等[5]设置6个施肥量处理,进行珍贵树种马来沉香与土沉香苗期 N 素指数施肥试验研究,探究不同 N 素供应水平对其苗期生长、光合生理特性的影响。杨晓清等[6]以土沉香3个种源的幼苗为材料,模拟水分胁迫,研究了氮素营养与水分胁迫对土沉香不同种源幼苗相关生理生化指标的耦合影响。但是利用叶绿素荧光技术进行土沉香营养诊断研究尚未见报道。因此,本试验试图利用温室营养液培养法,观测土沉香幼苗在不同缺素(即缺乏氮、磷、钾、钙、镁、铁、锰、硼、锌)条件下根系生长、根系活力和叶绿素荧光参数的变化,为土沉香在缺素环境下的生理适应机制以及土沉香人工林的健康发展提供科学依据。
1 材料与方法
1.1 材料
采用长势良好,大小均一(平均高度为9.2 cm,地径为2.1 mm)的土沉香幼苗,用去离子水洗净根部,于圆形黑色容器(上下口径19 cm×高20 cm,容积5 L)内,用海绵固定幼苗,置于中国林业科学研究院热带林业研究所温室内进行悬浮培养。
1.2 方法
1.2.1 试验设计及处理 试验采用随机区组设计,10个处理,重复3 次,每个处理3株苗。10个处理分别是:①对照(CK,全素营养液);②缺氮(-N);③缺磷(-P);④缺钾(-K);⑤缺钙(-Ca);⑥缺镁(-Mg);⑦缺铁(-Fe);⑧缺锰(-Mn);⑨缺硼(-B);⑩缺锌(Zn)。
移苗后用1/2浓度量的全营养液(即霍格兰大量元素及阿农微量元素1938 a配方[7])培养1周。从第2周起,采用全浓度和10个处理配置营养液(即不同缺素及对照)培养。容器内营养液以能淹没土沉香幼苗根系为宜,培养期间每天充氧气1次,持续45 min,每隔1周换1次营养液,并及时清理掉水箱内青苔。定期用NaOH溶液或HCl调整箱内 pH值至6.5,减少光照温度等外因的干扰并注意保持温室内的清洁卫生,通气良好。试验期间,温室不使用人工光源,白天平均温度应保持在23~38 ℃,平均湿度 54%~78%。试验从2012年9月开始到2013年6月结束,持续9个月,期间观测不同缺素处理幼苗的表型特征,收获时候测定幼苗的根系生物量、根系活力以及叶绿素荧光参数。
1.2.2 取样调查与指标测定 试验结束时,剪下植株根系测定其鲜重,并在70 ℃的烘箱中烘至恒重,称取根干重。用氯化三苯基四氮唑(TTC)法测定根活力[8];并于晴朗无风天气,采用便携式调制叶绿素荧光仪PAM2500测定每株苗木3片健康叶片的稳态荧光产量Ft(Chlorophyll fluorescence)、光适应时最大荧光Fm′( Maximum fluorescence)、叶绿体光系统II(PSII)在部分反应中心关闭下实际光化学量子产量Yield(Photochemical quantum yield of PSII in the light)以及非循环电子传输速率ETR(Electron transport rate);暗适应下的初始荧光Fo和最大荧光Fm值于晚上20 : 30测定。按下列公式计算光下最小荧光Fo′(Minimum fluorescence light)和潜在最大光化学量子产量Fv/Fm(Potential maximum PSII quantum yield)[9]。
Fo′=Fo/(Fv/Fm+Fo/Fm′);Fv/Fm=(Fm-Fo)/Fm。
1.3 数据处理
用Excel2007对数据进行整理统计,SPSS18.0进行方差分析和Duncan多重比较。
2 结果与分析
2.1 不同缺素处理对土沉香幼苗根系生长和活力的影响
植物根系是活跃的吸收器官和合成器官,根的生长情况和活力水平直接影响地上部分的营养状况及产量水平[10]。由表1可知,CK处理的根系生物量均显著高于各缺素处理(p<0.05),其值为根系生物量值最小-K和-Ca处理的3.53倍。9个缺素处理中,地下部分生物量最大的是-Mn和-B处理,比-K和-Ca处理增大了1.33倍,各处理之间差异不显著。
缺素处理环境下土沉香幼苗的根系活力有显著性差异(p<0.05)。各处理的根系活力从高到低依次為CK>-Zn>-Mn>-B>-N>-Fe>-P>-Mg>-K>-Ca,其中,CK处理的根系活力显著高于其他缺素处理;缺素处理中对根系活力影响最显著的是-Ca处理和-K处理,均比CK处理分别降低了80.0%;对根系活力影响最小的为-Zn处理,其次为-P、-Mg和-Fe处理,分别比CK处理降低了10.0%,60.0%和70.0%。
2.2 不同缺素处理对土沉香幼苗叶绿素荧光参数影响
植株的生长发育离不开光合作用,光合作用是生物界所有物质代谢和能量代谢的物质基础,通过测定植株光合作用对不同外界环境的生理响应,可以判断植物的生长和健康状况与其生境之间的关系,叶绿素荧光参数能够反映光合作用的“内在性”特点[9-11]。
2.2.1 缺素处理对稳态荧光产量(Ft)的影响 方差分析表明,不同缺素处理对土沉香幼苗的叶绿素荧光参数有显著影响(p<0.05)。除了-Ca和-B处理之外的其他缺素处理均使土沉香幼苗叶片的Ft值显著低于CK处理,其中Ft值最小的是-P处理,比CK处理降低了40.0%;其次为-K和-Fe处理,分别比CK处理降低了32.0%和36.0%。9个缺素处理的Ft值大小依次为-B>-Ca>-Zn>-Mn>-N>
-Mg>-K>-Fe>-P,其中-B处理与除-Ca处理外的其他缺素处理差异性显著,比-P处理增大了0.40倍(图1)。
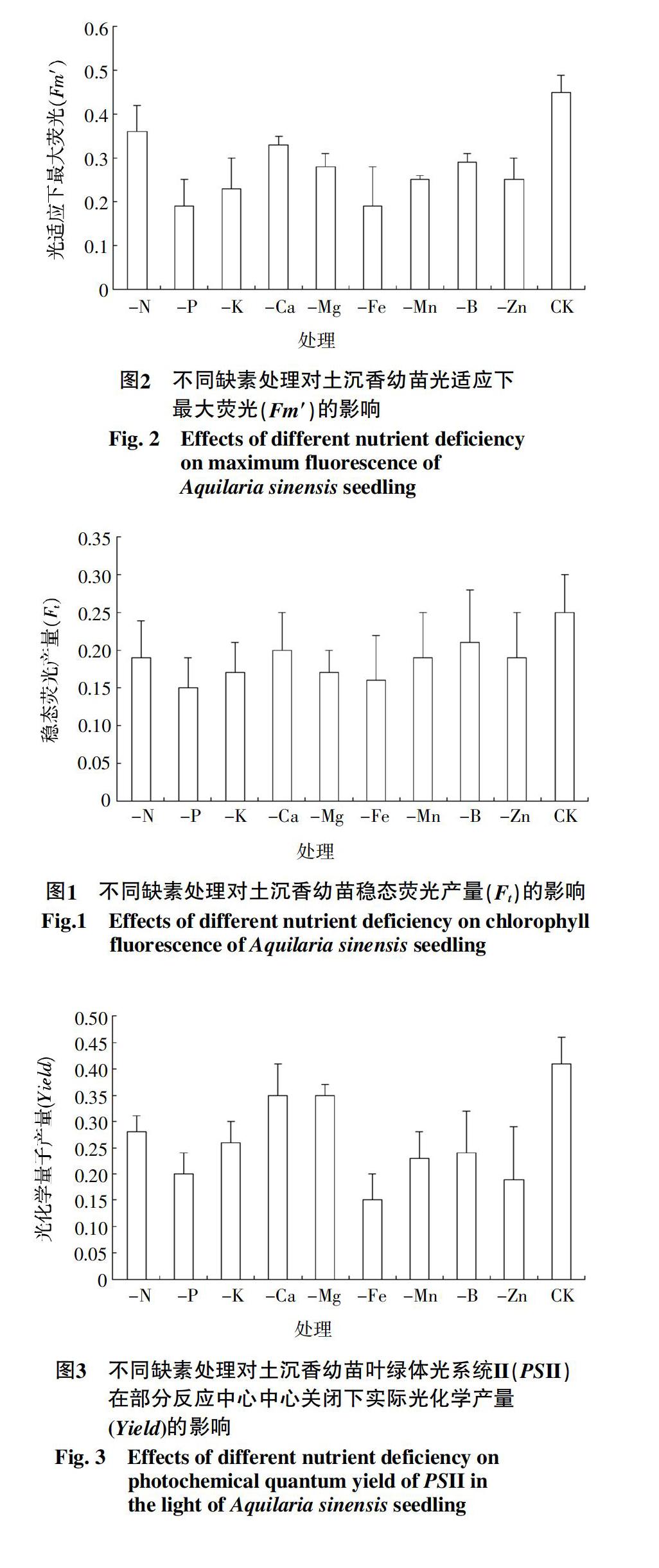
2.2.2 缺素处理对光适应下最大荧光(Fm′)的影响
各缺素处理中除-N处理之外均使土沉香幼苗的Fm′值显著低于CK处理,其中土沉香幼苗Fm′ 值受到最明显抑制的是-Fe处理和-P处理,值均为0.19,比CK处理降低了57.68%;其次为-K处理,比CK处理降低了20.0%。9个缺素处理的Fm′值大小顺序为-N>-Ca>-B>-Mg>-Mn>-Zn>-K>
-P>-Fe,其中-Fe处理与其他缺素处理的差异性显著,约占最大值-N处理的1/2(图2)。
2.2.3 缺素处理对叶绿体光系统II(PSII)在部分反应中心关闭下的实际光化学量子产量(Yield)的影响
由图3可知,-N,-Ca以及-Mg处理的土沉香幼苗的叶片Yield值与CK处理差异性不显著,-P,-K,-Fe,-Mn,-B和-Zn处理的Yield值显著低于CK处理,其中对其值抑制最明显的是-Fe处理,值仅为0.15,比CK处理降低了63.4%;其次为-Zn处理和-P处理,分别比CK处理降低了53.7%和51.2%。各个缺素处理的Yield值大小顺序是-N >-Mg>-Ca>-K>-B>-Mn>-P>-Zn>-Fe,其中-Fe处理与-N,-Mg,-Ca,-K,-B以及-Mn处理的差异性显著,约占最大值-N处理的2/5。
2.2.4 缺素处理对非循环电子传递速率(ETR)的影响
各个缺素处理的土沉香幼苗的叶片ETR值显著低于CK处理,其中-Mg处理的ETR值最小,仅为6.67,比CK处理降低了69.91%,其次为-Ca处理和-K处理,分别比CK处理降低了68.2%和67.7%。各个缺素处理的ETR值大小顺序是-N>
-Mn>-B>-Zn>-Fe>-P>-K>-Ca>-Mg,其中-Mg处理与-N,-Mn,-B,-Zn和-Fe处理的差异性显著,约占最大值-N处理的2/5(图4)。
2.2.5 缺素处理对光下最小荧光(Fo′)的影响 图5表明,9个缺素处理中,除-N和-Ca处理外的其他缺素处理的土沉香幼苗的叶片Fo′值均显著低于CK处理,其中抑制Fo′值最明显的是-Fe处理,比CK处理降低了48.3%;其次为-P处理和-K处理,分别比CK处理降低了44.8%和40.0%。各个缺素处理的Fo′值大小顺序是-N>-Ca>-B>-Mg>-Mn>
-Zn>-K>-P>-Fe,其中-Fe处理与-N处理的差异性显著,约占最大值-N处理的1/2。
2.2.6 缺素处理对潜在最大光化学量子产量(Fv/Fm)的影响 如图6所示,9个缺素处理中,除-Fe和-Zn处理外的其他缺素处理的土沉香幼苗叶片Fv/Fm值均显著低于CK处理,其中抑制Fv/Fm值最明显的是-P处理和-Ca处理,值均为0.09,比CK处理降低了60.9%;其次为-N处理,比CK处理降低了52.2%。各个缺素处理的Fv/Fm值大小顺序为-Zn>-Fe>-B>-K>-Mn>-Mg>-N>-Ca>-P,其中-Fe和-Zn处理与其他7个处理的差异性显著,且-Zn处理比-P处理增大了1.11倍。
3 讨论与结论
植物生长发育需要各种矿质元素以维持正常的生理活动[12],缺乏不同的矿质元素均会影响植株的根系生长和发育,进而影响叶片的光化学反应。根系生长对矿质元素反应敏感,营养元素的亏丰一定程度上影响根系生物量的积累。本试验中,9个缺素处理的土沉香根系总生物量都低于全素处理(CK),与King等[13]对火炬松(Pinus taeda Linn.)和北美黄松(Pinus ponderosa)苗木试验的结论相似。影响土沉香苗木根系生物量生长最显著的是-K处理,这是由于K是植物体内许多酶的活化剂,对韧皮部传导,渗透调节等具有重要作用。根系活力是反映植株根系代谢能力强弱的指标之一,其大小影响植株的生长和抗逆性,关系到植株对水分和矿质营养的吸收利用[14]。麦荣章等[15]的研究结果显示,缺磷、氮和钙处理的春石斛(Dendrobium spp.)普遍表现出根系活力的降低,试验处理l个月后缺磷处理最低;2个月后缺氮处理最低,其次为缺钙处理。本试验与其研究结果略有差异,9个缺素处理中,对土沉香根系活力影响最显著的是-K和-Ca处理,可能因为Ca是植物细胞壁胞间层中果胶酸钙的组成成分,因此,缺钙时,细胞分裂不能进行或不能完成,而形成多钙细胞。也有研究报道指出,钙对于植株细胞壁和原生质膜稳定性具有重要作用[16],因此,-Ca处理土沉香幼苗叶片呈淡绿色,继而根尖出现坏死现象。同时,K在植物根系的生长发育过程中扮演着重要的角色,推测原因可能是缺K改变了土沉香对K的吸收转运,引起表皮细胞内K+浓度的变化,进而影响根系活力高低。
多个研究结果表明,矿质元素的缺乏会导致植株叶绿素荧光参数(Fm,Fv,ETR等)值的降低[17-19]。本试验结果显示,P、Mg、Ca和Fe是影响土沉香幼苗叶绿素荧光参数的主要元素。P是许多参与光合和呼吸作用的辅酶如NAD+、NADP+等的成分,Mg是叶绿素的组成成分之一,在光合和呼吸过程中,可以活化各种磷酸变位酶和磷酸激酶,Fe也是许多和光合有關的酶的辅基,土沉香幼苗在缺乏这几种元素的生长环境下,Ft、Fm′、Yield、ETR、Fo′、Fv/Fm值显著降低,说明缺素已经造成了光抑制,导致其PSII潜在活性中心受到损伤,其反应中心的光化学效率受到明显影响,光合结构遭到破坏,不利于激发能由捕光色素蛋白复合体(LHC)向PSII进行传递。这与李绍长等[20]、杨勇等[21]研究结果一致。
本试验研究的时间比较长,可能原因是土沉香叶革质,耐贫瘠,对缺素培养的反应不灵敏造成的。而且本试验整个过程都是在温室内进行,试验环境与野外相比存在一定的差异,因此,今后还需要进一步通过其他诊断方法(砂培法等)开展野外试验,为土沉香的快速营养诊断提供科学参考。
参考文献
[1] 江苏新医学院. 中药大辞典[M]. 上海: 人民出版社, 1977.
[2] 裘树平, 刘仲荃. 中国保护植物[M]. 上海: 上海科技教育出版社, 1994.
[3] 李林海, 寿海洋, 马清温. 土沉香(瑞香科) 的地理分布研究[J]. 安徽农业科学, 2012, 40(17): 9 254-9 256.
[4] 国家林业部, 国家农业部令. 国家重点保护野生植物名录(第一批)(第4号)[EB]. http: //www.gov.cn/gongbao/content/2000/content_60072.htm, 1999-09-09.
[5] 王 冉, 何 茜, 丁晓纲, 等. N素指数施肥对沉香苗期光合生理特性的影响[J]. 北京林业大学学报, 2011, 33(6): 58-33.
[6] 杨晓清, 周再知, 梁坤南, 等. 氮素对模拟胁迫下土沉香幼苗抗旱生理的影响[J]. 热带作物学报, 2013, 34(6): 1 121-1 127.
[7] 陈 琳. 西南桦苗期营养诊断与氮素施肥[D]. 北京: 中国林业科学研究院, 2010.
[8] 李合生. 植物生理生化实验原理和技术[M].北京: 高等教育出版社, 2000.
[9] 朱成刚, 李卫红, 马晓东, 等. 塔里木河下游干旱胁迫下的胡杨叶绿素荧光特性研究[J].中国沙漠, 2011, 31(4): 917-936.
[10] 李 晓, 冯 伟, 曾晓春. 叶绿素荧光分析技术及应用进展[J]. 西北植物学报, 2006, 26(10): 2 186-2 196.
[11] Oxborought K, Baker N R. Resolving chlorophylla fluorescence images of photosynthetic into photo-chemical and non-photochemical components-calculation of qP and Fv′/Fm′ without measuring Fo′[J]. Photo- synthesis Research, 1997, 54: 135-142.
[12] 潘瑞炽. 植物生理学[M]. 北京: 高等教育出版社, 2003.
[13] King J S, Thomas R B, Strain B R. Morphology and tissue quality of seeding root systems of Pinus teada and Pinus ponderosas affected by varying CO2, temperature and nitrogen[J]. Plant and Soil, 1997, 195: 107-119.
[14] 顿 颖. 柑橘水培体系优化及N、 Fe胁迫下根系形态研究[D]. 武汉: 华中农业大学, 2011.
[15] 麦荣章, 李 丽, 甘林叶. 不同营养液和缺素处理对春石斛几个生长指标的影响[C]. 中国观赏园艺研究进展, 2009: 347-354.
[16] Ericsson T. Growth and shoot: root ratio of seedlings in relation to nutrient availability[J]. Plant and Soil, 1995, 168-169: 205-214.
[17] 郭培國, 陈建军, 郑燕玲. 氮素形态对烤烟光合特性影响的研究[J]. 植物学通报, 1999, 16(3): 262-267.
[18] 董彩霞, 赵世杰, 田纪春, 等. 不同浓度的硝酸盐对高蛋白小麦幼苗叶片叶绿素荧参数的影响[J]. 作物学报, 2002, 28(1): 59-64.
[19] 郭延平, 陈屏昭, 张良诚等. 缺磷胁迫加重柑橘叶片光合作用的光抑制及叶黄素循环的作用[J]. 植物营养与肥料学报, 2003, 9(3): 359-363.
[20] 李绍长, 胡昌浩, 龚 江, 等. 低磷胁迫对磷不同利用效率玉米叶绿素荧光参数的影响[J]. 作物学报, 2004, 30(4): 365-370.
[21] 杨 勇, 蒋德安, 孙骏威, 等. 不同供镁水平对水稻叶片叶绿素荧光特性和能量耗散的影响[J]. 植物营养和肥料学报, 2005, 11(1): 79-86.
